5.4. Транспортные системы клеток. Транспортеры
5.4. Транспортные системы клеток. Транспортеры
Перенос веществ через мембрану в большинстве случаев осуществляется с помощью неспецифических и специфических систем активного и пассивного транспорта — переносчиками, каналами и порами. Больших различий между этими понятиями не делается.» В последнее время используется термин «транспортер».
Переносчики и каналы — устройства, обеспечивающие специфические транспортные процессы. В клетках высших организмов, и в том числе в кишечных, существуют транспортеры многих типов. К ним относятся переносчики глюкозы, аминокислот и др. Каждое такое устройство переносит один или ограниченное число типов органических молекул через мембрану по электрохимическому градиенту или благодаря сопряжению с механизмом транспорта другого вещества, движение которого до градиенту концентраций служит источником энергии для сопряженного с ним процесса.
Для вторичной энергизации используются многие ионные градиенты, но преимущественно градиент Na+ без участия АТФ (табл. 8). Na+-зависимые переносчики используются в различных системах, включающих в себя систему всасывания (например, тонкая кишка), систему обеспечения собственных энергетических и пластических потребностей, транспорт ряда веществ (например, тех же глюкозы и аминокислот) для собственных синтезов и т.д. В печени механизмы активного транспорта глюкозы используются для депондрования веществ и их иммобилизации из депо. Мобильный переносчик, представляющий собой, как правило, белковую молекулу, движется от одной поверхности мембраны к другой, совершая вертикальные или вращательные движения, с тем чтобы связывать транспортируемые субстраты на одной поверхности мембраны и освобождать с другой. Типичным примером такого переносчика служат ионофоры. Канал как устройство характеризуется наличием постоянной или индуцированной поры, через которую проходит транспортируемое вещество.
Nа+-зависимый транспорт органических веществ в клетках животных
Вещество Ткани или клетки Глюкоза, галактоза и их дериваты Тонкая кишка, почечные канальцы Аминокислоты Тонкая кишка, почечные канальцы, желточный мешок, мозг, печень, ацинарные клетки поджелудочной железы, жировая ткань, мышцы, клетки асцитной карциномы Эрлиха, клетки карциномы КВ, эритроциты голубя, ретикулоциты, лейкоциты, покровы тела морских беспозвоночных Дипептиды, трипептиды Тонкая кишка Аскорбиновая кислота Тонкая кишка, мозг, надпочечники Биотия, тиамин Тонкая кишка, мозг Соли желчных кислот, билирубин Тонкая кишка ?-Аминогиппуровая кислота Почечные канальцы Лактат Тонкая кишка, почечные канальцы Холин Тонкая кишка Миоинозитол То же Рибофлавин » Урацил »Несмотря на то что механизм Na+-зависимoro транспорта не вполне ясен, ряд фактов свидетельствует о его сходстве или идентичности в разных клетках и органах. Так, не удается выявить переносчиков аминокислот, характерных для клеток одного типа и отсутствующих у других (табл. 9). В пользу идентичности транспортных блоков в разных клетках свидетельствуют генетические дефекты, в том числе точечные. Так, при нарушении всасывания одной из субстанций в кишечных клетках наблюдаются аналогичные нарушения всасывания в клетках почечных канальцев (см. гл. 7). Распространенность таких нарушений говорит, что дефект локализован в одном гене. У человека идентифицировано более 20 «транспортных» болезней. Однако молекулярная природа недостающих компонентов в транспортной системе не ясна. Примеры некоторых генетических нарушений приведены в табл. 10. При дефиците различных транспортных или связывающих белков в кишечных клетках наблюдается малабсорбция глюкозы и галактозы, нейтральных аминокислот, цистина и глицина, фолиевой кислоты и т.д.
Ткани и клетки, для которых характерен Na+-завксимын транспорт аминокислот
Эпителиальные ткани Тонкая кишка (животные различных видов) Почки Желчный пузырь (кролик) Мочевой пузырь земноводных Стенка тела (морские беспозвоночные) Неэпителиальные ткани Жировые ткани Хрусталик глаза Кости и хрящи Печень Мозг Поперечнополосатые мышцы Одиночные клетки Асцитная карцинома Эрлиха Фибробласты мыши Морские бактерии Ретикулоциты (кролик) Эритроциты голубя Лейкоциты (человек)Вместе с тем, как упомянуто выше, наряду с Na+-зависимым транспортом существует Ма+-пезависимый транспорт. Этот тип транспорта обнаружен при изучении переноса через мембрану кишечных клеток моносахаридов, аминокислот и других мономеров, образующихся в результате мембранного гидролиза соответствующих олигомеров.
Таблица 10
Некоторые примеры генетических нарушений транспортных систем у человека
Заболевание Вещество, транспорт которого нар; шен Почечная глюкозурия Сахара Нарушение всасывания в кишечнике глюкозы и галактозы » Цистинурия Аминокислоты Болезнь Хартнупа » Иминоглицинурия Нарушение всасывания витамина В12 Витамин В12 Семейный рахит Фосфат Наследственный сфероцитоз Na+, К+Благодаря распространению Na+-зависимых транспортеров на апикальной мембране, а натриевых насосов — на базолатеральной создается транзитный перенос, необходимый для обеспечения за счет работы каждой кишечной клетки потребностей многих тысяч других соматических клеток. Транспортные функции клеток других типов реализуются практически тем же самым набором функциональных блоков. Действительно, транспортные характеристики клеток тонкой кишки и плаценты во многом совпадают.
Ряд клеток характеризуется тем, что универсальные транспортные блоки расположены на клеточной поверхности иначе. В частности, в ацинарных клетках поджелудочной железы, осуществляющих внешнюю секрецию ферментов, насосы и Na+-зависимые транспортеры расположены в базолатеральной мембране. Этого достаточно для обеспечения градиента Na+ между вне- и внутриклеточной жидкостями и поступлением нутриентов из крови в цитоплазму, а также для энергизации многих других процессов. Кроме того, локализация насосов и каналов в базолатеральной мембране исключает интенсивное поступление потоков воды и солей в апикально-базальном направлении.
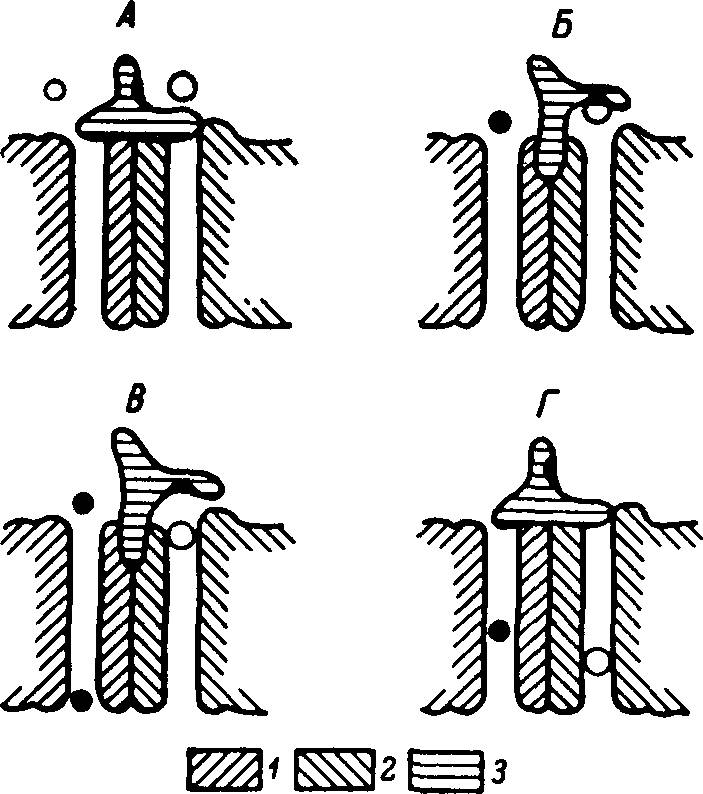
Рис. 28. Схема действия транспортера глюкозы и натрия в плазматической мембране.
А — исходное состояние транспортера: оба канала закрыты; Б — после связывания глюкозы с аллостерическим центром на воротном устройстве: натриевый канал открылся; В — при прохождении натрия по натриевому каналу произошла аллостерическая активация глюкозного канала и молекула глюкозы перешла с воротного устройства на вход своего канала; Г — освобождение центра на воротном устройстве влечет за собой закрытие натриевого и глюкозного каналов. 1 — натриевый канал; 2 — глюкозный канал; 3 — воротное устройство. Светлые пружки — молекулы глюкозы, черные — ионы натрия.
Согласно современным представлениям, при Na+-зависимом транспорте глюкозы через плазматическую мембрану клеток эукариотов происходит образование тройного комплекса: Na+—глюкоза—транспортер. Однако более адекватной представляется наша модель транспортера (рис. 28). Она характеризуется наличием двух параллельно взаимодействующих каналов (для Na+ и для глюкозы) и поверхностного (воротного) белка, связывающего глюкозу на входе в транспортную систему. Транспортный цикл такой модели состоит в следующем: 1) исходно глюкозный и натриевый каналы неактивны; 2) при связывании глюкозы с аллостерическим центром на воротном белке натриевый канал активируется (эта стадия напоминает контроль натриевой проницаемости ацетилхолином) и Na+ движется из экстра- в интрацеллюлярную жидкость; 3) на определенном этапе движения Na+ по каналу происходит аллостерическая активация глюкозного канала; 4) через активированный канал транспортируется молекула глюкозы, первоначально фиксированная в воротном устройстве; 5) освобождение воротного устройства сопровождается дезактивацией натриевого канала. Реактивация последнего происходит при связывании с аллостерическим центром следующей молекулы глюкозы. При температуре 16° С глюкозный канал инактивируется, а натриевый продолжает функционировать. Из нашей модели вытекает, что скорость транспорта глюкозы и Na+ в определенном диапазоне зависит от концентрации Na+ во внеклеточной жидкости. Важно, что Ка+-зависимые транспортеры глюкозы и аминокислот, поскольку показана их идентичность в клетках различных типов и у организмов разных групп, должны соответствовать одной из моделей молекулярных машин.
Насколько можно судить, все разнообразие процессов, связанных с мембранной проницаемостью электролитов и их мембранным транспортом (по крайней мере у всех эукариотов), осуществляется с помощью тех же стандартных функциональных блоков, которые описаны выше. Поразительно, но «примитивные» функции низших организмов и совершенные функции высших организмов реализуются близкими или идентичными функциональными блоками. На уровне элементарных функций организмы, занимающие крайнее положение на вершине и у основания эволюционной лестницы, трудно различимы. Одноименные транспортеры органических субстанций у животных разных видов, по-видимому, также близки или сходны по своей молекулярной структуре и принципам функционирования, хотя этот вопрос изучен меньше, чем в отношении АТФаз. В пользу большой близости этого типа функциональных блоков говорит сходство, а иногда идентичность реакций различных транспортеров на действие ингибиторов и активаторов.
Несмотря на то что сравнительная физиология ионных каналов изучена недостаточно, существуют данные о значительном сходстве их молекулярной структуры и функциональной организации. Вероятно, у млекопитающих и насекомых имеются все основные типы ионных каналов — натриевые, калиевые, хлорные и др.
Далее, у млекопитающих, рыб и насекомых продемонстрировано наличие специализированных натриевых каналов, чувствительных к амилориду, а также каналов, реализующих котранспорт Na+ и Сl-, чувствительных к фуросемиду. Интересно, что те же самые ингибиторы специфически тормозят транспорт ионов в мальпигиевых канальцах и толстой кишке насекомых. Недавно выявлен котранспортер, осуществляющий котранспорт в клетку одного иона Na+, одного иона К+ и двух ионов Сl-. Этот механизм обнаружен в клетках почки и тонкой кишки млекопитающих, в клетках различных органов рыб и насекомых.
До недавнего времени считалось, что функции секреции, экскреции и всасывания имеют глубокие различия, касающиеся механизмов функционирования и эволюционного происхождения. Однако между этими процессами принципиальной разницы не существует. Все основные функциональные блоки, упомянутые выше, лежат и в основе перечисленных функций, в ряде случаев полярных по своему физиологическому значению (например, всасывание и экскреция).
Характеристики ферментных и транспортных систем клеток, выполняющих различные функции (например, всасывание в тонкой кишке, реабсорбция воды в протоках пищеварительных желез и т.д.), очень близки. В апикальной мембране клеток тонкой кишки и почечных канальцев присутствуют трансмембранные ферменты, Ка+-зависимые транспортеры глюкозы, аминокислот и других нутриентов. В базолатеральной мембране этих клеток локализованы близкие или идентичные по молекулярной структуре АТФазы. Системы с множеством транспортеров, локализованных в апикальной мембране, и натриевых насосов, расположенных в базолатеральной мембране, распространены очень широко. В некоторых структурах они выполняют не только резорбтивную, но и секреторную функцию.
Существенно, что при электронной микроскопии многие субклеточные и молекулярные структуры клеток тонкой кишки человека и высших животных неотличимы от таковых клеток кишки рыб, насекомых, моллюсков, круглых червей и т.д., а клетки тонкой кишки — от клеток плаценты, канальцев почек и других клеток.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
1. Детерминанты половых клеток
1. Детерминанты половых клеток Будущие половые клетки — гоноциты иногда можно отличить уже на очень ранних стадиях развития. Классическим примером является аскарида, у которой после первого деления яйца в одном из двух бластомеров теряются концевые участки хромосом и
2. Пол половых клеток
2. Пол половых клеток После того как гоноциты попали в половые железы, их судьба зависит от того, какие половые клетки они должны образовать — мужские, т. е. сперматозоиды, или женские, т. е. яйца. В первом случае их путь развития называют сперматогенезом, а во втором —
1. Форма клеток
1. Форма клеток Форма клеток зависит от их внутренней структуры и свойств клеточной оболочки и от их окружения — соседних клеток и поверхностей контакта. Так, при культивировании отдельных клеток на поверхности стекла все клетки стремятся распластаться по субстрату.
3. Перемещения отдельных клеток
3. Перемещения отдельных клеток Многие процессы формообразования происходят не путем перемещения клеточных пластов, а путем перемещения отдельных клеток. Иногда движение клеточного пласта происходит при частичной потере его эпителиальной структуры. Так, например,
«Государство клеток»
«Государство клеток» Сходная точка зрения, акцентирующая, пожалуй, в еще более категорической форме приоритет части по отношению к целому, была высказана выдающимся немецким биологом Рудольфом Вирховом. В конце 50-х годов XIX века он выступил со своей концепцией, вошедшей
Два главных органа иммунной системы вырабатывают два сорта клеток: Т–и В–лимфоциты.
Два главных органа иммунной системы вырабатывают два сорта клеток: Т–и В–лимфоциты. — Следовательно, иммунная система каждого организма имеет два главных органа, не так ли? — Так? — Один, тимус, заведует выработкой лимфоцитов, способных превращаться в
§ 8. Заряды мембраны нервных клеток
§ 8. Заряды мембраны нервных клеток Однако основные свойства нервной системы обусловлены способностью быстро реагировать на изменение ситуации внутри или вне организма. Скоростные процессы не могут осуществляться по медленным гуморальным законам, они происходят по
7.8. Сигналы нервных клеток
7.8. Сигналы нервных клеток Нервная система возникает в ходе эволюции для анализа организмами поступающей информации. С этой целью нервные клетки используют электрические и химические сигналы. Электрические сигналы бывают двух видов.Градуальный потенциал возникает в
5.3. Транспортные системы клеток. Насосы
5.3. Транспортные системы клеток. Насосы Под насосами подразумеваются системы, в которых сочетаются механизмы энергизации и трансмембранного переноса. Принимается, что источником энергии в большинстве случаев служит энергия макроэргических связей АТФ. Деятельность
2.7. Попытки моделирования примитивных клеток
2.7. Попытки моделирования примитивных клеток В ранних работах о происхождении жизни была выдвинута идея предклеточных структур, существующих в водной среде в форме “микросфер” (Fox, 1965; 1991) и “коацерватных капель” (Опарин, 1966). Микросферы формировались из т. н.
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных