6.3. Функциональные блоки и механизмы эволюции
6.3. Функциональные блоки и механизмы эволюции
Концепция универсальных функциональных блоков дает возможность проанализировать некоторые стороны эволюционного процесса, так как функциональный блок является тем элементом, из которого построена вся система функций любой сложности, подобно тому как ген или кодон служат элементам, из которых построена информационная система организма. Попытаемся показать некоторые особенности эволюции на основе универсальных функциональных блоков.
6.3.1. Примеры эволюции функций на основе общих регуляторных блоков
Работа с ограниченным числом блоков обеспечивает более устойчивую регуляцию и более быстрые перестройки. Поиск нужных деталей и сборка машин из стандартных деталей являются, по-видимому, более простым актом при реализации естественных технологий. В пределах не только одного типа, но часто и одного класса организмов возможны резкие перестройки, что было проиллюстрировано на примере пищеварения. В пределах лишь одного типа плоских червей можно найти примеры эктодермального пищеварения и его исчезновения. Это означает заметную ограниченность закона Долло о необратимости эволюции на уровне функциональных блоков.
Еще одна закономерность проявляется в перемещении регуляторных блоков, например рецепторов и взаимодействующих с ними лигандов. Недооценка этого феномена служит причиной многочисленных ошибок, которые первоначально воспринимались как крупнейшие достижения научной мысли и несомненно были таковыми (см. гл. 7). Например, кальмодулин обнаружен в клетках множества видов как позвоночных, так и беспозвоночных организмов, а также высших и низших растений (табл. 16). Далее, система циклических нуклеотидов рассматривалась как новый эволюционный инструмент, возникший у сложных организмов и выполняющий функцию вторичных посредников. Однако, как отмечено выше, система циклических нуклеотидов развита уже у простейших, у которых нет ни внутренней среды, ни интегрированных частей сложного многоклеточного организма. У простейших цАМФ выполняет функции не вторичного, а первичного мессенжера. У простейших обнаружены и многие гормоны, которые действуют параллельно, а затем образуют последовательную цепь сигналов и ретрансляторов. Это справедливо также для основных нейротрансмиттеров. Так, ацетилхолин, норадреналин и вновь открытые нейротрансмиттеры, такие, как аминокислоты, АТФ и др., используются как совершенной, так и примитивной нервной системой, включая нервную систему плоских червей и кишечнополостных. Даже такие медиаторы, как ацетилхолин и катехол-амины, давно признаны универсальными и показаны у всех животных, имеющих нервную систему.
Различные организмы, у которых изолирован кальмодулин
Животные Простейшие Членистоногие Амеба Рак Эвглена Рыбы Парамеция Электрический скат Тетрахимена Амфибии Кишечнополостные Шпорцевая лягушка Морской анемон Птицы Морские анютины глазки Цыпленок Нематоды Млекопитающие Кольчатые черви Человек Земляной червь Корова Моллюски Овца Осьминог Свинья Гребешок Кролик Иглокожие Крыса Морской еж Хомяк Морская звезда Мышь Растения Покрытосемянные Грибы Ячмень Слизевики Хлопок Физарум Орех Зеленые водоросли Шпинат XламидомонадаСовременный уровень знаний позволяет думать, что на всех этапах эволюции в качестве медиаторов могут выступать молекулы различных типов, в том числе аминокислоты, их производные — адреналин и норадреналин, пептиды, а также некоторые нуклеотиды, эфиры и т.д. К медиаторам могут быть отнесены углеводы, медиирующие феномен узнавания в различных клетках организмов разных видов, начиная с вирусов и бактерий и кончая млекопитающими. Однако роль медиаторов в ходе эволюции меняется, хотя законы, по которым значимость разных нейротрансмиттеров увеличивается или уменьшается, остаются неизвестными. Даже если ограничиться нервно-мышечной передачей, то трудно ответить на вопрос, почему у некоторых беспозвоночных (в частности, ракообразных и насекомых) столь важную роль в качестве нейротрансмиттеров играют аминокислоты, а у млекопитающих — ацетилхолин? Точно так же широкое распространение имеют пептидные гормоны, первоначально обнаруженные у высших организмов, а затем у многих низших позвоночных и беспозвоночных. Интересно, что у позвоночных, насекомых и ракообразных, несмотря на отсутствие истинной гомологии, наблюдается поразительное сходство не только функций, но и организации эндокринной системы. При этом химическая структура гормонов у этих организмов обладает значительным сходством.
Важно, что процессы осморегуляции у костистых рыб находятся под контролем гормонов, которые известны у других животных, в том числе у млекопитающих. Однако у рыб они выполняют другую функцию. В частности, кортизол, как и NaCl, вызывает дифференциацию и пролиферацию хлоридных клеток, пролактин — их дифференциацию, глюкагон и ВИП оказывают преимущественно стимулирующий эффект. Таким образом, осморегуляция контролируется гормонами, которые у высших организмов мало связаны с водносолевым обменом.
Для организмов различных групп, естественно, общими являются не только химические сигналы, но и воспринимающие их рецепторы и, наконец, системы, связанные с переработкой и ретрансляцией сигналов. Например, универсальна система циклических нуклеотидов. Такие гормоны, как адреналин, глюкагон, парат-гормон, антидиуретический и тиреотропный гормоны, кортикотропин, лютеинизирующий гормон, гипотала-мические рилизинг-факторы и меланоцитстимулирующий гормон стимулируют в клетках-мишенях мембранносвязанную аденилатциклазу. В результате этого количество цАМФ внутри клетки удваивается. Образующийся в клетках цАМФ вызывает гормональный эффект, действуя в качестве вторичного мессенжера. Этот механизм у позвоночных и беспозвоночных организмов весьма сходен. Так, при действии серотонина на мышечную ткань печеночной двуустки посредником служит цАМФ. Существуют также сведения об участии цАМФ в гормональных эффектах у насекомых, а при действии хроматофоротропинов — у ракообразных. У большинства клеток эукариотов токсин холеры активирует аденилатциклазу.
6.3.2. Перемещение функциональных блоков в филогенезе
Перемещение гормонов и рецепторов хорошо документировано. Хотя каждый функциональный блок может выполнять одну или две-три высокоспециализированные операции, при определенных сочетаниях они могут образовывать различные молекулярные машины более высокого порядка. Так, осморегуляторная система, поддерживающая постоянство солевого состава внутренней среды организмов, и секреторные органы, участвующие в процессах пищеварения, состоят из одних и тех же функциональных блоков. Нередко одни и те же органы могут, несколько меняя количественные соотношения различных функциональных блоков, превращаться из секреторных в экскреторные и наоборот. Эволюционный переход секреторных клеток во всасывающие и наоборот, охарактеризованный как следствие временной и пространственной транспозиции определенных каналов и насосов, четко показан (рис. 36 и 37). Ярким примером может служить пищеварительная система насекомых, органы которой переходят от секреторной функции к всасывающей, от всасывающей к экскреторной и т.д.
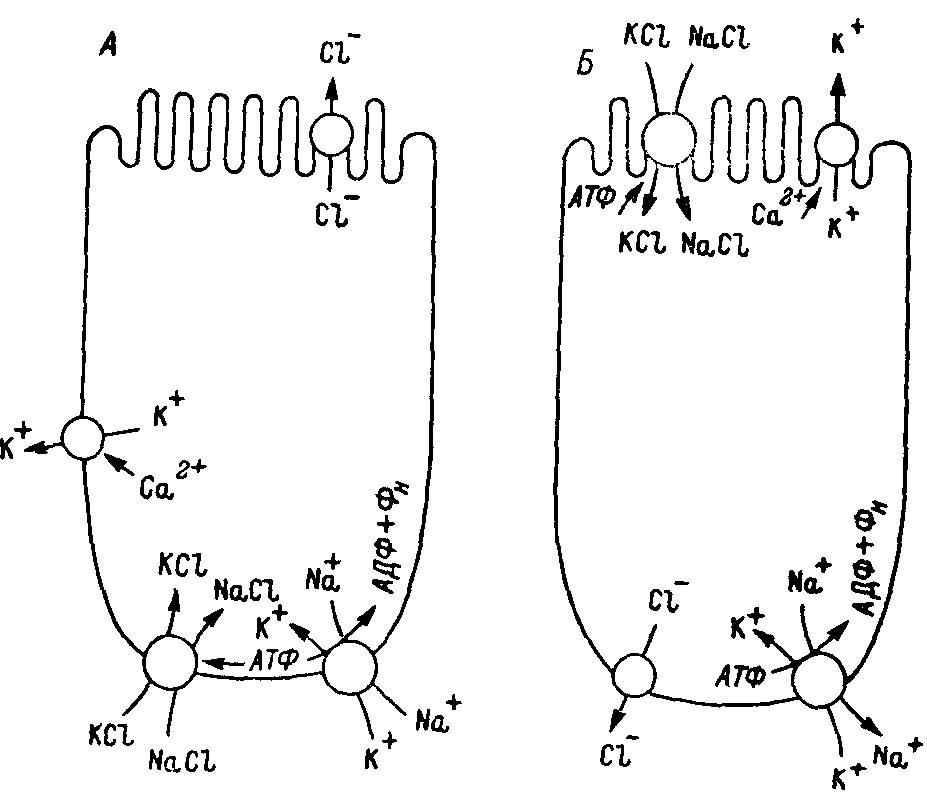
Рис. 36. Предполагаемая локализация систем, транспортирующих одновалентные ионы, в эпителиальных клетках, секретирующих (А) и всасывающих (Б) хлор.
В обеих системах Са2+ может активировать Са2+-зависимые каналы для транспорта К+. Эти каналы вместе с потоком Сl- через возможные каналы для транспорта С1- откачивают КСl из цитоплазмы, снижая электрохимический градиент, вызванный выходом Na+ и поступлением К+ за счет Na+, К+-АТФазы. Симпорт NaCl/KCl может быть активирован цАМФ зависимой или цАМФ-независимой протеиназой в различных тканях.
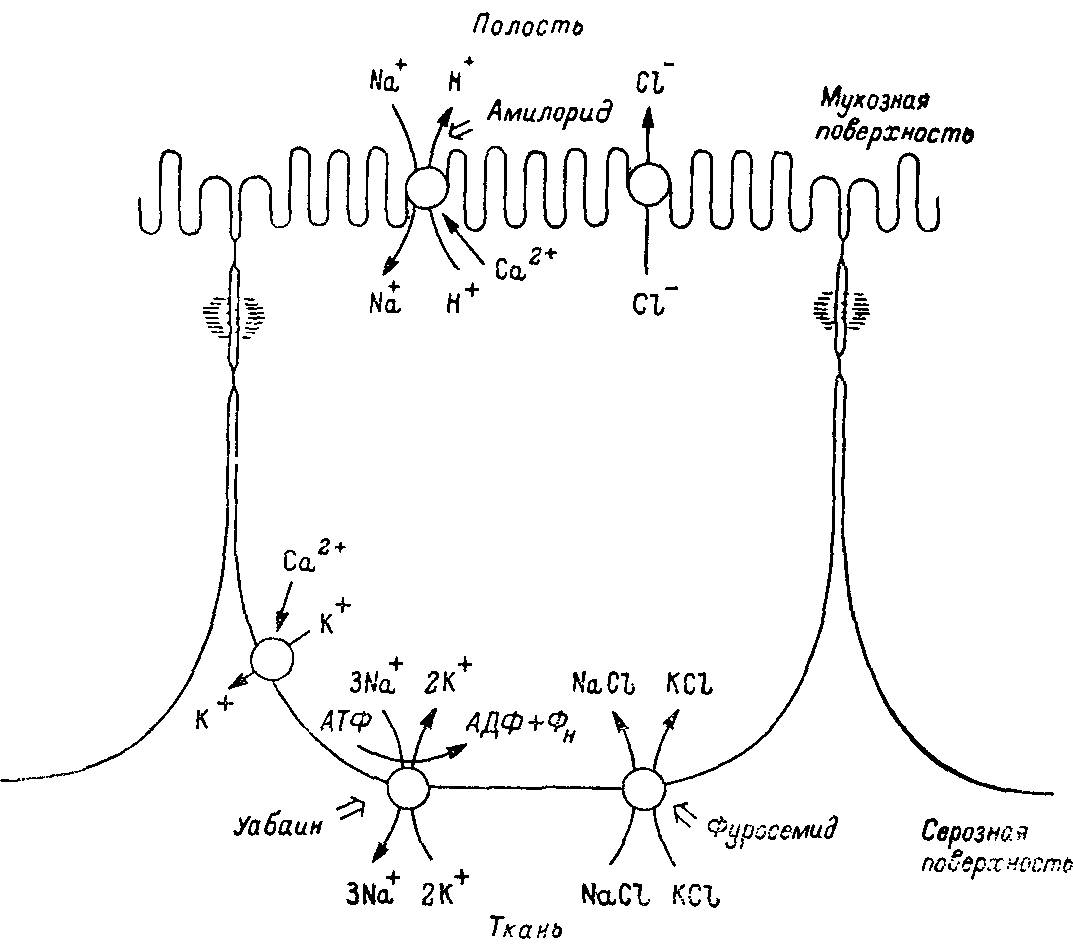
Рис. 37. Схема асимметричной локализации системы, транспортирующей одновалентные ионы, в эпителиальных клетках почки.
Локализация Са2+-актнвируемых каналов для К+ не ясна. Механизм транспорта Сl- через мембрану эпителиальных клеток не определялся.
Остановимся на процессах экскреции у млекопитающих и насекомых. Исходя из того, что эти организмы представляют собой высшие и, следовательно, наиболее разошедшиеся точки дивергентной эволюции первично- и вторичноротых, можно допустить следующее. Значительные различия в строении экскреторной системы насекомых и млекопитающих основаны на использовании различных молекулярных механизмов. Однако системы насосов и каналов, реализующие транспорт воды и ионов, практически идентичны в почке млекопитающих, в выделительной системе насекомых, где терминальная кишка превратилась в экскреторный орган, а также в кишке млекопитающих, выполняющей пищеварительные функции. Во всех случаях обнаружена уабаинчувствительная Na+,K+-АТФаза, которая служит главным первично энергизованным насосом. Идентифицированы также системы натриевых и хлорных каналов, чувствительных соответственно к эпокриновой кислоте и амилориду (рис. 38).
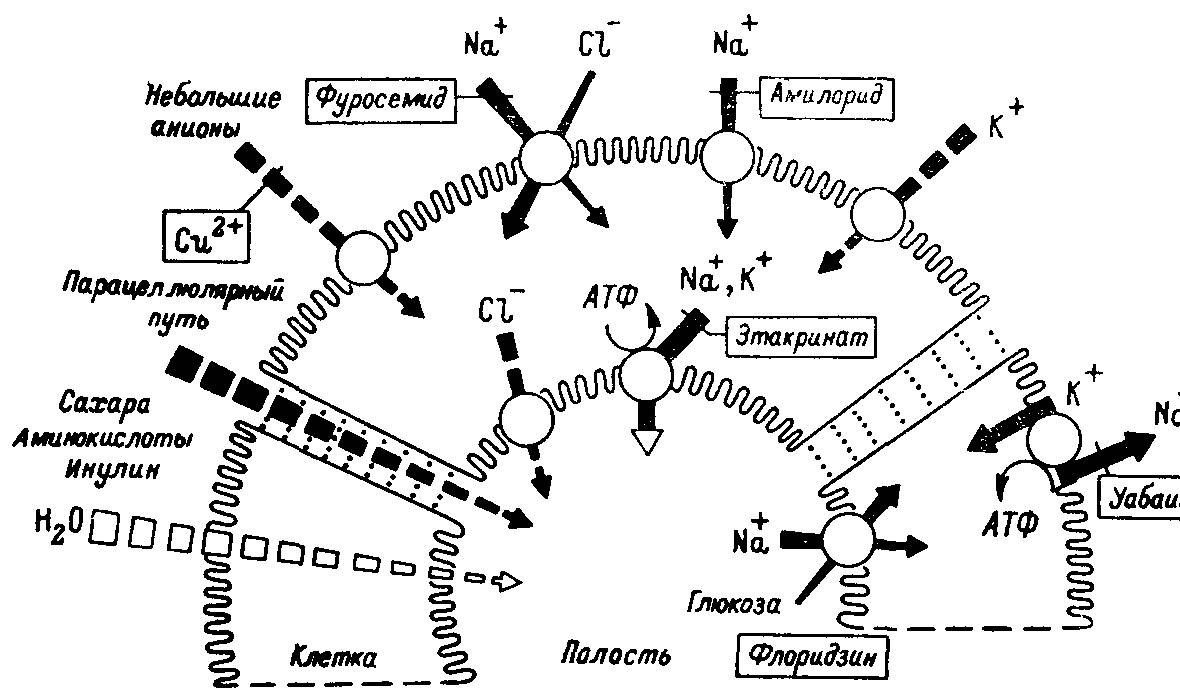
Рис. 38. Схема поперечного среза мальпигиева сосуда насекомых, иллюстрирующая мембранный механизм, связанный с секрецией жидкости.
До сих пор не удалось обнаружить ни одной транспортной системы у насекомых, которая отсутствовала бы у млекопитающих. Так, механизм транспорта аминокислот у насекомых и многих других животных обладает многими общими свойствами. Однако распределение транспортных систем и поэтому многие характеристики процессов экскреции существенно различны. Формирование мочи у насекомых и млекопитающих, как и у большинства многоклеточных организмов, состоит из двух этапов: 1) образование первичной мочи, которая у позвоночных напоминает безбелковую плазму крови; 2) последующие процессинги, которые приводят к образованию вторичной мочи, содержащей в повышенной концентрации многие конечные продукты метаболизма и ряд других веществ, избыток которых удаляется из организма для поддержания ионного гомеостаза. В то же время первичная моча у насекомых образуется в результате секреции, а не фильтрации, как это имеет место у млекопитающих и многих других организмов.
Насколько можно судить, экскреторный аппарат насекомых может обеспечить удаление избытка воды и солей и строгую экономию воды. Это достигается вариациями в распределении насосов и каналов и интенсификацией поглощения солей и воды (рис. 39).
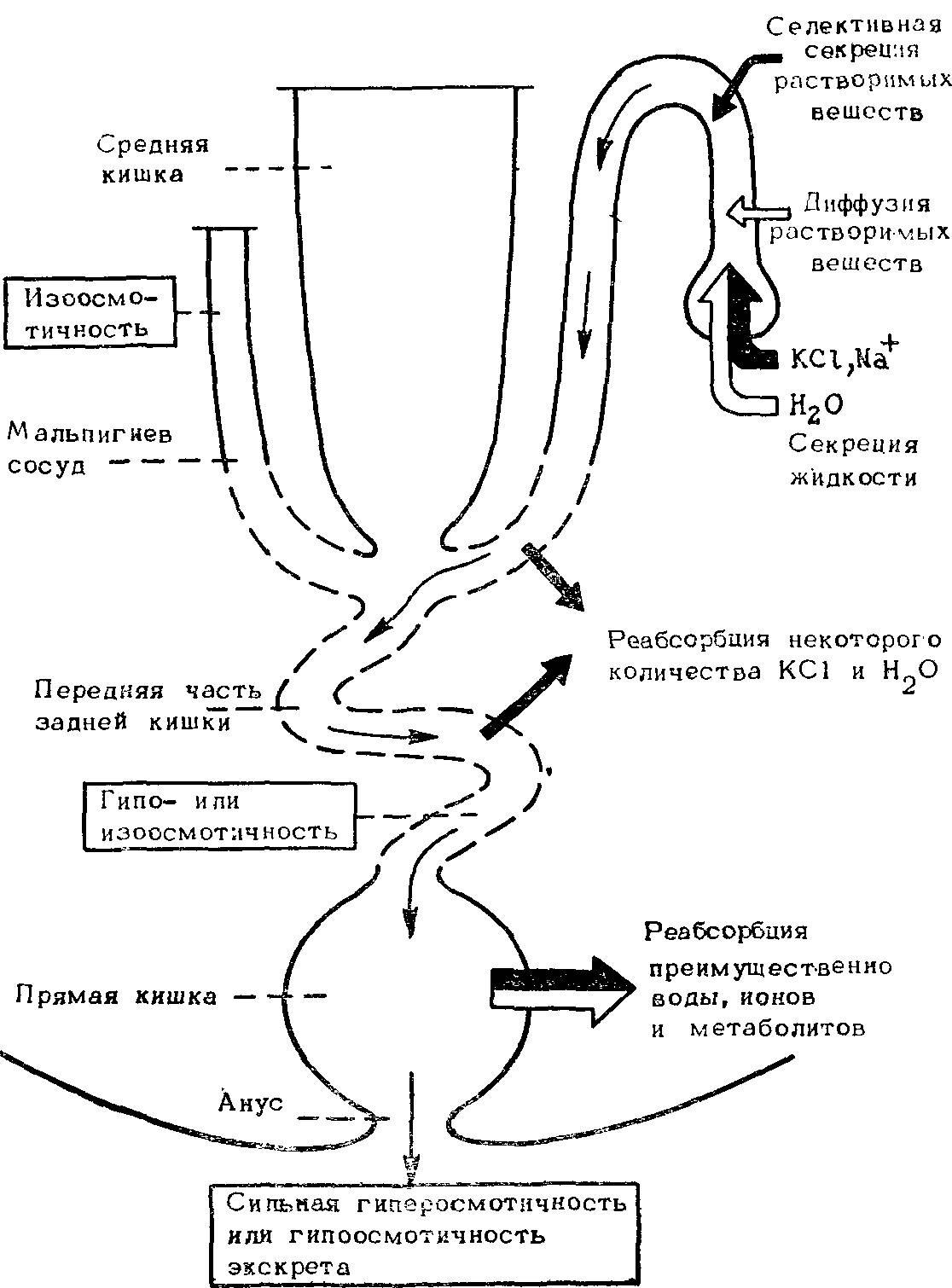
Рис. 39. Схема типичной экскреторной системы насекомых.
Указаны основные транспортные процессы и конечная осмолярность жидкости, покидающей каждый сегмент. Тонкие стрелки — выделение мочи, толстые — активный транспорт, светлые — пассивный транспорт.
Таким образом, адаптация выделительной системы насекомых и млекопитающих основана на использовании идентичных или близких функциональных блоков. Последние, однако, принадлежат к структурам, происходящим из различных зародышевых листков и, казалось бы, с различной эволюционной специализацией. Еще более замечательным является то, что многие регуляторные функции млекопитающих и насекомых осуществляются с помощью идентичных молекул.
Одним из примеров перемещения гормонально активных веществ и изменений контура эндокринной регуляции может служить формирование двух источников АКТГ — нейроэндокринного (гипоталамо-гипофизарная система) и желудочно-кишечного. Ясно, что в ходе эволюции подобные взаимоотношения могли складываться лишь как результат перераспределения функциональных блоков.
Интересно, что одни и те же пептиды могут синтезироваться в тканях, имеющих различное фило- и онтогенетическое происхождение. Так, холецистокинин и гастрин могут синтезироваться в эпителиальных клетках экто- и энтодермального происхождения и в нервных клетках.
АТФ — основной макроэрг в механизме первичной энергизации, в ходе эволюции может выступать также в роли нейротрансмиттера. При этом АТФ включен в синаптические везикулы, и его эффект, как и других нейротрансмиттеров, реализуется с помощью экзоцитоза. Не менее выразительным примером является использование для передачи сигналов синапсов аминокислот, которые накапливаются в гранулах и действуют при экзоцитозе. В настоящее время известны глициновая и глутаминовая сигнализации, а также рецепторы для связывания аминокислот.
Перестановка функциональных блоков в пределах клетки, их перераспределение между различными частями одного органа или различных органов не связаны с какими-либо ограничениями. Действительно, увеличение или уменьшение количества рецепторов инсулина, изменение типов секреции и количества выделяемого гормона не нарушают взаимоотношений рецептора и лиганда, но создают ряд новых ситуаций в таких взаимодействиях. Некоторые изменения могут оказаться полезными. Так, инсулинпродуцирующие клетки, первоначально локализованные в тонкой кишке (например, у круглоротых), затем переместились в поджелудочную железу. Глюкагонпродуцирующие клетки, также первоначально локализованные в тонкой кишке у высших позвоночных, частично сохранились в этой области. В то же время другая часть таких клеток транслоцировалась в поджелудочную железу.
Н. Трак в 1973 г. высказал гипотезу о существовании универсального кишечного прогормона. Автор предположил, что кишечные гормоны произошли в результате мутации из проинсулинподобной молекулы-предшественника в ходе эволюционного развития. При дупликации гена из проинсулинподобной молекулы возникли две молекулы проинсулина, одна из которых стала предшественником инсулина, а другая (при дальнейшей мутации и дупликации гена) — гастрина и секретина. Секретинподобная молекула стала, в свою очередь, предшественником секретина и глюкагона.
Еще более ярким примером того, как концепция функциональных блоков дает ключ к пониманию ранее таинственных явлений, может быть происхождение различных ядов. Этот пример демонстрирует перемещение функциональных блоков в филогенезе.
6.3.3. Происхождение ядов. Гормоны и яды
Еще Ч. Дарвин обращал внимание на трудности в объяснении происхождения ядов. Недавно мною высказано предположение, что объяснение происхождению ядов следует искать в концепции универсальных функциональных блоков. Допускается, что яды появляются в результате постепенного формирования новых физиологически активных факторов ради защиты, нападения и добывания пищи и образуются в специализированных органах благодаря транслокации гена или его экспрессии из одного оперона в другой. При этом первоначальная физиологическая роль данного фактора могла заключаться в поддержании регуляторных функций организма. Тот же фактор, продуцируемый в больших количествах и вводимый при защите и нападении, вызывает токсикоз у жертвы. Например, фосфолипазы — физиологические ферменты в составе мембран, при введении в кровь с ядом змей провоцируют ее гемолиз и другие деградационные процессы, приводящие к смерти. Точно так же функцию физиологических регуляторов в своем организме могут выполнять батрахотоксин и другие вещества. В то же время накопленные в слюнных железах и введенные в организм человека, они вызывают токсический эффект.
При анализе молекулярной структуры показано, что яды, как и гормоны, представляют собой пептиды и в ряде случаев имеют сходную структуру. В коже амфибий обнаружено много активных пептидов с широким спектром действия. Кожные яды амфибий присутствуют в качестве гормонов в желудочно-кишечном тракте млекопитающих, а также в качестве рилизинг-факторов в их организме. Примером этому может служить бомбезин — один из наиболее известных ядов кожи лягушки. Этот пептид у млекопитающих, в том числе у человека, по отношению к гормонам желудочно-кишечного тракта служит рилизинг-фактором, т.е. гормоном, обеспечивающим освобождение из клеток других гормонов. Точно так же бомбезин и некоторые другие гормоны и кожные яды присутствуют в центральной нервной системе высших организмов.
Таким образом, яды кожи лягушки — вещества, сходные с уже известными гормонами, а во многих случаях идентичные им. Подобное заключение сделано в отношении других ядов. В частности, недавно обнаружены два пептида. Один из них — саувагин, изолированный из кожи лягушки, с физиологическими эффектами на сердечно-сосудистую систему и переднюю долю гипофиза теплокровных, другой — дерморфин — вещество с мощными опиатными эффектами, в том числе аналгезийными.
Важно, что все обнаруженные кожные пептиды амфибий имеют функциональные аналоги с гормонами энтеронейрональной оси млекопитающих. Так, функциональные эффекты церулеица сходны с таковыми холецистокинина и гастрина, эффекты тахикинина — с субстанцией Р и т.д. В кишке и мозгу выявлена бомбезинподобная иммунореактивность (табл. 17). Еще более поразительно, что структура пептидов кожи амфибий близка, а в некоторых фрагментах идентична структуре гормонов человека и высших животных. Подобные совпадения не случайны и отражают единство происхождения пептидов. Это означает, что гипотеза происхождения ядов в результате генетических экспрессий сформировавшегося физиологически активного вещества в других органах получает подтверждения.
Семейство пептидов Кожи лягушки в аналогов пептидов в кишке в мозгу млекопитающих
Кожа Кишка Мозг Тахикинины Субстанция Р Субстанция Р (эледозин, физалемин) Физалеминподобные пептиды Физалеминподобные пептиды Церулеин Xолецистокинин/гастрины Xолецистокинин-8 Брадикинины ? Брадикининподоб-ная иммунореактивность Боыбезины Бомбезинподобные пептиды Бомбезинподобные пептиды Ксенопсин Нейротензин Нейротензпн Ангиотензин Ангиотензины Ангиотензины Гормон, освобождающий тиреотропин Гормон, освобождающий тиреотропин Гормон, освобождающий тиреотропин ВИП ВИП ВИП Саувагин ? ? Дерморфин Опиоидные пептиды Опиоидные пептидыИтак, в состав ядов змей, насекомых, амфибий и многих других организмов входят известные ферменты и физиологически активные вещества, выполняющие регуляторные или функциональные нагрузки. В этом случае одни и те же сигналы, переходя из одной системы в другую, участвуют в реализации не только
разных, но подчас совершенно экзотических функций. Это же справедливо для перемещения одних и тех же гормонов из желудочно-кишечного тракта в мозг, железы внутренней секреции и т.д. При новой локализации гормон может выполнять роль нейротрансмиттера, а нейротрансмиттер — гормона.
6.3.4. Перераспределение функциональных блоков в пределах клетки
Функционально неполяризованные и неспециализированные клетки характеризуются равномерным распределением каналов и насосов, благодаря чему создается наиболее равномерный и экономичный поток веществ из клетки в клетку. Однако в специализированных клетках, реализующих всасывание, наблюдается поляризация и распределение каналов и насосов так, что первые обращены в полость, из которой происходит всасывание, а вторые взаимодействуют с внутренней средой. По мнению многих исследователей, этого достаточно для обеспечения однонаправленного потока воды, натрия, нутриентов и т.д. В секреторных клетках насосы и каналы сосредоточены в области базолатеральной мембраны, которая осуществляет нутритивные функции.
Можно представить, что разные типы функциональной сцециализации в ходе эволюции или индивидуального развития связаны с транспозицией каналов в пределах клетки. Существуют данные о возможности перемещения блоков, характерных для одного типа внутриклеточных органелл, в другие органеллы. Например, Са2+, Мg2+-АТФаза — типичный насос эндоплазматической сети — может быть обнаружена в клеточной мембране. При сохранении функциональной топографии клетки, например кишечной, достаточно включения механизмов секреции Сl- на апикальной мембране, чтобы превратить механизм, обеспечивающий всасывание воды и солей, в механизм их выделения. Изменение направления потоков воды и солей, которое, в частности, наблюдается при диарее различной этиологии, имеет место также в органах, реализующих удаление избытка солей, а именно в солевых железах и жабрах.
6.3.5. Транспозиция функциональных блоков между клетками и органами
Гормоны, продуцируемые клетками желудочно-кишечного тракта, могут также выделяться клетками центральной нервной системы, где они выполняют функции нейромедиаторов и нейромодуляторов. У амфибий некоторые гормоны выделяются клетками кожи и играют роль ядов. Это служит одним из примеров того, как область, где синтезируются вещества, может сужаться и расширяться в ходе эволюции. Рядом исследователей, в том числе в нашей лаборатории, показано, что активный транспорт глюкозы у новорожденных крысят доминирует в дистальном отделе тонкой кишки, а у взрослых животных в этом же отделе он полностью отсутствует и наиболее выражен в проксимальном. Имеются также существенные видовые различия в распределении ферментативных и транспортных активностей вдоль тонкой кишки.
Распределение функциональных блоков между клетками и органами или изменение их соотношения — важный путь эволюции биологических систем, хотя свойства самих блоков могут меняться мало или совсем не меняться.
6.3.6. Рекомбинация функциональных блоков
Один из путей эволюции — объединение функциональных блоков в уникальные по своим свойствам ансамбли, примером которых могут служить транспортные АТФазы. Однако до сих пор неясно, какие функции выполняли блоки, ставшие обязательными компонентами этих сложных молекулярных машин. Другой впечатляющий пример роли сочетаний функциональных блоков представляет собой система, осуществляющая цикл Кребса. Компоненты этой системы возникли в анаэробный период, хотя вся система в целом является наиболее эффективным инструментом использования кислорода. Наконец, как упоминалось, недавно нами получены факты в пользу того, что Nа+-зависимый транспорт глюкозы реализуется с помощью устройства, в котором сочетаются два транспортера — натриевый, выполняющий функцию вторичного энергизатора, и глюкозный, выполняющий трансмембранный перенос глюкозы, а также функцию акцептора энергии. Предполагается, что натриевый энергизатор служит общим блоком для всех видов трансмембранного Na+зависимого переноса, появление которого обеспечило переход от облегченной диффузии на транспортерах к активному транспорту. В пользу такого предположения свидетельствует также то обстоятельство, что Nа+-зависимый транспорт органических веществ в том виде, в каком он существует у высших организмов, имеется лишь у эукариотов. По всей вероятности, формирование новых надмолекулярных комплексов в ходе эволюции представляет собой более редкое событие, чем внутри-или межклеточное перемещение.
Фундаментальная роль рекомбинаций не означает отрицания эволюции функциональных блоков. Наиболее разумным представляется допущение, что в ходе эволюций важны оба механизма эволюции функций:
1) рекомбинация стабильных функциональных блоков;
2) эволюция этих блоков. Введение представлений о коррелятивных ограничениях изменчивости функциональных блоков позволяет понять, в каких случаях действует первый, а в каких — второй механизмы. Если данный признак связан с многими взаимодействиями, его изменения вероятнее всего приводят к нарушению гармоничной системы. Тогда отбор действует как стабилизирующий- фактор и компоненты комплекса в ходе эволюции меняются мало.
При довольно широко распространенном процессе дупликации гена, кодирующего какой-либо функциональный блок, он может выходить из-под контроля стабилизирующего отбора. Так, блок, выведенный за пределы системы, может оказаться ценным по производимым им эффектам (например, по гидролизу определенного типа связей) независимо от того, включен он во взаимодействие с другими компонентами комплекса или нет. Другим примером этой закономерности может служить эволюция различных кислых (карбоксильных) протеаз, которые, мало меняясь в ходе эволюции, участвуют в качестве блоков в реализации различных функций (рис. 40).
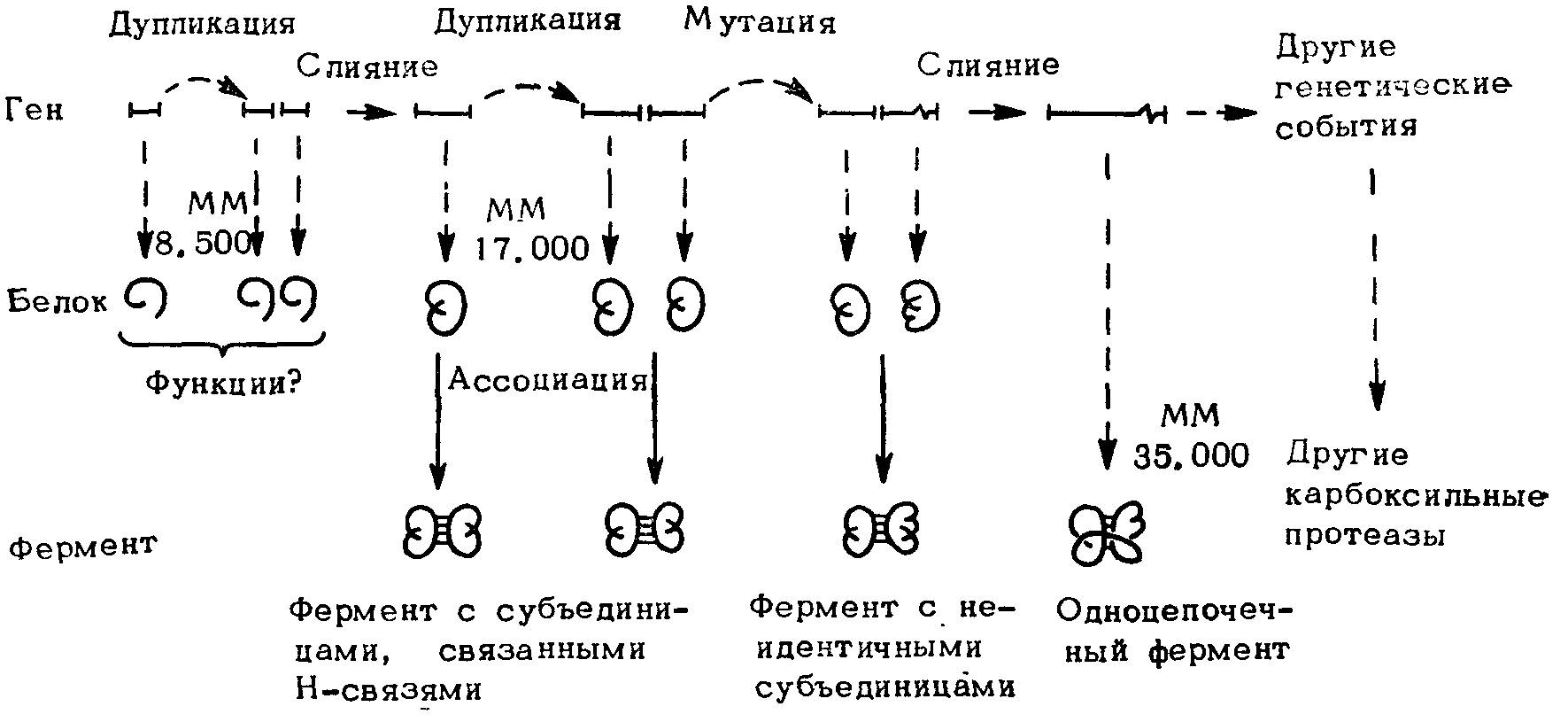
Рис. 40. Схема генетических событий в эволюции карбоксильных протеаз.
Размер анцестрального гена, по-видимому, составляет 1/4 настоящего. После дупликации первого гена и слияния, примитивная карбоксильная протеаза, вероятно, содержала две субъединицы. В результате дупликации второго гена и слияния образовался одноцепочечный фермент. Эти гипотетические события обобщены на основе структурной информации, которая показала, что карбоксильные протеазы содержат сходные структурные единицы, организованные в две отчетливо гомологичные доли.
Вероятно, после дупликации гена, контролирующего синтез трипсина (точнее, трипсиногена), мог происходить ряд мутаций. В результате этого свойства образующегося нового продукта в отличие от трипсиногена могли не зависеть от активации энтерокиназой или от способности трипсина активировать химотрипсиноген, прокарбоксипептидазы и т.д. Судя по всему, благодаря подобным дупликациям гена и следующим за ними мутациям могло возникнуть большое количество сериновых протеаз, выполняющих самые разнообразные функции.
Таким образом, в ходе эволюции сочетаются рекомбинации стабилизированных функциональных блоков и эволюция, подчас быстрая, тех блоков, на которые уменьшается стабилизирующее действие естественного отбора. По всей вероятности, дело не сводится к простой дивергенции исходного и дуплицированного гена. Дивергенция является скорее кажущейся, так как основной ген, включенный в определенную систему, стабилизирован, а меняется чаще всего ген, вырванный из системы.
К сожалению, генетические и физиологические аспекты эволюционного процесса характеризуются неоднозначностью. Имея в виду эти обстоятельства, попытаемся обобщить физиологические аспекты проблемы, опираясь на интерпретацию генетических закономерностей. По-видимому, физиологи должны базироваться на наиболее надежных законах генетики и эволюционной теории. К числу таких законов или закономерностей относятся следующие:
1) эволюция реализуется в результате случайных точечных мутаций, которые осуществляются с определенной частотой;
2) предполагается, что мутации приводят к изменению функциональных характеристик в генных экспрессиях или в собственных мутирующих структурах;
3) нейтральные мутации служат молекулярногенетической основой формирования новых побочных эффектов, которые могут стать материалом последующей селекции.
Частота замещения аминокислот может быть способом измерения времени возникновения двух родственных белков из общего предшественника. Частота замещения аминокислот, с одной стороны, определяется вероятностью мутаций, а с другой — закреплением возникающего замещения в ходе эволюции.
Ряд авторов отметили ограничения в мутационной изменчивости некоторых белков. Так, физиологически значимые участки одних и тех же белков, например инсулина и проинсулина, могут обладать различной эволюционной стабильностью, отличающейся во много раз (табл. 18). Схематизированная структура проинсулина и инсулина представлена на рис. 41.
Средние скорости замещения аминокислот на участок за год у позвоночных
Кожа Кишка Мозг Тахикинины Субстанция Р Субстанция Р (эледозин, физалемин) Физалеминподобные пептиды Физалеминподобные пептиды Церулеин Xолецистокинин/гастрины Xолецистокинин-8 Брадикинины ? Брадикининподоб-ная иммунореактивность Боыбезины Бомбезинподобные пептиды Бомбезинподобные пептиды Ксенопсин Нейротензин Нейротензпн Ангиотензин Ангиотензины Ангиотензины Гормон, освобождающий тиреотропин Гормон, освобождающий тиреотропин Гормон, освобождающий тиреотропин ВИП ВИП ВИП Саувагин ? ? Дерморфин Опиоидные пептиды Опиоидные пептиды
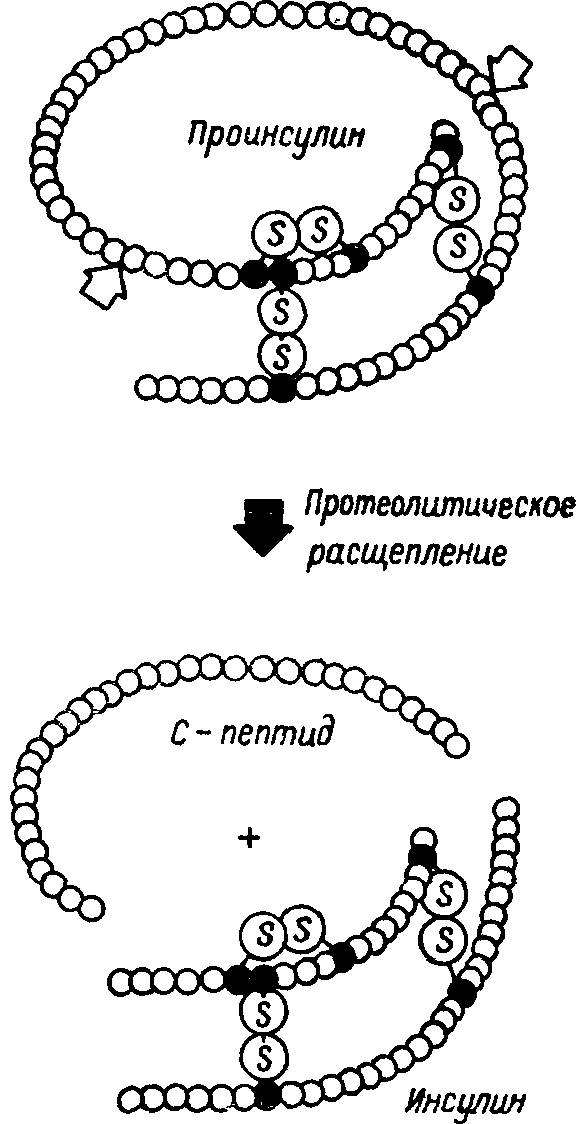
Рис. 41. Схема протеолитического расщепления молекулы проинсулина с образованием С-концевого пептида и инсулина.
Необходимо еще раз подчеркнуть, что функциональные блоки выполняют не биологические, а элементарные функции, которые могут иметь различное значение в зависимости от того, в какой функциональный комплекс они включены. Поэтому эволюция функциональных блоков лимитирована системой взаимодействий, причем такая система и функциональные операции хорошо подогнаны друг к другу. Это объясняет стабильность функциональных блоков, включенных в определенные системы, и быструю эволюцию сходных блоков, которые в результате дупликации генов или по каким-либо другим причинам выводятся за пределы системы взаимодействующих блоков.
***
Приведенные примеры на первый взгляд кажутся разрозненными. Между тем они демонстрируют, что: 1) мир живых существ состоит не только из одних и тех же строительных блоков, но и из близких, часто идентичных функциональных блоков; 2) общие функциональные блоки участвуют как в микро-, так и в макроэволюции; 3) на основе общих функциональных блоков возможны эволюционные и филогенетические революции, когда экто- или энтодерма могут стать органом дыхания и органом пищеварения, а желудочно-кишечный тракт реализовать не только пищеварительную, но и эндокринную, выделительную и другие функции; 4) общие функциональные блоки в ходе эволюции способны к перемещению; 5) на основе общих функциональных блоков могут происходить взаимодействия между различными системами одного организма, между организмами одного вида и между организмами, относящимися к разным видам и даже типам.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Блоки поведения
Блоки поведения Все сказанное выше было лишь нисколько растянутым введением, задача которого состояла в том, чтобы убедить читателей: в основе биологического разнообразия часто лежит комбинирование несложных блоков и модулей. Быть может, и сложное поведение животных,
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции Пер. А. НадирянВ этой и следующей главах дается краткое описание современного состояния эволюционной биологии, какой она была до 1995 года, когда возникло новое направление науки — сравнительная геномика.
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции Пер. А. НестеровойВ этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись
Глава 9 Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции
Глава 9 Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции Пер. Д. ТулиноваДрама ламаркизмаКак уже отмечалось в предисловии к данной книге, одной из ключевых заслуг
Глава 8 ФУНКЦИОНАЛЬНЫЕ СОСТОЯНИЯ
Глава 8 ФУНКЦИОНАЛЬНЫЕ СОСТОЯНИЯ 1. ОПРЕДЕЛЕНИЕ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ Наиболее часто функциональное состояние (ФС) определяют как фоновую активность нервных центров, при которой и реализуется та или иная конкретная деятельность человека. В классической физиологии
6. ПСИХОФИЗИОЛОГИЧЕСКИЕ ФУНКЦИОНАЛЬНЫЕ СОСТОЯНИЯ (ПФС)
6. ПСИХОФИЗИОЛОГИЧЕСКИЕ ФУНКЦИОНАЛЬНЫЕ СОСТОЯНИЯ (ПФС) Актуальность изучения ПФС определяется их вкладом в обеспечение эффективности деятельности и надёжности человека, а также увеличением количества профессий и изменением условий труда, когда психофизиологические
Функциональные нервные болезни — неврозы
Функциональные нервные болезни — неврозы К неврозам относятся: эпилепсия, эклампсия и хорея.Эпилепсия (epilеpsia). Эпилепсией называют хроническое нервное заболевание, которое проявляется нервными припадками с потерей «сознания» и чувствительности. Различают первичную,
Функциональные взаимосвязи в организме беременной суки
Функциональные взаимосвязи в организме беременной суки В организме суки при беременности возникает целый комплекс взаимовлияний. Также значительно изменяется обмен веществ и функции всех органов и систем. Функциональные изменения и перестройку организма беременных
Структурно-функциональные уровни организации жизни
Структурно-функциональные уровни организации жизни В биологии различают несколько структурно-функциональных уровней организации живой материи.Молекулярный уровень. Характеризуется биохимическими веществами, составляющими живой организм.Клеточный уровень.
Глава 5. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ КАК ОСНОВА ОРГАНИЗАЦИИ СЛОЖНЫХ ФУНКЦИЙ
Глава 5. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ КАК ОСНОВА ОРГАНИЗАЦИИ СЛОЖНЫХ ФУНКЦИЙ Анализ процессов пищеварения и транспорта на современном уровне и в особенности эволюции этих процессов открывает новые горизонты для понимания принципов организации сложных
5.5. Регуляторные блоки
5.5. Регуляторные блоки В течение долгого времени постулировалась специфичность регуляторных сигналов и рецепторов, воспринимающих эти сигналы. В отношении химических сигналов идея специфичности достигла наибольшего развития. Действительно, эти сигналы, действующие
Глава 6. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ. ЭВОЛЮЦИОННЫЕ И БИОЛОГИЧЕСКИЕ АСПЕКТЫ
Глава 6. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ. ЭВОЛЮЦИОННЫЕ И БИОЛОГИЧЕСКИЕ АСПЕКТЫ При рассмотрении эволюции пищеварения мы сталкиваемся с несколькими на первый взгляд исключающими друг друга закономерностями. В самом деле, в ходе эволюции механизмы, участвующие в
ФУНКЦИОНАЛЬНЫЕ ГРУППЫ
ФУНКЦИОНАЛЬНЫЕ ГРУППЫ Основное внимание в экологических исследованиях до недавних пор уделяли видам, при этом основной упор делался на уникальность каждого отдельного вида. При альтернативном подходе, например при изучении функциональных групп, основное внимание