6.2. Формирование универсальных функциональных блоков в филогенезе и механизмы их консервации
6.2. Формирование универсальных функциональных блоков в филогенезе и механизмы их консервации
Мы подошли к основному вопросу, касающемуся формирования универсальных функциональных блоков. С одной стороны, в условиях дивергентной эволюции свойства функциональных блоков, возникающих на ранних стадиях филогенеза из общего предшественника, должны все более и более различаться. С другой стороны, сходство одноименных блоков у представителей филогенетически далеких таксонов поразительно. Допустим, что существует постоянная тенденция к изменению свойств функциональных блоков вследствие различных причин, в том числе нейтральных мутаций. Мутационный процесс лишь частично нивелируется последующим действием естественного отбора, в результате которого элиминируются мутации с вредными селективными признаками. Многие мутации, охарактеризованные как нейтральные или, что более правильно, близкие к нейтральным, могут накапливаться.
Необходимо допустить существование специального механизма, стабилизирующего некоторые важные свойства функциональных блоков, что обеспечивает их сохранение в течение длительных интервалов времени и, следовательно, сходство со свойствами исходной структуры одноименных блоков в других таксонах. Мною было высказано предположение, что таким механизмом может быть стабилизирующее действие отбора в отношении нескольких функционально и структурно взаимосвязанных признаков, мутирующих независимо. Эта точка зрения получает подтверждение (табл. 15). Как можно видеть, частота эволюционных изменений структуры макромолекул варьирует в пределах трех и более порядков, хотя вероятность мутаций должна быть одинаковой или близкой на протяжении длительных интервалов времени. Различия возникают вследствие действия дополнительных механизмов, из которых хотелось бы уделить внимание связи между стабилизирующим действием естественного отбора и структурно-функциональными интеграциями. Фактором, стабилизирующим свойства функциональных блоков в ходе эволюции, возможно, является стабилизирующий отбор тех признаков, которые сопряжены, во-первых, со стабильностью свойств лиганда и, во-вторых, с высокой вероятностью превращения полезного признака во вредный при взаимодействии двух функциональных блоков или комплексов, контролируемых независимо мутирующими генными структурами.
Таблица 15
Скорость эволюции макромолекул
Макромолекулы Скорость эволюции, выраженная в замещениях на кодон за 1010 лет Гипервариабельные нуклеотидные последовательности ДНК 500 «Средняя» ДНК млекопитающих 48-50 мРНК цепей гемоглобина 100 мРНК гистона IV у морского ежа разных видов 35 «Средние» белки 1-50 Гемоглобин, а- и ?-цепи соответственно 14 Гистон IV 0.09 Цитохром с митохондриальный 5 Гипервариабельные аминокислотные последовательности белков 85Идея стабильности некоторых структур в ходе эволюции не нова. В настоящее время она общепризнанна для информационных блоков. В сущности вся эволюционная генетика и теория эволюции живых систем опираются на эти представления. Кроме того, на идее стабильности базируются представления об эволюции на основе строительных блоков — аминокислот, оснований нуклеиновых кислот, моносахаридов и т.д.
Следует обратить внимание на большую эволюционную стабильность олигомерных белков по сравнению с мономерными. Взаимодействие субъединиц в олигомерных белках — существенный селективный признак, который, по-видимому, стабилизируется естественным отбором. Отсюда полиморфизм у мономерных ферментов выражен больше, чем у олигомерных. Благодаря естественному отбору реализуется контроль активных центров как мономерных, так и олигомерных ферментов, а также взаимодействующих участков в субъединицах олигомерных ферментов. К. Райдер и К. Тейлор (1983) заключают, что большая степень полиморфизма у мономерных ферментов — аргумент скорее в пользу гипотезы нейтральных мутаций, чем в пользу гипотезы отбора. Однако в действительности должны учитываться и обе эти гипотезы, и функциональная анатомия макромолекул. Особенно четко это видно при рассмотрении эволюционной изменчивости различных участков полипептидной цепи инсулина. Многими показано, что нефункционирующий С-фрагмент инсулина характеризуется в 10 раз большей мутабельностью, чем А- и В-фрагменты, хотя они кодируются одним геном. Лишь последние взаимодействуют с рецептором, выполняя функцию химического сигнала. На рис. 34 и 35 представлены две модели взаимодействия инсулина и рецептора. По-видимому, происходящие от общего предшественника гастрина и холецистокипина С-концевые фрагменты пептидной цепи, выполняющие регуляторную функцию, в ходе эволюции не изменились. Что касается ферментов, то многочисленные данные свидетельствуют, что в течение длительных интервалов времени в ходе эволюции может меняться структура фермента, но оставаться неизменным его каталитически активный центр.
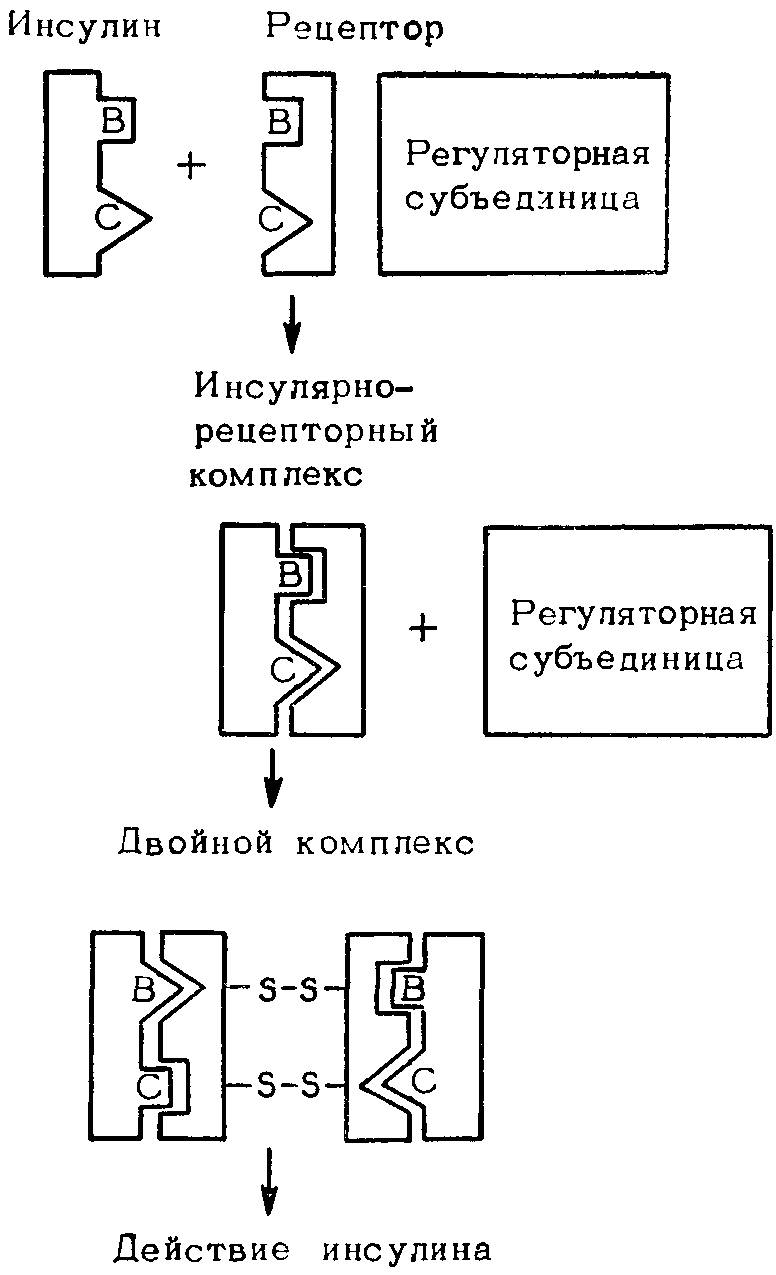
Рис. 34. Модель взаимодействий инсулина и рецептора с отдельными доменами молекулы инсулина для связывания и биологической активности.
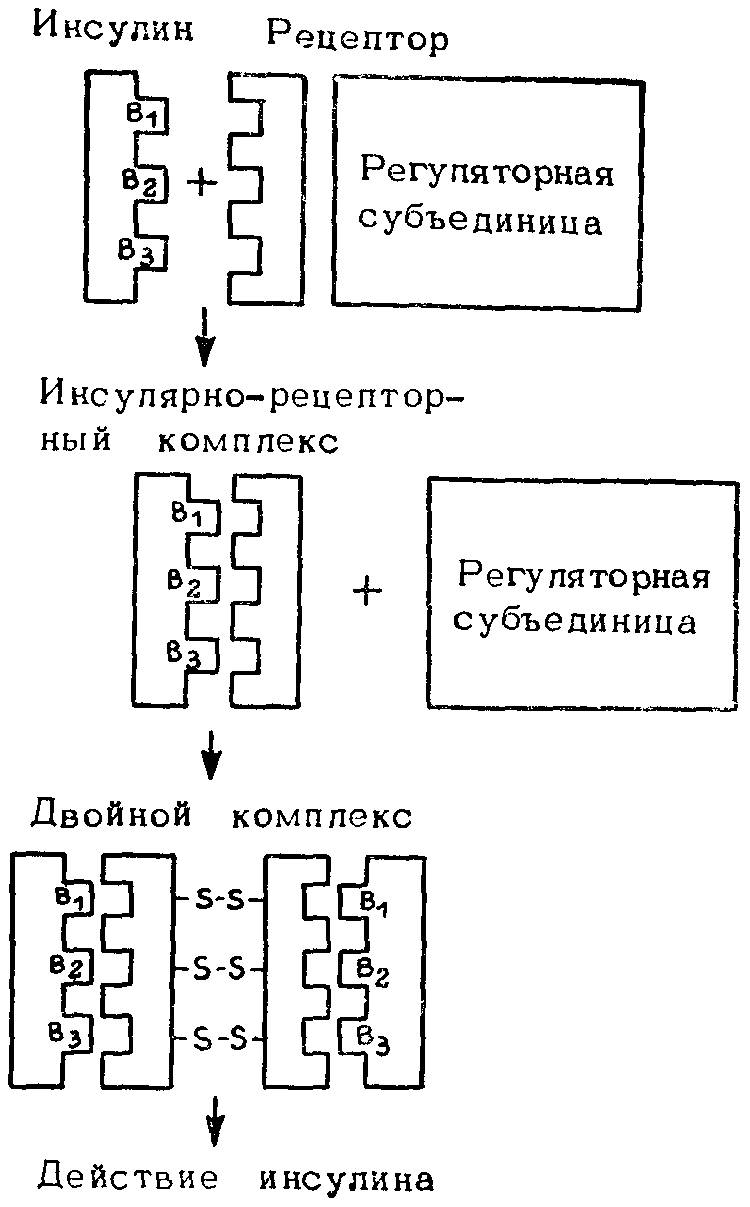
Рис. 35. Модель взаимодействий инсулина и рецептора бее отдельных доменов для связывания и биологической активности.
Следовательно, в ходе эволюции стабилизированными, консервативными являются, как правило, не макромолекулы в целом, а их работающие части, выполняющие специфические функции, особенно такие, как взаимодействие с другими элементами. Так, при взаимодействии пептидного гормона с рецептором значимая мутация в одном из двух независимых генов
привела бы к возникновению несоответствия между рецептором и гормоном. Во многих случаях это означало бы появление генетической болезни, результатом которой было бы снижение вероятности выживания ее носителей или их гибель (при тяжелой форме заболевания). С нашей точки зрения, подобные феномены в ходе эволюции происходили постоянно, но не оставляли следов, так как последствия таких мутаций, нарушавших жизненно важные взаимодействия между гормоном и рецептором, между субъединицами олигомеров и т.д., были элиминированы. Я предположил, что о правильности такого построения свидетельствовало бы существование заболевания эволюционным, или генетическим, диабетом, в частности с точечными значимыми мутациями в А- и В-цепях инсулина. Недавно это предположение получило подтверждение.
Несколько лет назад был выделен структурно-аномальный инсулин из сыворотки крови и поджелудочной железы человека, страдающего диабетом. Позднее обнаружено еще два случая диабета со сходными клиническими проявлениями. У всех трех больных имела место типичная для диабетических больных гипергликемия и выраженная гиперинсулинемия, характерная для инсулярной резистентности, но в то же время гормональная толерантность к экзогенному инсулину. Анализ показал, что инсулин сыворотки крови состоит из аномального инсулина, обладающего лишь 1—2% активности нормального гормона, а также нормального инсулина в молярном отношении 95 :5 соответственно. Высокий общий уровень инсулина в сыворотке больных объясняется медленным удалением аномального гормона из системы циркуляции. Авторы указывают, что различные варианты инсулина при таких формах диабета могут быть результатом мутаций в различных местах в пределах гена инсулина и приводить к многочисленным замещениям аминокислот.
Точечная мутация в гене, контролирующем синтез какого-либо пептидного гормона, например инсулина, может приводить к формированию лиганда, плохо взаимодействующего со своим рецептором. В терминах клинической эндокринологии после такой мутации будет формироваться эволюционный, или генетический, диабет. По крайней мере некоторые формы диабета, реально обнаруженного в настоящее время, являются результатом нарушенного соответствия между лигандом и рецептором и отражают одно из самых значимых эволюционных ограничений. В этом отношении важны данные А. И. Клиорина, показавшего, что в крови детей в начальной фазе заболевания сахарным диабетом инсулин содержится в значительных количествах, но он малоэффективен.
Исходя из концепции универсальных функциональных блоков, следует предположить, что подобно эволюционному диабету должны существовать некоторые другие формы эволюционной патологии, в частности связанной с мутациями в генах, контролирующих синтез соматотропного гормона и некоторых других регуляторных пептидов.
Современный функциональный подход дает возможность глубже понять эволюцию вообще и прогрессивную эволюцию в частности, а также качественные скачки — ароморфозы по терминологии А. Н. Северцова. Еще недавно допускалось, что ароморфоз на молекулярном и близком к нему уровнях организации живых систем связан с повышением эффективности деятельности молекулярных машин. Например, ряд крупных биохимиков Советского Союза и других стран полагали, что в ходе эволюции эффективность ферментативно активных белков увеличивается. В настоящее время принято, что ферменты, участвующие во взаимодействиях с внешней средой, более изменчивы, чем ферменты, реализующие различные этапы собственно клеточных процессов. Для того чтобы рассмотреть этот вопрос более подробно, наиболее подходящими кажутся пищеварительные ферменты, например амилаза. Этот фермент характерен как для млекопитающих, так и для бактерий и обнаружен у всех промежуточных форм.
Анализируя совокупность приведенных в литературе и собственных экспериментальных данных, еще в 1961 г. я пришел к выводу, что нет оснований говорить о совершенствовании этого фермента в ходе эволюции. В 1986 г., наконец, появились обобщающие результаты, полученные при изучении первичной структуры слюнной и панкреатической амилаз различных млекопитающих, а также амилазы ряда низших организмов. Анализ последовательности аминокислот показал несомненную гомологичность этих ферментов. Несмотря на то что условия функционирования амилазы и способы регулирования ее синтеза в ходе эволюции менялись неоднократно, принципиальных изменений в структуре этого фермента обнаружено не было. Эти результаты — чрезвычайно важный аргумент в пользу идеи относительно большой эволюционной устойчивости функциональных блоков. Не менее важно, что следствием такой устойчивости является не прекращение эволюции, а ее ускорение за счет различных пространственных сочетаний и комбинаций одних функциональных блоков с другими. Вместе с тем существует возможность использования организмом регуляторных путей. Теоретически ясно, что многие мутации в исполнительных системах связаны не только с изменением их эффекторных свойств, но и регуляторных характеристик, т.е. с выходом из-под контроля имеющихся в организме регуляторных систем.
Интересно, что дупликация гена, уход его из некоторого функционального комплекса, с которым он взаимодействовал, нередко приводит к потере стабилизирующего действия естественного отбора и в конечном итоге — к формированию многих существенных различий. Слюнная амилаза, по всей вероятности, является таким продуктом дуплицированного гена, имеющего ряд принципиальных отличий от гена панкреатической амилазы. В то же время все другие амилазы организма, в том числе амилаза печени, практически полные генетические копии панкреатической амилазы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
4. СОН В ОТНО- И ФИЛОГЕНЕЗЕ
4. СОН В ОТНО- И ФИЛОГЕНЕЗЕ В онтогенезе соотношение «сон–бодрствование» изменяется. Так, у новорождённых состояние бодрствования составляет лишь незначительную часть суток, и значительную часть сна занимает быстрый сон. По мере взросления уменьшается общее количество
2. ТЕОРИЯ ФУНКЦИОНАЛЬНЫХ СИСТЕМ
2. ТЕОРИЯ ФУНКЦИОНАЛЬНЫХ СИСТЕМ 2.1. Что такое система? Термин «система» обычно применяется для того, чтобы указать на собранность, организованность группы элементов и отграниченность её от других групп и элементов. Давалось множество определений системы, которые
Из какого количества структурно-функциональных единиц состоят фасеточные глаза насекомых?
Из какого количества структурно-функциональных единиц состоят фасеточные глаза насекомых? Структурно-функциональную единицу фасеточного глаза насекомых и некоторых других беспозвоночных называют оммадитием. Каждый оммадитий состоит из роговицы, хрусталика и нервных
Глава 8 Гомеозис в онтогенезе и филогенезе
Глава 8 Гомеозис в онтогенезе и филогенезе Смотри! Смотри! Что же я вижу? Лошадиную голову Там, где должен быть хвост! Руби Гольдберг
Формирование пола
Формирование пола Процесс формирования пола занимает длительное время, и принципиальным следствием такого процесса являются глубокие различия между мужскими и женскими организмами, которые невозможно изменить декларативно или с помощью пластической операции на
5.1. Концепция универсальных функциональных блоков. Общие положения
5.1. Концепция универсальных функциональных блоков. Общие положения Суть концепции, охарактеризованной мною в 1985 г., сводится к следующему.1. Различные функции, в том числе специализированные, выполняемые клетками различных тканей и органов высших организмов,
5.6. Концепция универсальных функциональных блоков и кишечная гормональная система
5.6. Концепция универсальных функциональных блоков и кишечная гормональная система Недавно, когда среди гормонов, продуцируемых эндокринными клетками тонкой кишки, оказались гормоны, характерные для гипоталамуса, гипофиза, щитовидной железы, коры надпочечников и т.д.,
6.1. Основные принципы эволюции в свете концепции универсальных функциональных блоков
6.1. Основные принципы эволюции в свете концепции универсальных функциональных блоков В какой степени закономерности и принципы функционирования организмов, органов и клеток справедливы для молекулярных и близких к ним структурно-функциональных систем, какими являются
6.4. Концепция универсальных функциональных блоков и экология
6.4. Концепция универсальных функциональных блоков и экология Анализ структуры различных экосистем и взаимоотношений ее членов может дать новые доказательства правильности концепции универсальных функциональных блоков. Жизнь возможна лишь как циркуляция веществ и
Глава 7 НЕКОТОРЫЕ СЛЕДСТВИЯ КОНЦЕПЦИИ УНИВЕРСАЛЬНЫХ ФУНКЦИОНАЛЬНЫХ БЛОКОВ
Глава 7 НЕКОТОРЫЕ СЛЕДСТВИЯ КОНЦЕПЦИИ УНИВЕРСАЛЬНЫХ ФУНКЦИОНАЛЬНЫХ БЛОКОВ Каждый функциональный блок (или ограниченное число блоков, так называемые изоблоки), реализующий определенную элементарную функцию, универсален в том смысле, что выполняет соответствующую
7.1. «Синдром общих блоков»
7.1. «Синдром общих блоков» Концепция универсальных функциональных блоков вводит нас в новую область медицины, относящуюся к молекулярным заболеваниям. В настоящее время уже обнаружены заболевания, которые должны быть истолкованы с позиций этой концепции. Такие
7.4. Адаптационно-компенсаторные реакция на уровне функциональных блоков
7.4. Адаптационно-компенсаторные реакция на уровне функциональных блоков Рассмотрим три примера развития адаптационно-компенсаторных реакций на основе концепции универсальных функциональных блоков.Адаптация поджелудочной железы к качеству пищи. Способность
СООБЩЕСТВО: ФОРМИРОВАНИЕ
СООБЩЕСТВО: ФОРМИРОВАНИЕ Как из всех присутствующих в данном географическом регионе видов формируется сообщество? Является ли сообщество просто «сборищем» всех возможных для данного типа местности видов или его образование подчиняется каким-то правилам комбинации
1.4. Универсальность строительных и функциональных блоков на различных уровнях организации биологических систем как условие динамического и трофического единства биосферы
1.4. Универсальность строительных и функциональных блоков на различных уровнях организации биологических систем как условие динамического и трофического единства биосферы Длительное существование биосферы как динамической системы невозможно без циркуляции веществ и
1.4. Универсальность строительных и функциональных блоков на различных уровнях организации биологических систем как условие динамического и трофического единства биосферы
1.4. Универсальность строительных и функциональных блоков на различных уровнях организации биологических систем как условие динамического и трофического единства биосферы Длительное существование биосферы как динамической системы невозможно без циркуляции веществ и