4.3. Примеры деятельности пищеварительной системы у животных, стоящих на разных ступенях эволюции
4.3. Примеры деятельности пищеварительной системы у животных, стоящих на разных ступенях эволюции
На примерах пищеварения у животных, стоящих на разных ступенях эволюции, можно проследить его постепенную эволюцию, а также адаптацию клеток и региональную специализацию функций в пределах пищеварительной системы. При этом основные характеристики процессов гидролиза пищевых веществ по существу являются одинаковыми у животных всех групп.
4.3.1. Пищеварение у простейших
Этот процесс рассмотрим на примере парамеций, пищей которых служат бактерии, одноклеточные водоросли, органические остатки. У парамеций при питании происходит образование вакуолей, которые, подвергаясь конденсации и слиянию, проделывают сложный путь внутри одноклеточного организма, а затем снова сливаются с клеточной мембраной вблизи цитостомы (рис. 22). С помощью движения ресничек вдоль орального отверстия пища заглатывается в пищевод (цитофаринкс), слепая оконечность которого (цитостома) образует карман, где она накапливается. Этот мешок в виде пищевой вакуоли отделяется от цитостомы, а на его месте начинает образовываться другой карман. Пищевая вакуоль, проделав определенный путь вокруг органелл одноклеточного организма, возвращается к точке, близкой к цитостоме. Во время этого цикла содержимое вакуоли гидролизуется и растворимые продукты всасываются в окружающую цитоплазму. Непереваренные остатки пищи выводятся наружу через анальную пору, открывающуюся вблизи цитостомы.
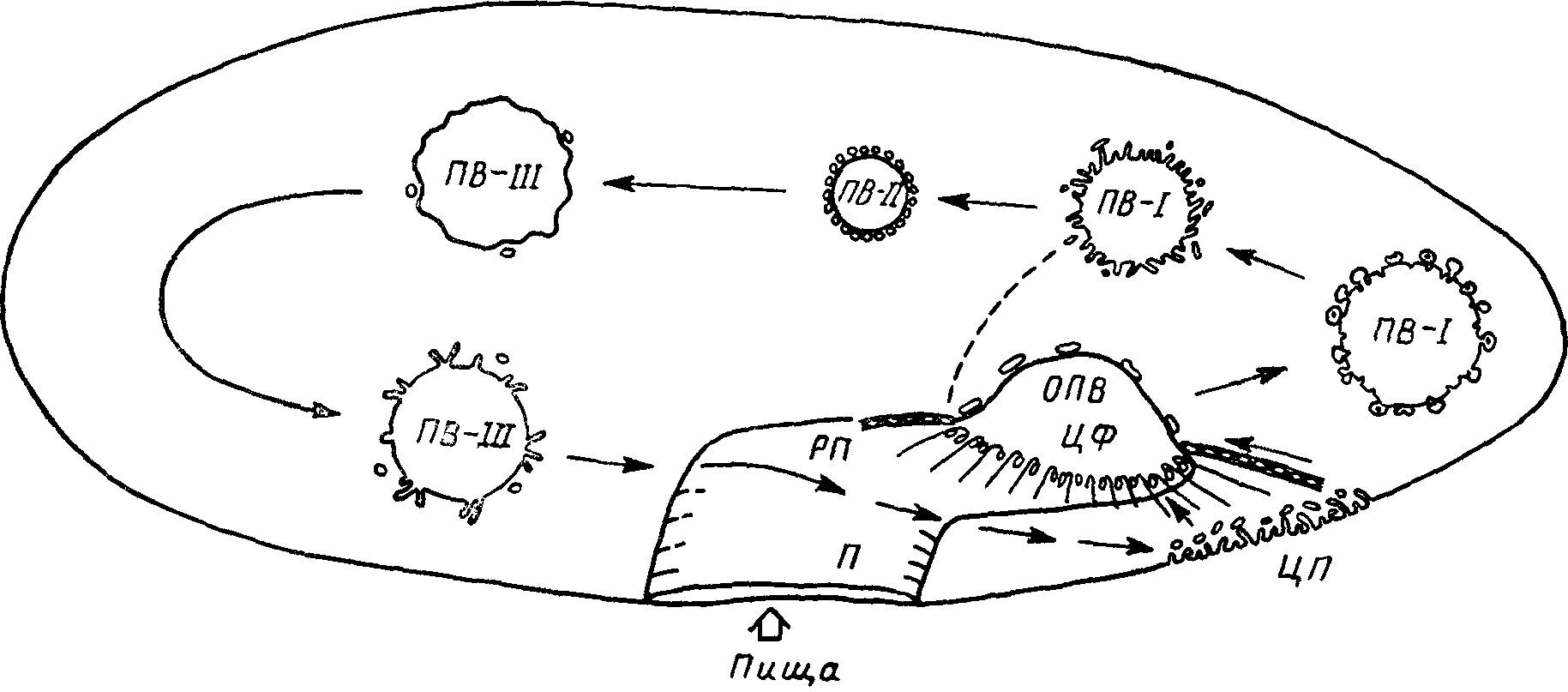
Рис. 22. Пути потока мембран во время эндоцитоза у парамеции.
Пища проникает в преддверие (II) ротовой полости (РП); образующаяся пищеварительная вакуоль (ОПВ) переносится в цитофаринкс (ЦФ). Пищеварительная вакуоль ПВ-I соединяется с везикулами и затем конденсируется, образуя пищеварительную вакуоль ПВ-II с высокой кислотностью. Вакуоль ПВ-II сливается с лизосомами, образуя пищеварительную вакуоль ПВ-III, которая подвергается зкзоцитозу в цитопрокте (ЦП).
Выйдя из фаринкса, пищевая вакуоль благодаря всасыванию воды в цитоплазму уменьшается в размерах, ее содержимое становится более компактным, а pH снижается до 1.4. Однако неизвестно, является ли такое снижение pH результатом истинной секреции кислоты, аутолиза или даже всасывания щелочного материала, заключенного в вакуоли. По-видимому, правильна первая точка зрения. Почти не имеется данных о пищеварении, происходящем в кислой среде. Однако эта фаза играет важную роль в умерщвлении жертвы и денатурации ее белков, как это имеет место у многих других животных. Хотя она и не гомологична первой фазе пищеварения у высокоорганизованных животных, тем не менее ее можно рассматривать как предвестник действительно кислотной фазы гидролиза.
Уменьшение размеров вакуоли является временным; в конце концов она вновь увеличивается, однако ее диаметр никогда не достигает исходной величины. При этом pH содержимого быстро повышается до 7.5—8.0, что, по-видимому, вызвано секрецией в вакуоль щелочных соков из окружающей цитоплазмы. Секрет, поступающий в вакуоль во время щелочной фазы, содержит протеазы, карбогидразы и липазы. Протеазы сходны с трипсином хордовых тем, что они активны в щелочной среде; пепсинподобных ферментов не обнаружено. Пищеварение в щелочной среде происходит очень быстро. По мере пищеварения, а также всасывания продуктов гидролиза и воды вакуоль уменьшается в размерах и, достигнув области анальной поры, содержит только непереваренные элементы. Имеются данные, что пищеварение (по крайней мере протеолиз) у парамеций не заканчивается в пищевой вакуоли, а продолжается в цитоплазме за счет содержащихся в ней катепсинов и дипептидаз.
У простейших других групп последовательность событий аналогична. В экстрактах цитоплазмы различных амеб обнаружены эндопептидазы, дипептидазы, карбогидразы, липазы, щелочная фосфатаза, часть которых, вероятно, секретируется в вакуоли. Однако, как и в случае парамеций, о месте синтеза и последовательности образования этих гидролаз известно мало.
4.3.2. Пищеварение у плоских червей
Для свободноживущих плоских червей характерно внутриклеточное пищеварение. Однако большинство животных этого типа способно и к внеклеточному пищеварению, а их пищеварительная система обладает признаками клеточной и региональной специализации в этом направлении. У большинства плоских червей передняя кишка специализирована для питания и заглатывания пищи, а пищеварение происходит в результате сочетания внутриклеточных и внеклеточных процессов.
Пищеварение может быть охарактеризовано на примере турбеллярий. Как водные, так и наземные животные являются плотоядными. Они захватывают пищу (олигохеты, ракообразные, насекомые, моллюски или водоросли) и удерживают ее, пока фаринкс не проникнет в тело жертвы. В этот момент железистые клетки фаринкса начинают секретировать эндопептидазу, способствующую проникновению фаринкса и расщеплению тела добычи, всасываемого фаринксом. При поступлении пищи в отдел, следующий за фаринксом, железистые клетки секретируют эндопептидазу с оптимумом pH 5.0, участвующую во внеклеточном пищеварении. Этот фермент сходен (если не идентичен) с эндопептидазой фаринкса. Он также во многих отношениях сходен с пепсином хордовых, за исключением того, что эндопептидаза активируется веществами, содержащими сульфгидрильные группы, и, следовательно, по этому показателю больше похожа на внутриклеточную эндопептидазу хордовых. Первая внеклеточная фаза гидролиза происходит в кислой среде и сводится к протеолизу. Однако неизвестно, идет ли он до конца и образуются ли при этом дипептиды и аминокислоты. Эндопептидаза расщепляет часть белков до полипептидов, что в конечном итоге превращает пищу в однородную массу, суспензированную в жидкостях тела самой добычи. Кроме внеклеточного пищеварения, в этот период отмечается значительный фагоцитоз и со временем вся пища переходит в фагоцитарные клетки.
Фагоцитоз осуществляется свободными дистальными краями клеток, поглощающих частицы подобно амебам. Первоначально довольно большие вакуоли в процессе внутриклеточного гидролиза (за счет всасывания воды в цитоплазму) уменьшаются, а содержимое их становится компактней и однородней. Отшнуровываясь, вакуоли двигаются в глубь клетки, а на их месте образуются новые.
Последовательность внутриклеточного гидролиза позволила идентифицировать и локализовать типы соответствующих ферментов. При всасывании воды из вакуоли в нее из цитоплазмы поступает кислый секрет, содержащий активную эндопептидазу. Последняя продолжает протеолиз, начавшийся внеклеточно эндопептидазой железистых клеток. Одновременно в цитоплазме и в меньшей степени в вакуолярной жидкости появляется кислая фосфатаза, возможно, имеющая отношение к расщеплению фосфатсодержащих компонентов пищи. Но более вероятно, что она каким-то образом связана с выработкой эндопептидазы (по-видимому, как компонент системы освобождения энергии). Действие этих ферментов может продолжаться в течение 18 ч после приема пищи. При этом белки расщепляются до полипептидов, а стенки клеток, ядерные мембраны и другие плазматические структуры, состоящие в основном из белков, растворяются, так что жиры и углеводы становятся доступными соответствующим ферментам.
К концу первой фазы внутриклеточного пищеварения действие эндопептидазы прекращается, а активность кислой фосфатазы уменьшается. Далее наступает вторая (щелочная) фаза пищеварения. Благодаря секреции щелочных соков pH содержимого вакуоли повышается до 7.0—7.5 и активность кислой фосфатазы как в цитоплазме, так и в вакуолярной жидкости сменяется щелочнофосфатазной. Возможно, на более поздних этапах внутриклеточного гидролиза она участвует также во всасывании продуктов реакции. Выделяемые в вакуоль экзопептидазы завершают протеолиз полипептидов, образовавшихся под действием эндопептидазы. Жировые и углеводные компоненты пищи гидролизуются липазами и карбогидразами соответственно.
Продукты внутриклеточного гидролиза всасываются из вакуоли в цитоплазму фагоцитарных клеток и оттуда с помощью диффузии переносятся к остальным частям тела, так как у турбеллярий нет сосудистой системы — обычной транспортной среды тела животного. Кишка у них разделена на сильно разветвленные переднюю и две задние ветви, так что пищеварительная область охватывает все тело. В итоге все ткани или органы находятся достаточно близко от места пищеварения даже у тех тропических видов, тело которых достигает значительных размеров. Непереваренные остатки пищи движениями цитоплазмы выпятится обратно к свободному дистальному краю фагоцитарных клеток. Здесь они выталкиваются в полость, присоединяются к остаткам внеклеточного пищеварения и выделяются наружу через ротовое отверстие, так как анальное отверстие у плоских червей отсутствует.
Анализ имеющихся сведений о пищеварении у организмов, стоящих на разных уровнях эволюционного развития, в том числе у млекопитающих, позволил прийти к следующему заключению. Несмотря на различия в организации животных, относящихся к разным типам, процессы пищеварения у них характеризуются общими чертами. В самом деле, схема И. И. Мечникова «эволюция от внутриклеточного пищеварения к внеклеточному» могла рассматриваться как эволюция от простого к сложному. Однако с использованием методов ультрамикроанализа показано, что как у наиболее высокоорганизованных, так и у простейших животных существует общая последовательность деградации пищевых веществ: первоначальный гидролиз белков в кислой среде в отсутствие всасывания. У высших животных и человека пища из желудка поступает в тонкую кишку, где pH из кислого становится нейтральным или слабощелочным и инициируется гидролиз основных групп пищевых веществ за счет ряда соответствующих ферментов. Этот процесс совмещается с интенсивным всасыванием. У простейших, хотя весь цикл переваривания осуществляется в одной вакуоли, на определенном этапе также происходит сдвиг pH в щелочную сторону, появление в вакуоли различных ферментов, сходных с теми, которые свойственны высшим организмам, а также интенсификация всасывания. Наконец, электронно-микроскопический анализ демонстрирует, что механизмы секреторного процесса, в частности у тетрахимены, напоминают таковые у высших животных, например в клетках поджелудочной железы млекопитающих. Таким образом, у простейших не только доказана секреция, но и сходство этого механизма с аналогичным механизмом высших организмов. Характеристика кишечных эпителиев и секреторных процессов у низших животных весьма существенна, так как кишечные эпителии этих организмов являются филогенетическими предшественниками секреторных эпителиев таких важных органов высших организмов, как поджелудочная железа, печень и тонкая кишка.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 1.4. Отбор животных по признаку уровня рассудочной деятельности
Глава 1.4. Отбор животных по признаку уровня рассудочной деятельности Значение изучения рассудочной деятельности животных трудно переоценить как с научной, так и с этической точки зрения. Большой вклад в изучение этой проблемы внес Л. В. Крушинский и его сотрудники
8.3. Основные тенденции эволюции нервной системы позвоночных
8.3. Основные тенденции эволюции нервной системы позвоночных Позвоночные обычно рассматриваются как подтип Vertebrata в типе хордовых (Chordata). Они представлены бесчелюстными, рыбами, амфибиями, рептилиями, птицами и млекопитающими. В систематике позвоночных много спорных
Глава 9 ИЗМЕНЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА
Глава 9 ИЗМЕНЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА Гибкий и подвижный механизм высшей нервной деятельности обнаруживает громадные приспособительные возможности нормального функционирования при самых разных состояниях организма.Знание
Особенности наблюдения и лечения животных с заболеваниями костно- мышечной системы
Особенности наблюдения и лечения животных с заболеваниями костно- мышечной системы Большинство заболеваний органов движения у животных (за исключением травматических повреждений) имеют относительно доброкачественное течение и не представляют собой непосредственной
Направления эволюции нервной системы
Направления эволюции нервной системы Мозг – структура нервной системы. Появление нервной системы у животных давало им возможность быстро адаптироваться к меняющимся условиям среды, что, безусловно, можно рассматривать как эволюционное преимущество. Общей
Ранние представления о психической деятельности животных
Ранние представления о психической деятельности животных Начало познания поведения животных При изучении любой формы психической деятельности прежде всего встает вопрос о врожденном и индивидуально приобретаемом, об элементах инстинкта и научения в поведении
Высшие формы игровой исследовательской деятельности животных
Высшие формы игровой исследовательской деятельности животных При всем многообразии форм игры их объединяет большая общая подвижность животного, большое разнообразие производимых им телодвижений и интенсивное перемещение в пространстве (рис. 23). Наиболее ярко это
ЕСТЕСТВЕННОНАУЧНОЕ ИССЛЕДОВАНИЕ ТАК НАЗЫВАЕМОЙ ДУШЕВНОЙ ДЕЯТЕЛЬНОСТИ ВЫСШИХ ЖИВОТНЫХ [ 2 ]
ЕСТЕСТВЕННОНАУЧНОЕ ИССЛЕДОВАНИЕ ТАК НАЗЫВАЕМОЙ ДУШЕВНОЙ ДЕЯТЕЛЬНОСТИ ВЫСШИХ ЖИВОТНЫХ[ 2 ] Частным предметом моего сегодняшнего чтения, посвящаемого памяти великого естествоиспытателя и энергичнейшего борца за величайшее биологическое учение — учение о развитии —
ОБЪЕКТИВНОЕ ИЗУЧЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ [ 20 ]
ОБЪЕКТИВНОЕ ИЗУЧЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ[ 20 ] Второй раз я имею высокую честь и глубокое удовлетворение принимать участие в торжестве открытия деятельности чрезвычайных русских обществ, народившиеся здесь, в Москве, и свидетельствующих о том, как
ФИЗИОЛОГИЯ И ПСИХОЛОГИЯ ПРИ ИЗУЧЕНИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ [ 25 ]
ФИЗИОЛОГИЯ И ПСИХОЛОГИЯ ПРИ ИЗУЧЕНИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ[ 25 ] Прежде всего я считаю своим долгом благодарить Философское общество, что оно в лице своего председателя изъявило готовность выслушать мое сообщение. Мне трудно было сообразить, насколько это
ОБЩИЕ ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ И ЧЕЛОВЕКА1
ОБЩИЕ ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ И ЧЕЛОВЕКА1 Образы, картины поведения как нас самих, так и близких к нам высших животных, с которыми мы находимся в постоянных жизненных отношениях (как, например, собак), представляют огромное разнообразие, прямо
5.5. О значении элементов случайности на разных этапах клеточной эволюции
5.5. О значении элементов случайности на разных этапах клеточной эволюции Мы рассмотрели основные этапы химической, предклеточной и клеточной эволюции вплоть до формирования эукариотических клеток, основавших царства животных и растений. Это сделано, в частности, для