9 .2. Происхождение и ранние стадии развития жизни
9.2. Происхождение и ранние стадии развития жизни
Современная теория происхождения жизни — это теория появления структур, которые, развиваясь и усложняясь, приобретали определенные функциональные свойства. По-видимому, эти свойства могли возникнуть уже на ранних этапах эволюции, так как некоторые полимеры обладали рядом характеристик, присущих феномену жизни. Однако, несмотря на огромные успехи теории происхождения жизни на Земле, существует множество проблем, которые ждут своего решения.
После знаменитой работы А. И. Опарина, увидевшей свет в 1924 г., а затем работ Дж. Холдейна (Haldane, 1969), Дж. Бернала (Bernal, 1969) и многих других проблема происхождения жизни стала предметом сначала теоретического, а затем и экспериментального научного анализа. В течение длительного времени эту проблему относили главным образом к проблемам химии и биохимии, которые рассматривались применительно к условиям определенного этапа геологического развития Земли. Затем к этим представлениям были присоединены некоторые другие взгляды, в том числе взгляды физиологов.
Интерпретация происхождения жизни на Земле в значительной степени зависит от понимания ее сущности. Мы не будем пытаться уточнять понятие жизни и отошлем читателя к обширной и противоречивой литературе. Заметим лишь, что в отличие от многих натуралистов, и в частности А. Сент-Дьердьи (Szent-Gyorgyi, 1947), которые считают, что в основе жизни лежат лишь немногие фундаментальные принципы организации и эволюции живых систем, мы придерживаемся другой точки зрения. Мы полагаем, что число таких принципов очень велико и многие из них еще предстоит открыть (Уголев, 1985, 1987а, 1989). Другими словами, жизнь не может получить объяснение в свете небольшого числа сколь угодно важных законов, так как фундаментальной особенностью жизни как процесса является многосущность, из чего вытекает и невозможность одного способа ее описания. Нам, однако, достаточно понимания того, что жизнь — это многосущностный процесс с чрезвычайно сложной организацией во времени и пространстве.
В отношении происхождения первых клеток были высказаны две основные гипотезы. Одна из них, впервые выдвинутая еще А. И. Опариным, основное значение придает образованию ферментов и формированию клеточных границ. При этом постулируется, что генетическая система возникла позднее и первооснову жизни составляли белки. Сходной точки зрения придерживается С. Фокс (Fox, 1980). Вторая гипотеза утверждает, что потенциальной формой жизни была нуклеиновая кислота, названная «голый ген», на основе которого стали возможны построение и эволюция белков (см., например, Lehninger, 1974; Florkin, 1975; Dubos, 1978).
Питание является одним из первичных процессов. Мы полагаем, что оно имеется у тех систем, которые носят название протоклеток и микросфер (Уголев, 1985, 1989). Согласно гипотезе А. И. Опарина, первые предклетки появились в тот момент, когда вокруг одной или нескольких макромолекул каталитически активных белков возникла граница, или мембрана. Термин «протоклетка» принят для обозначения промежуточного звена в процессе возникновения жизни. Это звено, по-видимому, находилось между эволюирующими химическими соединениями и биологическими формами, обладающими генетическим аппаратом и подвергающимися естественному отбору. Существенно, что протоклетки обладали уже некоторыми биологическими признаками.
Древнейшие органические микроструктуры имеют возраст около 4 млрд. лет. Вполне вероятно, что они и были протоклетками. Как отмечал К. Фолсом (Folsome, 1982), протоклетки образовались в первобытных водоемах из полимерного материала одновременно с органическими соединениями. А. И. Опарин в качестве модели протоклеток предложил коацерватные капли, состоящие из смеси коллоидных частиц. Этими частицами могли быть неспецифические макромолекулы, связывающие молекулы воды, и при определенных значениях pH среды, концентрации солей и температуры соединяющиеся друг с другом с образованием коацерватных структур. Последние по своим размерам напоминали клетки. Некоторые коацерватные капли могли поглощать какое-либо низкомолекулярное вещество (глюкозу или аминокислоты), а также примитивный катализатор. Тогда внутри капли могли аккумулироваться как субстрат, так и катализатор. Из этого следует, что такие капли могли обладать примитивным метаболизмом, состоящим из одной реакции.
В соответствии с гипотезой А. И. Опарина, капли, наделенные метаболизмом, должны были взаимодействовать с водным окружением и поглощать все новые соединения, включающиеся в структуру капель, что давало возможность их роста. Под действием ряда физических факторов капли могли распадаться на более мелкие. Некоторые из них могли сохранять в себе молекулы катализатора, что способствовало росту и образованию нового поколения капель. Идеи А. И. Опарина получили экспериментальное подтверждение.
С. Фокс и многие другие (см. обзоры: Fox, Dose, 1975; Fox, 1980; Эволюция, 1981) описали структуры, полученные в модельных экспериментах, которые были названы протеиноидами. Протеиноиды представляют собой белковоподобные полимерные молекулы, преимущественно со случайной последовательностью аминокислот, образующиеся при отщеплении одной молекулы воды в расчете на каждую пептидную связь. Протеиноиды в растворе имеют форму сфер диаметром около 10 мкм и напоминают клетки, ограниченные толстой мембраной. Протеиноиды с большой молекулярной массой, иногда превосходящей 10000, содержат до 18 различных аминокислотных остатков и обладают рядом свойств, характерных для белков. Более того, у некоторых протеиноидов обнаружена способность повышать скорость определенных химических реакций, а у других — хотя и слабая, но отчетливая гормональная активность. Эти факты свидетельствуют, что полипептидные цепи с определенной аминокислотной последовательностью могут возникать самопроизвольно при сравнительно простых условиях.
Протеиноиды обладают также функциями узнавания и дискриминации, которые проявляются в избирательном взаимодействии или отсутствии такового. Наконец, несомненный интерес представляют сообщения о связи различных ферментативных активностей с присутствием в протеиноидах молекул различных типов. Так, протеиноиды, содержащие гистидин, характеризуются эстеразной активностью, а протеиноиды, содержащие гем, — пероксидазной. Вместе с тем, как справедливо отмечает С. Фокс (Fox, 1980), между протеиноидными системами и настоящей жизнью лежит пропасть, так как они не могут осуществлять саморепродукцию. Эта трудность интерпретации не преодолена до сих пор.
С. Фокс и его сотрудники (обзоры: Fox, Dose, 1975; Fox, 1980) описали также самоорганизующиеся структуры — микросферы, напоминающие клетки. Эти структуры появлялись, в частности, при медленном охлаждении образовавшихся при повышенной температуре концентрированных растворов протеиноидов.
Авторы полагают, что аминокислоты концентрировались в испаряющихся водоемах под влиянием тепла и потоков лавы или полимеризовались при высыхании под действием солнечных лучей. После дождя протеиноиды, возникшие в результате самосборки, могли образовывать микросферы, в первом приближении напоминающие примитивные клетки. Последние представляли собой популяцию протоклеток, подвергавшихся отбору в соответствии с их каталитическими активностями, необходимыми для первичного обмена веществ. С. Фокс (Fox, 1980) в своем известном обзоре привел подробный перечень свойств протеиноидных макромолекул и микросфер.
Микросферы — довольно однородные сферические капли диаметром около 2 мкм. При определенных значениях pH внешняя граница микросфер могла приобретать структуру, напоминающую мембрану, хотя липиды в микросферах отсутствуют. Микросферы могли распадаться, делиться или почковаться. Почки могли отделяться от основной микросферы и давать начало новому поколению микросфер. При слиянии микросфер, содержащих различные каталитические активности, возникали микросферы, обладающие исходными активностями. Эти наблюдения важны для понимания механизмов возникновения систем с многими функциями и имеют значение для понимания развития «сложной» жизни из элементов. Таким образом, микросферы являются самоорганизующимися системами и представляют собой полезные модели первых примитивных структур, напоминающих клетки. Сам факт, что микросферы и коацерватные капли обладают зачатками метаболизма, позволяет предполагать, что метаболизм мог служить первым этапом формирования жизни.
Существование никросфер обычно характеризуется как «преджизнь», или «прозябание» (Уголев, 1985). Следует обратить внимание на некоторые важные обстоятельства. Во-первых, при определенных условиях преджизнь, развиваясь, превратилась в жизнь. Во-вторых, на протяжении длительного времени (возможно, многих сотен миллионов лет) феномены преджизни и жизни сосуществовали, что важно для понимания как их баланса, так и эволюции жизни в целом.
Многие авторы обратили внимание на то, что микросферам свойственны функции, характерные для живых систем. Поэтому весьма привлекательно характеризовать эти первичные функции как протофункции. Анализ процессов, реализуемых микросферами, показал, что они воспроизводят взаимодействия, которые С. Фокс обозначил как протосекс (протосексульные реакции). К протофункциям микросфер могут быть отнесены также протодвижение, протосвязь, проторепродукция (проторазмножение), компартментализация, защитные свойства и др., охарактеризованные С. Фоксом в 1980 г. Кроме того, микросферы растут (это характеризуется как проторост), стареют и погибают (табл. 9.1).
Таблица 9.1. Протоповеденческие свойства протеиноидов и протоклеток (по: Fox, 1980)
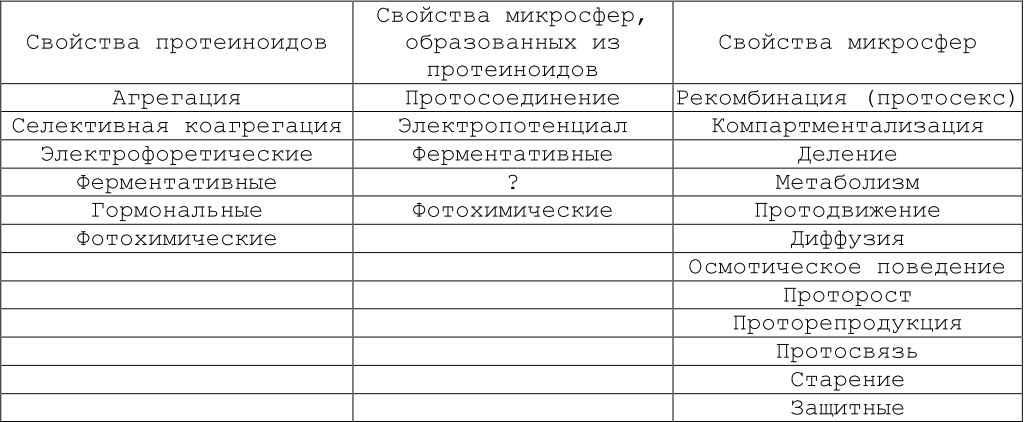
Примечание. Все свойства, обсуждаемые здесь, рассматриваются как протоповеденческие, эволюирующие до современного поведения. <?> — исследования не проводились
Мы ввели понятия прототрофии и протопепсии, имея в виду первичное питание и первичное пищеварение (Уголев, 1985). Мы также выделили два типа прототрофии. К первому из них может быть отнесена диффузия молекул (аминокислот, пептидов, сахаров) или частиц в протоклетки, ко второму — фузия, или слияние, двух протоклеток. Последний механизм во многом похож на первичный процесс размножения. Действительно, в обоих случаях имеет место слияние двух протоклеток с образованием одной большего размера и интеграцией часто неоднородных элементов. Мы допустили, что некоторые протоклетки, богатые гидролитическими ферментами, будут активно лизировать другие протоклетки, являясь своеобразными протохищниками. Этим мы не хотели показать, что такие протохищники могли служить предками современных редуцентов. Важно другое — уже на заре жизни могли формироваться основные свойства и механизмы биосферы с ее трофическими цепями и взаимодействиями, переносом веществ и энергии и т. д. При этом, как упомянуто выше, трофические цепи начинали строиться «с конца», т. е. с дессимиляторной, а не с синтетической части цикла (первая уже создавалась абиогенным путем). На самых ранних этапах возникновения жизни, по-видимому, появилась дифференциация на утилизируемые организмы и организмы, утилизирующие биологические материалы.
Наконец, согласно нашей гипотезе, весьма важно, что способность к аутолизу у протоклеток при некоторых условиях могла трансформироваться в гетеролиз. Этот процесс, вероятно, был по-разному выражен в различных протоклетках. В связи с этим появилась первичная дифференциация на так называемые клетки-протоаккумуляторы, накапливающие органические материалы, и клетки-проторедукторы, которые этот материал первично использовали. При всех обстоятельствах возникали первичные примитивные трофические цепи.
Наиболее ранний этап возникновения жизни на Земле в современных теориях, как и в теории А. И. Опарина, заключался в формировании основных типов мономеров, составляющих основу биологических структур, т. е. аминокислот, моносахаридов, оснований нуклеиновых кислот, порфиринов и т. д. (рис. 9.1). Вслед за этим должно происходить образование полимеров, характерных для живых систем (протеиноидов и некоторых других). Предбиологическая эволюция приводила к формированию примитивных клеток. С этого момента начинается биологическая эволюция, связанная с естественным отбором. Вероятно, образование первичных клеток явилось критическим моментом в эволюции жизни и тем рубежом, который разделял химическую эволюцию и биологическую. Именно клетка обладает рядом свойств, которые характеризуют истинную жизнь,(и, по-видимому, именно на этом этапе сформировались основные универсальные функциональные блоки, в сжатой форме охарактеризованные в гл. 1 (рис. 9.2).
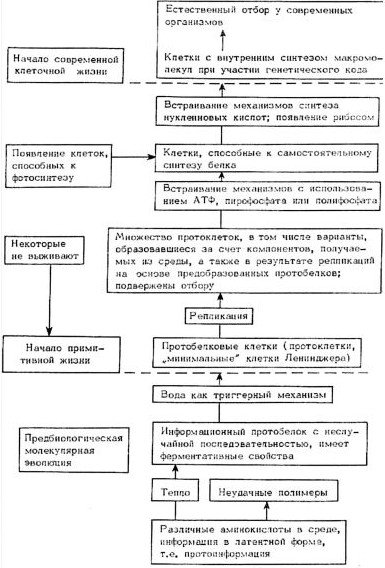
Рис. 9.1. Схема основных этапов молекулярной эволюции и происхождения жизни на Земле (по: Fox Dose, 1975).
Заслуживает внимания аналитико-исторический обзор Б. М. Кедрова и К. Б. Серебровской (1980), в котором хорошо документировано мнение большинства исследователей. Авторы отмечают, что современная теория происхождения жизни — это теория возникновения структур, приобретающих при усложнении и развитии определенные функциональные свойства. По-видимому, эти свойства могут проявляться уже на ранних этапах эволюции, так как некоторые полимеры, особенно протеиноиды, обладают рядом характеристик, присущих жизни. Протеиноиды, организованные в микросферы, обладают уже многими свойствами клеток, в том числе прототрофией и ее первичными механизмами (протопепсией). Вероятно, уже на этом этапе (а может быть, еще ранее) началась организация живого в протобиосферу, что существенно для понимания закономерностей дальнейшей эволюции.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Главные ступени развития жизни на земле
Главные ступени развития жизни на земле Развитие жизни на земле от её первого начала до нашего времени продолжается миллиарды лет. За это долгое время жизнь на земле прошла через ряд ступеней от более простого к более сложному и совершенному. Вот главные такие
Глава VI Этнология, происхождение и история человеческой половой жизни до брака
Глава VI Этнология, происхождение и история человеческой половой жизни до брака Самым важным, как мы видели, при изучении полового вопроса является умение оставаться в пределах объективности, избегая также сентиментального многословия, как эротизма. Но в истории половой
Глава 1. Происхождение жизни
Глава 1. Происхождение жизни Вопрос о происхождении жизни волнует всех, и очень жаль, что он пока еще далек от разрешения. Основная сложность тут в том, что путь от неорганических молекул к первой живой клетке был долгим и трудным. За один шаг такие превращения не
ГЛАВА 4 Происхождение жизни: абиогенез и панспермия. Гиперцикл. Геохимический подход к проблеме
ГЛАВА 4 Происхождение жизни: абиогенез и панспермия. Гиперцикл. Геохимический подход к проблеме Рассмотрев вопросы, связанные с эволюцией самой Земли, мы приступаем теперь к изучению эволюции жизни на ней. Сразу оговорюсь: я не собираюсь ни углубляться в дебри
Происхождение жизни. Наука и вера Предисловие
Происхождение жизни. Наука и вера Предисловие Влияние достижений науки и техники на жизнь человека огромно. Еще в XIX веке в большинстве семей можно было ожидать смерти одного или нескольких детей от различных болезней. Сегодня же в Соединенных Штатах и других развитых
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы Пер. А. НеизвестногоВ предыдущей главе мы обсудили возможные сценарии возникновения клеток и (будем надеяться) достигли
ПРОИСХОЖДЕНИЕ ЖИЗНИ
ПРОИСХОЖДЕНИЕ ЖИЗНИ Основные теории, предложенные на этот счет, можно свести к четырем гипотезам:1. Жизнь не имеет начала. Жизнь, материя и энергия сосуществуют в бесконечной и вечной Вселенной.2. Жизнь была создана в результате сверхъестественного события в особый
Глава 3. Происхождение жизни: химическая эволюция
Глава 3. Происхождение жизни: химическая эволюция Ничтожное ничто — начало всех начал. Теодор Рётке, "Вожделение" Теория химической эволюции — современная теория происхождения жизни — также опирается на идею самозарождения. Однако в основе ее лежит не внезапное (de novo)
Происхождение жизни
Происхождение жизни Как уже отмечалось, теория биохимической эволюции является единственной теорией в рамках научной методологии по вопросу происхождения жизни. Впервые она была предложена А. И. Опариным (1894–1980) в 1924 году. В дальнейшем автор неоднократно вносил в нее
Глава 1. Происхождение жизни
Глава 1. Происхождение жизни Планета бешено вращалась. Ночь и день сменяли друг друга с головокружительной быстротой: день длился всего пять-шесть часов. Тяжелая Луна угрожающе висела в небе (гораздо ближе, чем сегодня), отчего казалась крупнее. Звезды выглядывали редко,
1.5. Происхождение жизни
1.5. Происхождение жизни Эта проблема так же неразрешима, как и проблема эволюции, по тем же самым причинам. Во-первых, никогда нельзя точно знать, что происходило в отдаленные времена. Вероятно, всегда будет изобилие спекуляций относительно обстоятельств появления жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а
9 .2. Происхождение и ранние стадии развития жизни
9.2. Происхождение и ранние стадии развития жизни Современная теория происхождения жизни — это теория появления структур, которые, развиваясь и усложняясь, приобретали определенные функциональные свойства. По-видимому, эти свойства могли возникнуть уже на ранних этапах
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а