4.5. Теории происхождения внешнейи внутренней секреции
4.5. Теории происхождения внешнейи внутренней секреции
Существует две основные теории происхождения секреции. Рассмотрим их отдельно.
4.5.1. Теория происхождения внешней секреции Иордана
Эта теория рассматривает пути гипотетического перехода от внутриклеточного пищеварения к внеклеточному. X. Иордан полагал, что высокоспециализированная голокриновая секреция ферментов и других биологически активных веществ в полость пищеварительного тракта возникла лишь у многоклеточных организмов на сравнительно поздних этапах эволюции. Первоначально пищеварение в кишечной полости происходило под влиянием ферментов, поступающих в просвет кишки в результате естественного разрушения и отмирания части эпителиальных клеток и последующей солюбилизации внутриклеточных ферментов при деструкции этих клеток (рис. 24). По мнению Иордана, такая форма пищеварения была более выгодна для организма, чем внутриклеточное пищеварение. Он назвал эту форму секреции морфокинетической, так как выделение пищеварительных ферментов и секретов связано с изменением структуры клеток. В отличие от этого я предложил назвать такую форму секреции морфонекротической, так как поступление ферментов в полость кишки связано с гибелью клеток. Таким образом, согласно Иордану, наиболее древним типом пищеварительных соков была цитоплазма отторгаемых клеток.
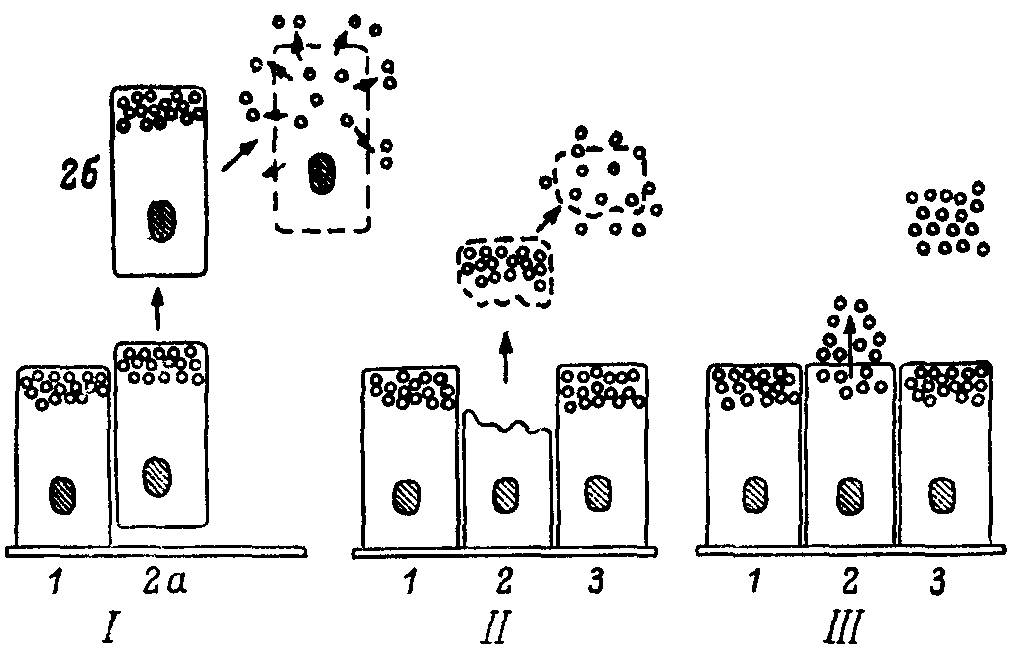
Рис. 24. Схематическая интерпретация перехода от морфокинетической к морфостатической секреции в ходе эволюции согласно теории Иордана. 1—морфонекротическая (голокриновая), II—морфокинетическая (апокриновая) и III — морфостатическая (мерокриновая) стадии секреции. 1—з — последовательные состояния секреторной клетки при разных типах секреции; а, б — этапы выхода ферментов в полость пищеварительного аппарата.
Следующей ступенью эволюции секреторного процесса являлось отторжение лишь апикальной части клетки с ее последующей регенерацией. Я полагаю, что этот апокриновый тип секреции и является действительно морфокинетическим. Дальнейшее развитие секреторного процесса заключалось в уменьшении структурных нарушений клетки при секреции, что характерно для морфостатической (мерокриновой) секреции высших позвоночных. Можно представить себе, что в конце концов должен возникнуть такой тип клеток, секреторная деятельность которых протекает без значительного изменения их структуры. А это уже представляет собой морфостатическую секрецию, свойственную высшим позвоночным.
Мысль Иордана, что первоначально секреция была связана с гибелью целой клетки, а затем лишь с гибелью ее части (которая по мере совершенствования функции становилась все меньше), подкупает своей простотой. Между тем морфонекротическая и морфокинетическая секреция — явления совершенно различного порядка. Можно представить себе переход морфо-кинетической секреции в морфонекротическую: клетки при некоторых условиях начинают отторгать при секреции вое большее количество цитоплазмы, что в конечном итоге должно привести к преобразованию морфокипетической секреции в морфонекротическую. Но как гибель неспециализированной клетки превращается в работу живой, высокоспециализированной клетки, понять трудно. Действительно, морфонекротическая секреция не предполагает высокой дифференциации структур и функций клеток, тогда как морфокинетическая секреция требует достаточно высокой и своеобразной специализации эпителиальных клеток. Но как только мы согласимся с этим, возникает вопрос: какие менее специализированные формы работы клетки предшествовали морфокинетичеокой секреции? Теория Иордана не дает ответа на этот вопрос.
Следует предположить, что морфонекротическая секреция не могла быть основой для возникновения морфокинетической, а последняя настолько специализирована, что сама имеет длинную историю. Но можно согласиться с Иорданом в том, что морфостатическая секреция возникла из функций, свойственных малодифференцированным структурам простейших животных.
Другим недостатком теории X. Иордана является ее преимущественно морфологическая сущность. В настоящее время признано, что эволюция структуры связана с изменением направления или интенсивности функционирования органа. Наконец, теория Иордана кажется устаревшей уже по одному тому, что связывает появление секреции с высокими стадиями эволюции, тогда как в последние десятилетия появилось большое число работ, доказывающих наличие истинной секреции у простейших и бактерий.
Теория Иордана могла бы считаться доказанной, если бы удалось обнаружить, что в ходе эволюции морфонекротическая секреция сменяется морфокинетической, а последняя — морфостатической. Но на всех этапах эволюции мы обнаруживаем существование всех трех основных типов секреции, что позволило высказать не одно, а несколько предположений. Например, можно считать, что морфонекротический, морфокинетический и морфостатический типы секреции являются тремя различными направлениями развития секреторной деятельности. Можно также предположить, что морфонекротическая секреция возникла из морфокинетической и т.д.
4.5.2. Экскреторная теория происхождения внешней и внутренней секреции
Более четверти века назад мною было высказано предположение относительно происхождения секреторной деятельности пищеварительных желез. Оно в некотором отношении было противопоставлением теории Иордана и казалось полезным при объяснении природы отдельных феноменов, связанных с работой пищеварительных желез. Согласно экскреторной теории, секреция возникает из неспецифической экскреции, которая существует на самых ранних стадиях эволюции живого как элемент обмена веществ между вне- и внутриклеточной средами (рис. 25). В условиях такого обмена осуществляется удаление продуктов катаболизма. Однако этот процесс не идеален даже у высокоорганизованных форм, так как вместе с продуктами распада во внешнюю по отношению к клетке среду попутно выделяется некоторое количество биологически полезных и активных молекул. Их молекулярная масса варьирует и может достигать значительной величины, например у ферментов, некоторых гормонов и витаминов. Неизбежная «потеря» полезных материалов может при соответствующих условиях стать полезным признаком. В результате естественного отбора на основе усиленного выделения определенного продукта может происходить формирование секреторного процесса.
Оказалось, что экскреторная теория способна объяснить такое парадоксальное явление, как потеря ферментов и гормонов с мочой. Она объясняет также внутреннюю секрецию пищеварительных ферментов и экзосекрецию гормонально активных веществ в составе пищеварительных соков. Формирование секреторного процесса на базе неспецифической экскреции, по-видимому, может сводиться к следующему. Полезным признаком становится выделение во внешнюю или во внутреннюю среду организма определенного типа молекул, а их синтез и экспорт становятся доминирующей функцией данного органа или типа клеток.
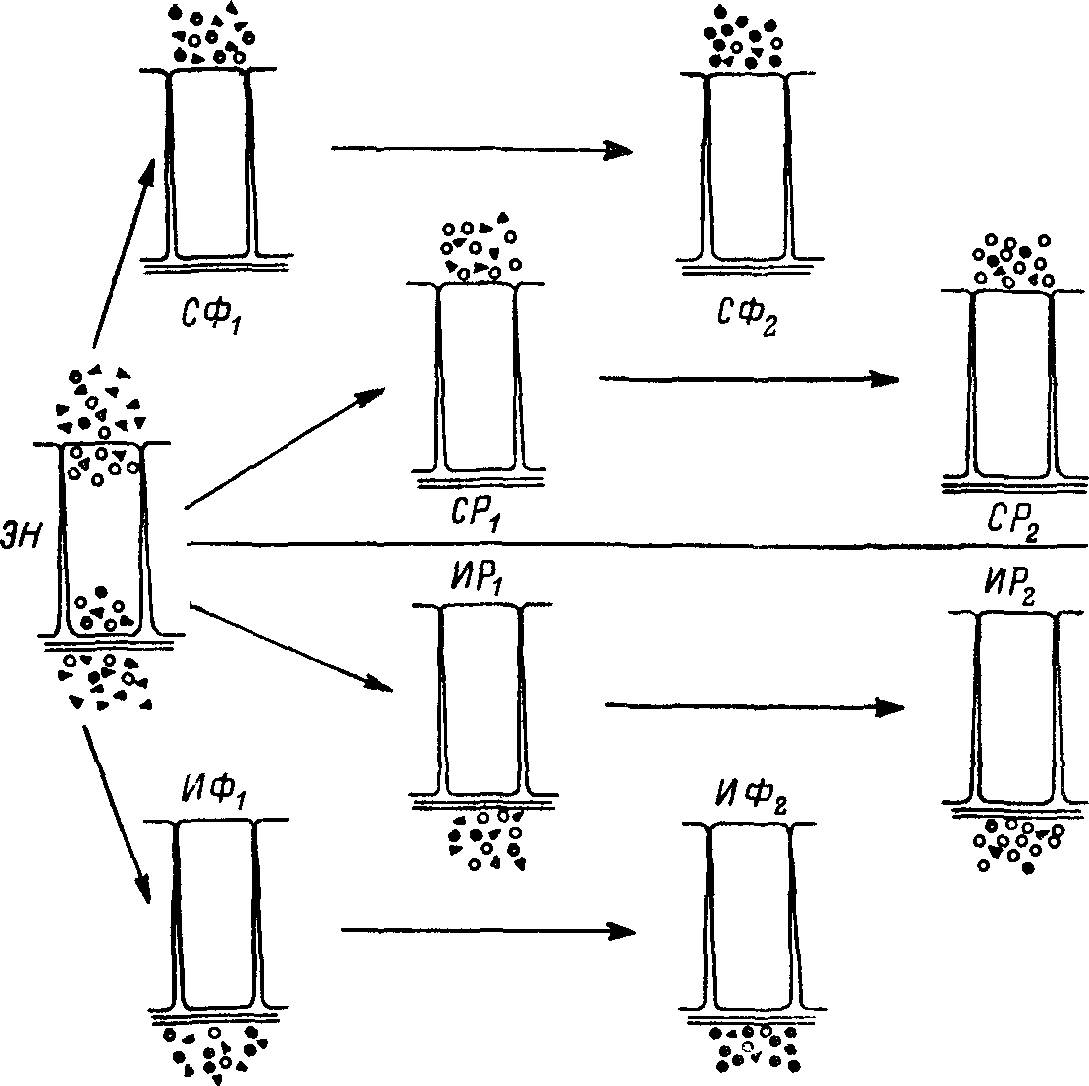
Рис. 25. Схематическая интерпретация формирования различных типов внешней и внутренней секреции в ходе эволюции на основе неспецифической экскреции.
Схема — для многоклеточных организмов, ряды СФ и СР справедливы также для одноклеточных. ЭН — неспецифическая экскреция, ряды СФ и СР — формирование внешней секреции ферментов (СФ) и регуляторов (СР); ряды ИФ и ИР — формирование соответственно внутренней секреции ферментов и регуляторов. Черные пружки — ферменты, светлые — молекулы регуляторов; треугольники — молекулы экскретируемых веществ.
На основе экскреторной теории сделан ряд предсказаний. Во-первых, истинная секреция возможна уже у простейших, в частности у амеб при их переходе от свободного к паразитическому образу жизни. Во-вторых, внешняя и внутренняя секреция могут формироваться многократно в процессе филогенеза на основе неспецифической экскреции в различных по своему происхождению структурах. Кроме того, в ходе эволюции внешняя и внутренняя секреция могут превращаться одна в другую путем сравнительно небольших транспозиций. По-видимому, экскреторная теория происхождения секреторных процессов может объяснить наличие в составе пищеварительных соков таких необычных веществ, как групповые и типовые антигены, ферменты, обеспечивающие дыхание и окислительное фосфорилирование, а также ряд других ферментов.
Вероятно, морфонекротическая секреция является особым направлением эволюции пищеварительной системы или ее некоторых отделов, тупиковой ветвью развития секреции. Морфонекротическая секреция в известных случаях оказалась настолько эффективной, что сохранилась даже у наиболее высокоорганизованных животных. Так, например, кишечная секреция позвоночных происходит в основном по морфонекротическому типу. Что касается морфокинетической секреции, то ее происхождение остается неясным. Нельзя исключить, что по крайней мере в некоторых случаях морфокинетическая секреция возникает из морфостатической.
Итак, экскреторная теория является единственной, способной объяснить с единых позиций происхождение как внешней, так и внутренней секреции и предсказать ряд феноменов, обнаруженных впоследствии. Становится ясно, что внешняя и внутренняя секреция — родственные процессы, причем некоторые органы (слюнные и поджелудочная железы, желудок, тонкая кишка) обладают как внешнесекреторной, так и внутрисекреторной функциями.
Развитие экскреторной теории приводит на первый взгляд к определенным трудностям. Так, в ходе эволюции выделяется много метаболитов, в связи с чем возникает вопрос, при каких условиях и под влиянием каких факторов некоторые из них становятся жизненно важными регуляторами типа гормонов, кейлонов и т.д. При анализе условий возникновения гормональных эффектов можно допустить экскрецию метаболитов, не имеющих мишени и приобретающих ее на более поздних стадиях эволюции, когда формируется соответствующая гормональная система. Первоначально это может быть побочным эффектом. По крайней мере один подобный случай описан М. Флоркэном в 1947 г., который сообщил, что у асцидий содержится гормон окситоцин, действующий лишь на структуры позвоноч-ных животных. Однако возможно существование мишеней, которые в течение длительного времени остаются без вещества-регулятора. Не исключено, что время, разделяющее появление этих компонентов, может исчисляться миллионами лет.
Экскреторная теория позволила преодолеть возражение М. Боданского, заключающееся в том, что одновременное возникновение в ходе эволюции гормона и соответствующего рецептора кажется невероятным. С точки зрения развиваемой теории возможно появление рецепторов без регуляторов и регуляторов без рецепторов. Такая парадоксальная ситуация подтверждена экспериментальными находками. В частности, удалось найти пример внешней секреции (а не инкреции) гормона, или биологически активного фактора. Такой феномен обнаружен нами при анализе химусных эффектов — так называемое контактное торможение пролиферации кишечных клеток химусными факторами.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Экваториальный пояс лесов и причины его происхождения
Экваториальный пояс лесов и причины его происхождения Земной шар на экваторе опоясан почти непрерывным поясом лесов от тысячи до пятисот миль шириной, который покрывает своим вечнозеленым покровом холмы, равнины и горные хребты. Только высокие пики да обрывистые склоны
ТАЙНА ПРОИСХОЖДЕНИЯ ЖИЗНИ
ТАЙНА ПРОИСХОЖДЕНИЯ ЖИЗНИ БЕЗЖИЗНЕННАЯ ЗЕМЛЯ Мы знаем, что Вселенная бесконечна. О ее протяженности можно судить по скорости света. Проходя 300000 километров в секунду, свет от самых близких звезд достигает Земли через годы. Среди многочисленных звездных систем,
§ 25. Теории происхождения хордовых
§ 25. Теории происхождения хордовых Бытует несколько точек зрения на возникновение хордовых, которые различаются как по подходам к решению проблемы, так и по животным, избранным в качестве представителя предковых групп. Наиболее известны гипотезы возникновения хордовых
Трудности теории происхождения посредством модификации.
Трудности теории происхождения посредством модификации. Уже задолго до того, как читатель дошел до этого раздела моей книги, он столкнулся с множеством трудностей. Некоторые из них настолько серьезны, что я до сих пор не в состоянии был подумать о них без некоторого
Концепции происхождения жизни
Концепции происхождения жизни С вопросом о сущности жизни неразрывно связан вопрос о ее происхождении. Существует несколько концепций происхождения жизни на Земле.Креационизм. Преподносит возникновение жизни как результат творческого акта Высшего существа в какое-то
Теория симбиотического происхождения эукариотической клетки
Теория симбиотического происхождения эукариотической клетки Кольцевая структура ДНК, структура рибосом и ряд биохимических особенностей, общие у прокариот, митохондрий и пластид, послужили доводом теории симбиотического происхождения эукариотической клетки,
ТЕОРИИ О ТЕОРИИ ЭВОЛЮЦИИ
ТЕОРИИ О ТЕОРИИ ЭВОЛЮЦИИ Немало недоразумений возникает из-за неумения отличить общеэволюционный подход от частных метаэволюционных проблем и эти последние друг от друга.На вопрос, в чем различие между теориями Ж. Б. Ламарка и Ч. Дарвина, большинство отвечает: Ламарк
Проблема происхождения трудовой деятельности
Проблема происхождения трудовой деятельности Эволюция гаптических и сенсорных функций высших млекопитающих Общеизвестно, что решающий фактор превращения животного предка — ископаемой человекообразной обезьяны — в человека был открыт около ста лет тому назад
4. Гомеостаз — поддержание внутренней среды организма
4. Гомеостаз — поддержание внутренней среды организма Мир вокруг нас постоянно изменяется. Зимние ветры заставляют нас надевать теплое платье и перчатки, а центральное отопление побуждает снимать их. Летнее солнце уменьшает потребность в сохранении тепла, по крайней
Поддержание постоянства внутренней среды
Поддержание постоянства внутренней среды В этой главе мы рассмотрели две основные системы, используемые мозгом для координации деятельности организма в соответствии с требованиями внутренней среды и внешними условиями,Вегетативная нервная система осуществляет общее
17. Гипотезы происхождения человека
17. Гипотезы происхождения человека Вспомните!Какие гипотезы происхождения человека вам известны?Проблема антропогенеза (исторического развития человека) относится к числу сложнейших философских и естественно-научных проблем. Вопрос о происхождении человека всегда
8. Система органов внутренней секреции
8. Система органов внутренней секреции Органами внутренней секреции называются железы, вырабатывающие и выделяющие непосредственно в кровь особые вещества — гормоны. Характерной особенностью гормонов является их способность в ничтожно малых количествах оказывать
Глава 1. В поисках нашей внутренней рыбы{1}
Глава 1. В поисках нашей внутренней рыбы{1} С тех пор как я стал взрослым, мое лето обычно проходит среди снега и слякоти далеко к северу от полярного круга за раскалыванием камней. Большую часть времени я мерзну, натираю мозоли и не нахожу ровным счетом ничего. Но если