Глава 4. Сложная клетка
Глава 4. Сложная клетка
Ботаник — это тот, кто умеет давать одинаковые названия одинаковым растениям и разные названия разным, причем так, чтобы в этом мог разобраться каждый”, - писал великий шведский систематик Карл Линней (сам ботаник). Это определение может поразить нас скромностью, но именно Линней, разработав классификацию живой природы, основанную на признаках видов, заложил фундамент современной биологии. Он, несомненно, гордился своими достижениями. “Бог творит, Линней упорядочивает”, — любил он говорить (и наверняка счел бы вполне справедливым, что ученые и сегодня пользуются его системой, в которой все живое разделено на царства, классы, роды и виды).
Это стремление к систематизации, к извлечению порядка из хаоса, помогает нам разбираться в мире и лежит у истоков целого ряда естественных наук. Чем была бы химия без периодической системы элементов? Или геология без эр, периодов и эпох? Но у биологии в этом отношении есть одно существенное отличие: здесь классификация по-прежнему остается предметом одного из главных направлений исследований.
О том, как именно устроено “древо жизни”, великая схема, отражающая родство всех живых организмов, по-прежнему ожесточенно спорят ученые, которые в остальном могут быть весьма сдержанными людьми. Название одной из статей Форда Дулиттла, вежливейшего из ученых, передает характерный настрой: “С топором на древо жизни”.
Проблема не в каких-то сокровенных тонкостях, а в одном из самых что ни на есть ключевых разделений. Для большинства из нас, как некогда для Линнея, по-прежнему естественно разделять все в природе на животных, растения и минералы. В конце концов, ничего, кроме них, мы вокруг и не видим. И что может отличаться сильнее? Животные носятся повсюду благодаря своей сложной нервной системе и питаются растениями и другими животными. Растения изготавливают собственные строительные материалы из углекислого газа и воды за счет энергии солнечного света. Они не двигаются с места и не нуждаются ни в каких мозгах. А минералы явно не живые, хотя их рост и убедил в свое время Линнея (впрочем, не без колебаний) включить и их в свою классификацию.
Биология еще в начале своего развития разделилась на зоологию и ботанику, и в течение жизни не одного поколения ученых этим двум дисциплинам было не сойтись. Даже открытие микроскопических форм жизни не разрушило этого давнего разделения. “Животные-крошки” (“анималькули”) вроде амеб, способные двигаться, были отнесены к царству животных и впоследствии получили название “простейшие животные” (Protozoa), в то время как пигментированные водоросли и бактерии были включены в царство растений. Хотя Линней, должно быть, порадовался бы, узнав, что его система по-прежнему в ходу, его бы наверняка поразило, насколько его ввела в заблуждение обманчивая внешность. Расстояние между растениями и животными теперь довольно невелико, в то время как между бактериями и более сложными формами жизни нам открылась зияющая пропасть. Именно преодоление этой пропасти вызывает у ученых столько разногласий. Как именно жизнь перешла от примитивной простоты бактерий к сложности растений и животных? Был ли этот переход изначально вероятен — или же чудовищно маловероятен? Могло ли это случиться и на других планетах, или же мы (скорее всего) одиноки во Вселенной?
Чтобы эта неопределенность не играла на руку тем, кто хотел бы “добавить немного Бога” для ее устранения, следует отметить, что в правдоподобных версиях механизма такого перехода недостатка нет. Проблема здесь в данных, которые смогли бы подтвердить или опровергнуть те или иные версии, особенно в интерпретации данных, относящихся к тому давнему времени (вероятно, около двух миллиардов лет назад), когда, по-видимому, появились первые сложные клетки. Главный вопрос — почему сложные формы жизни возникли за всю историю нашей планеты лишь однажды. Все растения и животные, несомненно, родственны друг другу, то есть у всех был некий общий предок. Сложные формы жизни не развивались из бактерий неоднократно и разновременно (растения от одной разновидности бактерий, животные — от второй, грибы и водоросли — от третьей). Напротив, сложная клетка возникла из бактериальной один-единственный раз, и потомки этой клетки основали все великие царства сложной жизни: растения, животные, грибы и водоросли. И эта клетка-прародительница, предок всех сложных форм жизни, сильно отличалась от бактерий. Когда мы воображаем древо жизни, мы обычно помещаем бактерий у его основания, а хорошо знакомые нам сложные организмы “развешиваем” на ветвях. Но каким тогда был ствол? Хотя мы можем составить его из предполагаемых промежуточных форм вроде одноклеточных протистов (например амеб), на самом деле они во многих отношениях почти столь же сложны, как растения и животные. Они, разумеется, “висят” на ветвях пониже, но все же довольно далеко от ствола.
Пропасть, разделяющая бактерий и всех остальных существ, обусловлена устройством организмов на клеточном уровне. Клетки бактерий, по крайней мере в отношении морфологии (формы, размеров и содержимого), довольно просты. Форма у них незамысловатая, чаще всего шарообразная или палочковидная. Она поддерживается за счет наружной жесткой клеточной стенки. Если же мы заглянем внутрь такой клетки (даже через электронный микроскоп), мы найдем не так уж много структур. По своему строению бактерии сводятся к минимально сложному уровню, совместимому с образом жизни свободноживущего организма. В их клетках все безжалостно рационализировано и нацелено на быстрое размножение. Многие из них хранят лишь столько генов, сколько им жизненно необходимо хранить, и склонны в трудных ситуациях пополнять свои генетические ресурсы, прихватывая гены у других бактерий, а затем, при первой же возможности, снова от этих генов избавляться. Недлинные геномы быстро копируются. Некоторые бактерии могут делиться каждые двадцать минут. Это позволяет им в геометрической прогрессии и с поразительной скоростью размножаться, пока хватает сырья. Если дать одной-единственной бактерии весом в одну триллионную грамма достаточно ресурсов (что, конечно, невозможно), менее чем за два дня она смогла бы произвести популяцию, которая весила бы столько же, сколько земной шар.
Рассмотрим теперь сложные клетки тех организмов, которые называют эукариотами. Мне жаль, что им не дали другого, более приятного названия, потому что по важности с ними не сравнится никто. Все сколько-нибудь солидные организмы на нашей планете, то есть все сложные формы жизни, о которых мы только что говорили, суть эукариоты. Этот термин происходит от греческой приставки эу-, означающей “настоящий”, и греческого же слова “карион”, означающего ядро ореха — или клетки. Действительно, в клетках эукариот имеется настоящее ядро. Этим они и отличаются от клеток бактерий, которых из-за отсутствия ядра называют прокариотами. Приставка про- здесь в некотором роде оценочное суждение: оно предполагает, что прокариоты возникли прежде эукариот. По-моему, это почти наверняка так, хотя некоторые (немногие!) исследователи со мной не согласятся. Так или иначе, независимо от того, когда возникло клеточное ядро, именно его присутствие составляет главное отличие эукариотических клеток. Не стоит и пытаться объяснить их появление в ходе эволюции, не разобравшись, как и почему их клетки обзавелись ядром, и наоборот, почему все известные нам бактерии никогда настоящим ядром не обзавелись.
Ядро представляет собой “центр управления” клеткой. Именно в него упакована ДНК, в молекулах которой записаны гены. Эукариоты отличаются от бактерий не только наличием ядра, но и несколькими связанными с ним особенностями. В их клетках, в отличие от клеток бактерий, не одна кольцевая хромосома, а несколько линейных хромосом, часто имеющихся в клетке в удвоенном числе. Сами гены в хромосомах у нас, эукариот, в отличие от бактерий, не идут один за другим, как бусины на нитке, а по каким-то причинам разбиты на кусочки, разделенные участками некодирующей ДНК, и перемежаются другими некодирующими участками. И наконец, у нас гены не “голые”, как у бактерий, а причудливо обвязаны белками, так что докопаться до них так же трудно, как до современного подарка в пластиковой упаковке.

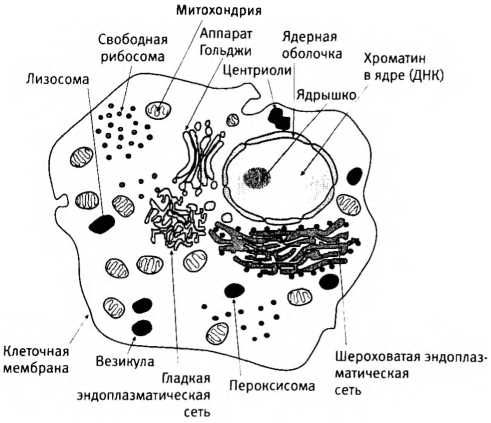
Различия между прокариотическими клетками бактерий и сложными эукариотическими клетками, у которых внутри “много всего” (в том числе ядро, различные органеллы и внутренние системы мембран). Масштаб не соблюден: по объему клетки эукариот в среднем больше клеток бактерий в 10-100 тысяч раз.
Но и помимо ядра эукариотические и прокариотические клетки различаются как небо и земля. Обычно первые гораздо крупнее клеток бактерий и превосходят их по объему в среднем в ю-100 тысяч раз. Кроме того, эукариотические клетки прямо-таки набиты всякой всячиной: стопками окруженных мембраной цистерн, массой пузырьков с различным содержимым, а также подвижными компонентами клеточного скелета, создающими опору для всех внутриклеточных структур и способными одновременно разбираться и отстраиваться заново по всей клетке, позволяя ей менять форму и двигаться. Наверное, самое важное внутри эукариотической клетки — ее органеллы. Эти микроскопические клеточные органы выполняют в клетке различные специализированные функции, совсем как почки или печень в человеческом организме. Важнейшие из них — митохондрии, которые называют электростанциями клетки, потому что они вырабатывают для нее энергию в форме молекул АТФ. В одной эукариотической клетке содержится в среднем около нескольких сотен митохондрий, но в некоторых клетках их может быть и сто тысяч. Далекие предки митохондрий были свободноживущими бактериями, и обсуждению последствий их поглощения эукариотической клеткой будет посвящена значительная часть этой главы.
Но все это внешние отличия. Поведение эукариотических клеток не менее поразительно и принципиально отличается от поведения бактериальных клеток. За немногими исключениями, которые, скажем так, не особенно портят картину, почти всем эукариотам свойствен секс, то есть половое размножение. Они производят половые клетки (сперматозоиды или яйцеклетки), сливающиеся друг с другом и образующие гибридную клетку, которая получает половину генов от отца, а половину — от матери (подробнее мы поговорим об этом в следующей главе). При делении всех эукариотических клеток их хромосомы танцуют удивительный гавот, сходясь попарно и выстраиваясь в ряд в середине веретена из микротрубочек, после чего расходятся в противоположные концы клетки, как танцоры после поклонов и реверансов. Этот список причуд эукариотических клеток можно продолжать, но мне хотелось бы упомянуть еще одну — фагоцитоз: способность заглатывать крупные куски пищи (вплоть до целых клеток) и переваривать их. Судя по всему, это одно из древних свойств эукариот, хотя у некоторых групп, как грибы и растения, оно утрачено. Большинство клеток животных не гоняется за другими клетками, чтобы их съесть, но некоторые клетки иммунной системы занимаются именно этим: поглощают бактерий посредством того же механизма, что и амебы.
Все сказанное относится к клеткам самых разных эукариот, от животных и растений до амеб. Между их клетками есть, разумеется, немало различий, но по сравнению с общими свойствами эти различия представляются несущественными. Так, во многих клетках растений содержатся хлоропласты — органеллы, ответственные за фотосинтез. Древними предками хлоропластов, как и митохондрий, были свободноживущие бактерии (в случае хлоропластов — цианобактерии), проглоченные целиком общим предком растений и водорослей. По каким-то причинам этот предок не переварил обед и в результате обзавелся усовершенствованием, сделавшим его самого и потомков самодостаточными: возможностью получать энергию от солнца, питаясь водой и углекислым газом. Так благодаря одному глотку была запущена вся последовательность событий, которые привели к разделению статичного мира растений и динамичного мира животных. Но, заглянув в растительную клетку, можно убедиться, что, наряду с этим отличием, у нее и у животной клетки имеются тысячи общих черт. Есть и другие примеры. Растения и грибы научились, подобно бактериям, строить наружные клеточные стенки. В некоторых растительных клетках есть вакуоли, и так далее. Но все это ничто по сравнению с бездонным провалом, отделяющим эукариотические клетки от бактерий.
Сам этот провал — одновременно реальный и воображаемый — требует внимательного рассмотрения. Почти по всем признакам между бактериями и эукариотическими клетками наблюдается хоть какое-то перекрывание. Существует некоторое число крупных бактерий и немало крошечных эукариот, так что диапазоны размеров тех и других вполне перекрываются. У бактерий имеется не только клеточная стенка, но и внутренний клеточный скелет из волокон, очень похожих на волокна, образующие клеточный скелет у эукариот. Причем бактериальному скелету, похоже, свойственна даже определенная подвижность. Существуют бактерии и с линейными (не замкнутыми в кольцо) хромосомами, и с напоминающими ядро структурами, и с внутриклеточными мембранами. У некоторых бактерий нет даже клеточной стенки, по крайней мере на одном из этапов жизненного цикла. Некоторые образуют сложные колонии, и их, при желании, можно назвать многоклеточными организмами. Известны один или два случая, когда внутри клеток одних бактерий живут другие, еще меньшего размера, хотя ни у одной бактерии до сих пор не отмечена способность проглатывать другие клетки путем фагоцитоза. У меня складывается ощущение, что бактерии сделали первые шаги в направлениях почти всех эукариотических признаков, но затем остановились, по той или иной причине не сумев продолжить эти эксперименты.
Может показаться, и не без оснований, что перекрывание означает непрерывный переход, а значит, и объяснять тут нечего. Если от простых бактерий до всего спектра сложных эукариот существует непрерывный мост, ни о какой пропасти между теми и другими говорить не приходится. Что-то в этом есть. Но, думаю, это все же неверный вывод, потому что хотя некоторое перекрывание и в самом деле наблюдается, в действительности оно касается двух отдельных спектров — усеченного у бактерий (от “крайней простоты” до “ограниченной сложности”) и гораздо более протяженного у эукариот (от “ограниченной сложности” до “умопомрачительной сложности”). Да, перекрывание есть, но бактериям не удалось особенно продвинуться по эукариотической части этого непрерывного ряда.
Об огромной разнице между прокариотами и эукариотами недвусмысленно свидетельствует история. В течение первых трех миллиардов лет жизни на Земле или около того (с четырех миллиардов до одного миллиарда лет назад) главную роль в ней играли бактерии. Они совершенно изменили мир, в котором жили, хотя сами при этом почти не изменились. Вызванные ими изменения среды были столь грандиозны, что даже нам, людям, сложно осознать их масштабы. Так, например, весь кислород в земной атмосфере происходит из фотосинтеза, который сначала осуществляли исключительно цианобактерии. Свершившаяся около 2,2 миллиарда лет назад “кислородная революция”, в результате которой атмосфера и освещенные солнцем поверхностные воды океана оказались насыщены кислородом, навсегда преобразила облик планеты, но на бактерий эта перемена не произвела особого впечатления. Она вызвала у них лишь сдвиги в экологии в сторону кислородолюбивых форм. Одни разновидности бактерий получили преимущество перед другими, но при этом они остались во всех отношениях бактериями. То же самое относится к другим монументальным переменам условий среды. Из-за деятельности бактерий океанские глубины несколько миллиардов лет были насыщены сероводородом, но бактерии оставались бактериями. Из-за деятельности бактерий окислился атмосферный метан, вызвав глобальное оледенение — первый период “Земли-снежка”, но они по-прежнему оставались бактериями. Еще одним изменением — может быть, самым существенным из всех — было развитие сложных многоклеточных эукариот, продолжающееся последние шестьсот миллионов лет. Эукариоты позволили бактериям освоить новые способы существования, например дали им возможность вызывать инфекционные заболевания, но все-таки бактерии — это по-прежнему бактерии. Нет ничего консервативнее, чем бактерия.
Настоящая история началась с эукариот. Лишь с их появлением жизнь стала по-настоящему сложной и перестала быть бесконечной чередой одного и того же. В некоторых случаях события происходили чертовски быстро. Так называемый кембрийский взрыв, например, — дело рук эукариот. В тот момент (геологический момент, занявший, может быть, пару миллионов лет) впервые в палеонтологической летописи внезапно материализовались крупные животные. Они отнюдь не отличались морфологической скромностью, составляя вовсе не какой-то ряд безликих червей, а поразительную процессию удивительных планов строения, иным из которых предстояло исчезнуть почти так же быстро, как они появились. Все это выглядело так, будто некий безумный творец вдруг пробудился ото сна и немедленно приступил к делу, пытаясь разом отработать миллиарды упущенных лет.
В научной литературе такого рода взрывы называют термином “эволюционная радиация”. Во время таких радиаций та или иная жизненная форма по какой-либо причине переживает относительно непродолжительный период бурной эволюции. “Лучи” возникших на ее основе новых форм отходят от предковой формы, как спицы от оси колеса. “Кембрийский взрыв” — самый известный пример “эволюционной радиации”. Можно привести немало других примеров: освоение суши, появление цветковых растений, распространение злаков, диверсификация млекопитающих — и это далеко не все. Такие события обычно происходят, когда перспективные генетические достижения совпадают с возможностями среды (например, вслед за массовыми вымираниями). Но чем бы они ни были вызваны, “эволюционные радиации” — прерогатива эукариот. Каждый раз, когда происходило нечто подобное, наблюдался очередной расцвет эукариотических организмов. Бактерии же оставались бактериями. Из этого приходится сделать вывод, что человеческий интеллект, сознание, веете качества, которые нам так дороги и проявления которых мы ищем в других уголках Вселенной, просто не могли развиться у бактерий, по крайней мере на Земле: это исключительно эукариотические свойства.
Это должно подействовать на нас отрезвляюще. Хотя бактерии и затмевают нас, эукариот, изобретательностью своей биохимии, их морфологический потенциал следует признать весьма ограниченным. Судя по всему, они не способны породить чудеса, которые мы наблюдаем вокруг, будь то цветок гибискуса или полет колибри. А это делает переход от простых бактерий к сложным эукариотам, наверное, самым важным из всех переходов в истории нашей планеты.
Дарвинисты недолюбливают резкие переходы. Концепция естественного отбора как последовательности очень маленьких усовершенствований предполагает, что мы должны видеть намного больше переходных форм, чем наблюдаем в действительности. Это кажущееся затруднение обсуждал сам Дарвин в “Происхождении видов”. Он отмечал, что все переходные формы по определению не так хорошо приспособлены, как те “конечные”, которые окружают нас сегодня. В силу самой природы отбора менее приспособленные организмы будут проигрывать более приспособленным. Очевидно, что птица, способная к полноценному полету, окажется в выигрыше по сравнению с любыми своими родичами с культями вместо крыльев. То же самое происходит, когда новые компьютерные программы вытесняют с рынка устаревающие: когда вам в последний раз доводилось видеть операционную систему Windows 286 или 386? Когда-то они отражали современный уровень развития, точно так же, как, должно быть, отражали его в свое время и недоразвитые приспособления к полету (каковыми белки-летяги и планирующие древесные змеи обходятся и сегодня).
Но со временем эти ранние версии “операционных систем” исчезли без следа, оставив кажущуюся “пропасть” на месте переходных форм, из которых развилась, скажем, Windows ХР1. Мы понимаем, что операционные системы Windows со временем совершенствовались, но если мы будем искать свидетельства их эволюции, просто сравнивая системы, установленные на компьютерах сегодня, мы мало что сможем найти, не раскопав где-нибудь на чердаках старые модели. Похожим образом обстоят дела и в живой природе: если мы хотим найти свидетельства непрерывного ряда переходов, нам придется обратиться к ископаемым, сохранившимся с тех времен, когда происходили интересующие нас изменения.
Палеонтологическая летопись, разумеется, неполна, но она содержит гораздо больше переходных форм, чем готовы признать не столь многочисленные, но громкоголосые фанатики. Во времена Дарвина действительно существовала проблема “недостающего звена” между обезьянами и людьми: тогда еще не были известны никакие из ископаемых гоминид, промежуточные по своим чертам. Но за последние полвека палеонтологи откопали десятки таких ископаемых. В целом они занимают именно те места в спектре признаков (размеры мозга или степень развития двуногости), каких от них и можно было ожидать. Здесь не только не приходится говорить об отсутствии переходных форм, но даже следует признать их затруднительную избыточность. Трудность состоит в том, что нам сложно выяснить, от каких гоминид произошли современные люди, а какие просто вымерли. Поскольку мы не знаем (пока) все ответы на подобные вопросы, нам по-прежнему приходится слышать громкие заявления, что недостающее звено не найдено, чего по совести сказать нельзя.
Но я, будучи биохимиком, смотрю на ископаемые как на красивые игрушки, только отвлекающие нас от дела. Учитывая, насколько маловероятно и непредсказуемо успешное захоронение, как исключительно трудно сохраниться в виде окаменелостей мягкотелым созданиям вроде медуз, а также растениям и животным, обитающим на суше, ископаемые в принципе не могли сохранить для нас безукоризненную летопись. Если бы они ее сохранили, нам следовало бы заподозрить обман. Те редкие случаи, когда им что-то удалось, следует воспринимать как удивительные подарки судьбы, редкое стечение обстоятельств, граничащее с чудом, но в итоге не более чем приятное дополнение к подлинным свидетельствам естественного отбора. Подлинные же свидетельства окружают нас повсюду: в наш век геномных исследований их дают нам последовательности ДНК-букв в генах.
Эти последовательности гораздо лучше отражают ход эволюции, чем все, что могут нам дать ископаемые. Возьмите любой ген — какой угодно. Его последовательность состоит из длинного ряда букв, порядок которых определяет последовательность аминокислот в белке. Один белок обычно содержит несколько сотен аминокислот, каждая из которых кодируется триплетом ДНК-букв (см. главу 2). Как мы уже отмечали, гены эукариот часто включают длинные вставки некодирующих последовательностей, перемежающихся с более короткими кодирующими. В сумме последовательность одного гена обычно состоит из нескольких тысяч букв. Генов (каждый из них имеет подобное строение) в геноме десятки тысяч. Таким образом, весь геном представляет собой последовательность из миллионов или миллиардов букв, порядок которых может очень многое сказать нам об эволюционном наследии его владельца.
Одни и те же гены, кодирующие белки, выполняющие одни и те же функции, можно найти у множества видов, от бактерий до человека. Вредные мутации, возникающие время от времени в их последовательностях, в ходе эволюции отсеиваются отбором. В результате на соответствующих местах остаются одни и те же буквы. В чисто практическом плане это означает, что мы по-прежнему можем узнать родственные гены, имеющиеся у разных видов, даже если их общий предок жил невообразимо давно. Однако, как правило, лишь малая доля из многих тысяч букв каждого гена по-настоящему важна, а все остальные могут более или менее свободно меняться по мере накопления мутаций, потому что изменения в этих буквах не имеют серьезного значения и не отсеиваются отбором. Чем больше проходит времени, тем больше таких мутаций накапливается и тем сильнее отличаются друг от друга две последовательности любого гена у разошедшихся видов. Виды, общий предок которых жил сравнительно недавно, например шимпанзе и человек, имеют немало общего в последовательностях своих генов, в то время как виды, последний общий предок которых жил намного раньше, например нарцисс и человек, обнаруживают уже меньше общих черт. Здесь работает примерно тот же принцип, что и с языками, которые со временем постепенно расходятся, утрачивая признаки, указывающие на общее происхождение, за исключением некоторых скрытых черт, которые их по-прежнему объединяют.
Генетические деревья основаны на различиях между видами в последовательностях генов. Хотя в накоплении мутаций есть элемент случайности, он уравновешивается за счет тысяч задействованных букв, позволяющих с помощью статистических вероятностей определять степени родства. Пользуясь одним-единственным геном, мы можем реконструировать генеалогическое древо всех эукариот с такой степенью точности, какая не снилась охотникам за ископаемыми. А если у нас возникнут какие-либо сомнения, мы можем просто повторить проведенный анализ, воспользовавшись другим геном, и проверить, получим ли мы в итоге ту же самую схему. Поскольку у разных эукариотических организмов имеются сотни, если не тысячи, общих генов, этот метод можно применять многократно, накладывая получаемые деревья одно на другое. Несложная компьютерная программа позволяет построить на их основе единое “консенсусное” древо, отражающее наиболее вероятные степени родства между всеми эукариотами. По сравнению с палеонтологической летописью с ее пробелами этот метод — настоящий клад. Он позволяет точно определять степень своего родства с растениями, грибами, водорослями и так далее. Дарвин ничего не знал о генах, но именно их тонкое строение более, чем что-либо, позволило заполнить белые места в дарвинистской картине мира.
Все это замечательно, но с этим подходом связаны и некоторые трудности. Одна из основных причин — статистические ошибки при замере изменений, происходивших за огромные промежутки времени. Беда здесь прежде всего в том, что ДНК состоит всего из четырех букв, и в результате мутаций (по крайней мере, мутаций того типа, который нас сейчас интересует) одна буква обычно меняется на другую. Если большинство букв заменялось лишь однажды, то все в порядке, но за длительные периоды эволюции многие буквы неизбежно должны были заменяться неоднократно. Каждое такое изменение — лотерея, поэтому трудно определить, менялась ли та или иная буква один раз, пять раз, десять раз. А если буква осталась прежней, это могло получиться как в случае, если она ни разу не менялась, так и в случае, если она менялась неоднократно: ведь с вероятностью 25 % каждая замена могла приводить к восстановлению исходной буквы. Поскольку анализ таких изменений основан на статистической вероятности, наступает момент, когда мы уже не в состоянии сделать выбор между альтернативными возможностями. К несчастью, тот момент, где мы начинаем барахтаться в море статистической недостоверности, примерно соответствует появлению самой эукариотической клетки. Принципиально важный переход от бактерий к эукариотам скрывается в волнах генетической неопределенности. Единственный способ решить эту проблему — использовать более мелкое статистическое сито, то есть тщательнее отбирать гены для наших исследований.
Гены эукариотических клеток можно разделить на два больших класса: соответствующие бактериальным генам и свойственные, судя по всему, исключительно эукариотам, то есть гены, ничего похожего на которые у бактерий пока обнаружено не было2. Последние получили название “характерные гены эукариот” (eukaryotic signature genes), и их происхождение служит предметом ожесточенных споров. По мнению некоторых исследователей, существование этих генов доказывает, что эукариоты — организмы столь же почтенного возраста, как и бактерии. Сторонники этого вывода утверждают, что раз у эукариот столь много особых генов, значит, они точно начали отдаляться от бактерий в самые древние времена. Если учитывать, что скорость расхождения этих двух эволюционных линий оставалась постоянной (постепенное “тиканье” мутаций, играющее роль молекулярных часов), то масштабы накопленных различий заставляют нас заключить, что эукариоты старше пяти миллиардов лет, то есть по крайней мере на полмиллиарда лет старше самой Земли. Что-то тут не так.
Другие исследователи утверждают, что характерные гены эукариот ничего не говорят нам об эволюционном наследии этой группы, потому что у нас нет никакой возможности узнать, с какой скоростью гены могли эволюционировать в далеком прошлом, и нет никаких оснований предполагать, что их эволюционное расхождение должно было работать как часы. Более того, нам известно, что в настоящее время одни гены эволюционируют быстрее, чем другие. А тот факт, что молекулярные часы указывают на столь сомнительную древность эукариот, заставляет предположить, что либо жизнь была занесена на Землю из космоса (на мой взгляд, дешевая уловка), либо наши часы показывают неправильное время. Почему они могут быть настолько неточны? Потому что скорость, с которой эволюционируют гены, зависит от множества обстоятельств, прежде всего от особенностей организма, в котором они находятся. Как мы уже убедились, бактерии — неисправимые консерваторы, они всегда остаются бактериями, в то время как эукариоты, судя по всему, склонны время от времени испытывать впечатляющие перемены, как было, например, во времена “кембрийского взрыва”. Возможно, самым драматичным из подобных периодов (с точки зрения генов) было возникновение самой эукариотической клетки, а если так, то у нас есть все основания предполагать, что в те давние времена скорость эволюционных изменений была просто бешеной. Если эукариоты возникли позже бактерий (как полагает большинство исследователей), то их гены так сильно отличаются от бактериальных потому, что в течение некоторого времени они эволюционировали очень быстро, без конца претерпевая мутации, рекомбинации, дупликации и вновь мутации.
Так что характерные гены эукариот не так уж много могут рассказать нам об их эволюции. Эти гены эволюционировали так быстро и так основательно, что их происхождение просто теряется во тьме веков. А что же второй класс генов, для которых известны соответствующие гены бактерий? Они намного “разговорчивее”, потому что позволяют непосредственно сравнивать подобное с подобным. Гены, которые можно найти как у бактерий, так и у эукариот, часто отвечают за ключевые функции клетки, будь то ключевые процессы обмена веществ (механизмы выработки энергии и использования ее для изготовления главных “строительных блоков” всего живого, таких как аминокислоты и липиды) или ключевые информационные процессы (механизмы считывания с ДНК информации и ее трансляции, то есть перевода на действующий язык белков). Подобные базовые процессы обычно эволюционируют медленно, потому что от них зависит очень многое. Стоит поменять хоть что-нибудь в механизме синтеза белков, и эти изменения затронут все белки, а не только какой-то один. И точно так же стоит лишь слегка изменить механизм выработки энергии, и это может поставить под угрозу всю работу клетки. Поскольку изменения в ключевых генах имеют особенно много шансов отсеяться в ходе отбора, они эволюционируют медленно и потому должны давать нам более тонкий инструмент для изучения эволюции. Таким образом, существует принципиальная возможность того, что древо, построенное на основе таких генов, прольет свет на родство эукариот с бактериями, указав нам, из какой группы бактерий возникли эукариоты, а также, может быть, и подсказав, как они могли возникнуть.
Подобное дерево первым построил американский микробиолог Карл Везе в конце 70-х годов. Он выбрал ген, ответственный за один из ключевых информационных процессов, происходящих в клетке, — кодирующий одну из деталей крошечных молекулярных машин, так называемых рибосом, которые осуществляют синтез белков. В силу технических причин Везе первоначально использовал не сам ген, а его РНК-копию, которая считывается с данного гена и непосредственно встраивается в рибосому. Он выделял рибосомальную РНК (рРНК) из различных бактерий и эукариот, прочитывал ее последовательность и строил дерево на основе сравнения таких последовательностей, свойственных разным организмам. Результаты этого исследования вызвали потрясение, поставив под сомнение общепризнанные взгляды на систему органического мира.
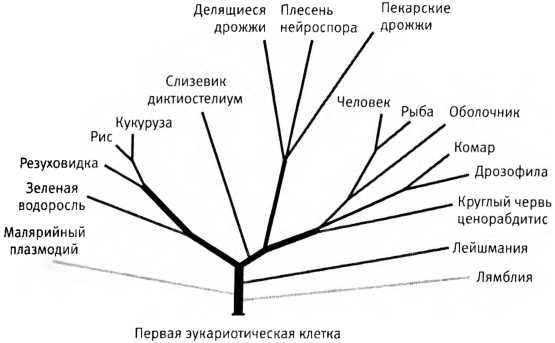
Обычное древо жизни, изображающее происхождение эукариот от общего предка — одноклеточного организма, жившего, вероятно, около двух миллиардов лет назад. Чем длиннее ветвь, тем больше эволюционное расстояние, то есть тем сильнее отличаются гены.
Везе открыл, что все живое на нашей планете делится на три больших группы, соответствующие трем ветвям древа жизни. Первая из этих групп — бактерии (чего и следовало ожидать), вторая — эукариоты. Но существование третьей группы, представителей которой теперь называют археями и которая с тех пор прочно заняла место на мировой арене, оказалось полной неожиданностью. Хотя немногочисленные виды архей были известны уже лет сто, до появления нового дерева, построенного Везе, их рассматривали как небольшую фракцию в составе бактерий. После открытия Везе они оказались такой же важной группой, как эукариоты, несмотря на то, что выглядят они в точности как бактерии. Они очень малы, обычно имеют наружную клеточную стенку, не имеют ядра (и никаких других примечательных внутренних структур) и никогда не образуют колоний, которые можно было бы принять за многоклеточные организмы. Придание им столь большой важности показалось многим ученым дерзкой попыткой перекроить мир, низведя все изумительное разнообразие растений, животных, грибов, водорослей и протистов к незначительным ответвлениям на древе, где главенствующее положение заняли прокариоты. Везе уверял, что все многообра-
*** Страницы 162-163 отсутствуют ***
Проблема схемы Везе в том, что она построена на основании единственного гена и потому лишена статистической надежности схем, получаемых наложением разных деревьев. На эту схему стоит полагаться лишь в том случае, если мы можем быть уверены, что выбранный для нее ген действительно отражает наследие эукариотических клеток. Лучший способ проверить, так ли это, состоит в том, чтобы наложить на эту схему другие, полученные на основе иных медленно эволюционирующих генов, и выяснить, будет ли воспроизведено и на них разделение на три большие ветви. Но результаты подобных проверок открывают нам парадоксальную картину. Если выбрать лишь гены, имеющиеся у представителей всех трех больших ветвей жизни (бактерий, архей и эукариот), надежные деревья удается построить только для бактерий и архей, но не для эукариот. Они, как выясняется, представляют собой странную смесь. Получается, что одни из наших генов происходят от архей, а другие — от бактерий. Чем больше генов мы изучаем (а в одном недавнем исследовании для построения нового “супердерева” были использованы данные 5-700 генов, взятых у 165 разных видов), тем яснее становится, что эукариотические клетки произошли не обычным “дарвиновским” путем, а посредством какого-то чудовищного объединения генов. С генетической точки зрения первый эукариотический организм был химерой — полуархеей-полубактерией.
По Дарвину, все живое развивается путем медленного накопления различий, возникающих по мере того, как эволюционные линии расходятся, отделившись от общего предка. Результатом этого процесса оказывается ветвящееся дерево, и нет никаких сомнений, что именно такие деревья лучше всего отображают эволюцию большинства организмов, которых видно невооруженным глазом, то есть, по сути, большинства крупных эукариот. Но так же ясно, что деревья — не лучшие схемы для отображения эволюции микробов, будь то бактерии, архей или одноклеточные эукариоты.
Есть два процесса, вступающие в противоречие с дарвиновскими генеалогическими деревьями: горизонтальный перенос генов и слияние целых геномов. Специалисты по эволюции микробов, пытающиеся разобраться во взаимоотношениях бактерий и архей, с досадной регулярностью сталкиваются с горизонтальным переносом генов. Этот несколько неуклюжий термин означает просто передачу генов от одного организма к другому. В результате геном, который материнская клетка бактерии передает дочерней клетке, может быть и таким же, и не таким, как тот, который сама материнская клетка унаследовала от своей “матери”. Некоторые гены, например исследованный Везе ген рибосомальной РНК, обычно передаются вертикально, из поколения в поколение, в то время как другими клетки микробов, часто совершенно неродственные, постоянно обмениваются3. Общая картина оказывается чем-то средним между деревом и сетью, где ключевые гены (как гены рибосомальной РНК) обычно образуют дерево, а для других характерно образование сети. Вопрос, существует ли такой набор ключевых генов, представителями которого микроорганизмы вообще никогда не обмениваются путем горизонтального переноса, по-прежнему спорен. Если таких генов не бывает, то сама идея проследить за ходом эволюции эукариот вплоть до неких конкретных групп прокариот лишена смысла. Те или иные группы организмов могут иметь определенное происхождение лишь в том случае, если они наследуют признаки непосредственно от собственных предков, а не от каких-то других случайно подвернувшихся групп. Но что если небольшой набор ключевых генов все-таки никогда не передается горизонтально? Что тогда можно сказать о происхождении той или иной группы? Можно ли считать, что кишечная палочка остается кишечной палочкой, если 99 % ее генов случайным образом заменяются другими?4
Со слиянием геномов связаны похожие трудности. Здесь проблема состоит в том, что дарвиновское дерево переворачивается вверх ногами: вместо эволюционного расхождения мы получаем схождение. Тогда возникает вопрос, кто из двух (или большего числа) участников такого схождения отражает истинный ход эволюции? Если проследить наследование одного лишь гена рибосомальной РНК, мы получим обычное ветвящееся дарвиновское древо, но если рассмотреть большое число генов или целые геномы, мы получим что-то вроде кольца, в котором разошедшиеся ветви вновь сходятся и сливаются.
Нет никаких сомнений, что клетки эукариот представляют собой генетические химеры. Данные, свидетельствующие об этом, никто не ставит под сомнение. Вопрос, по поводу которого исследователи теперь разделяются на враждующие фракции, в том, насколько большое значение нужно придавать эволюции по Дарвину, и насколько большое — интенсивным процессам генетического слияния. Иначе говоря, сколько свойств эукариотических клеток возникло путем постепенной эволюции клетки-хозяина, а сколько могло развиться только лишь после генетического слияния. За несколько десятилетий о происхождении эукариотической клетки были выдвинуты десятки теорий, от умозрительных, если не сказать надуманных, до основанных на детальных биохимических реконструкциях. Ни одна из них пока не получила однозначного подтверждения. Все эти теории можно разделить на две большие группы: придающие особое значение постепенному дарвиновскому расхождению и резкому генетическому слиянию. Эти группы соответствуют двум противоборствующим сторонам другого, более давнего спора между биологами: теми, кто доказывает, что эволюция идет путем постепенных непрерывных изменений, и теми, кто настаивает на существовании продолжительных периодов застоя, или равновесия, прерывающихся иногда внезапными впечатляющими изменениями. То есть, как шутили в свое время, споров о том, ползком или прыжками идет эволюция5.
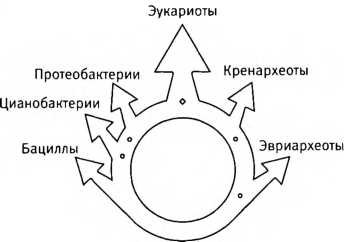
“Кольцо жизни”. Последний общий предок всего живого располагается внизу, где его потомки разделяются на бактерий (слева) и архей (справа). Представители тех и других вновь сливаются вверху, давая начало химерным организмам — эукариотам.
Применительно к эукариотической клетке Кристиан де Дюв назвал представления первой группы гипотезой “примитивного фагоцита”, а второй — гипотезой “судьбоносной встречи”. Идея “примитивного фагоцита” соответствует дарвиновской концепции, и среди ее сторонников выделяются оксфордский эволюционист Том Кавалир-Смит и сам Кристиан де Дюв. В основе этой гипотезы лежит предположение, что предки эукариотических клеток постепенно накопили все признаки, свойственные клеткам современных эукариот: ядро, настоящий половой процесс, клеточный скелет и, что самое важное, способность к фагоцитозу, то есть поглощению других клеток путем изменения формы, окружения их, заглатывания и последующего внутреннего переваривания. Единственной чертой, которой, в отличие от клеток современных эукариот, их предполагаемый непосредственный предок, примитивный фагоцит, не обладал, было наличие митохондрий, которые вырабатывают энергию, используя для этого кислород. Вероятно, ему приходилось получать энергию посредством брожения — гораздо менее эффективного процесса.
Но для фагоцита проглотить предков нынешних митохондрий было тривиальной задачей. Что могло быть проще? Более того, если не путем фагоцитоза, то как могла одна клетка оказаться внутри другой? Обладание митохондриями, разумеется, предоставило примитивному фагоциту важное преимущество (они должны были произвести революцию в его способе получения энергии), но не привело ни к каким принципиальным изменениям его устройства. После того как фагоцит обзавелся митохондриями, он по-прежнему остался фагоцитом, хотя теперь у него появилась возможность получать больше энергии. Однако многие гены из порабощенных митохондрий могли постепенно перейти в ядро и встроиться в геном клетки-хозяина и именно этой передачей объясняется, согласно данной гипотезе, химерная природа современных эукариотических клеток. Их бактериальное наследие составляют гены, взятые у митохондрий. Таким образом, сторонники гипотезы примитивно го фагоцита не оспаривают химерную природу современных эукариот, но предполагают, что предковой для нынешних эукариот клеткой-хозяином, то есть первой настоящей, хотя и примитивной эукариотической клеткой, был нехимерный фагоцит
Том Кавалир-Смит еще в начале 8о-х годов обратил внимание на группу из тысячи с лишним видов одноклеточных эукариот примитивного облика, у которых отсутствуют митохондрии. Он предположил, что какие-то из них могли дожить до наших дней с тех времен, когда возникли первые эукариотические клетки, и могут быть прямыми потомками того примитивного фагоцита, у которого еще не было митохондрий. Если так то они не должны обнаруживать никаких признаков генетической химерности, поскольку их происхождение было обусловлено чисто дарвиновскими процессами. Но в следующие два десятилетия выяснилось, что все эти организмы — химеры. Получалось, что у предков их всех когда-то были митохондрии, впоследствии утраченные или преобразившиеся во что-то другое. Все без исключения современные эукариотические клетки имеют митохондрии или происходят от предков, у которых имелись митохондрии. Если когда-то и существовал примитивный фагоцит, у которого митохондрий не было, он не оставил прямых потомков. Это не означает, что его никогда не существовало, а просто говорит о том, что его существование остается гипотезой.
Вторая группа теорий происхождения эукариотической клетки равняется на знамя “судьбоносной встречи”. Все эти теории предполагают, что в свое время между двумя или несколькими прокариотическими клетками возникла связь того или иного рода, которая привела к формированию сплоченного объединения клеток — химерного организма. Если клетка-хозяин была не фагоцитом, а археей, обладавшей клеточной стенкой, то главный вопрос в том, как другим клеткам вообще удалось проникнуть внутрь? Ведущие сторонники этой идеи, особенно Линн Маргулис и Билл Мартин (с которым мы познакомились в главе 1), указывают на целый ряд возможностей, которые могли к этому привести. Например, Линн Маргулис предположила, что бактерия-хищник могла силой прорываться во внутренности других бактерий (и тому известен ряд примеров). Билл Мартин, напротив, отстаивает идею, что в основе лежала подробно проанализированная им взаимовыгодная метаболическая связь между клетками, каждая из которых обменивалась с другой определенным сырьем6. Если так, то сложно понять, как одна прокариотическая клетка проникла внутрь другой без помощи фагоцитоза, но Мартин приводит два примера, где у бактерий произошло именно это.
Теории “судьбоносной встречи” можно назвать недарвиновскими в том смысле, что они предполагают не эволюцию путем маленьких изменений, а внезапное возникновение совершенно нового организма. Принципиально здесь также представление о том, что все эукариотические признаки развились в ходе эволюции лишь после “судьбоносной встречи” и последовавшего объединения. Сами объединившиеся клетки были чисто прокариотическими: неспособными к фагоцитозу, лишенными настоящего полового процесса, подвижного клеточного скелета, ядра и прочих атрибутов эукариот. Все эти признаки развились лишь после закрепления возникшего союза. Эти версии подразумевают, что в самом таком союзе было нечто, что преобразовало архиконсервативных, вечно неизменных прокариот в свою прямую противоположность: одержимых быстрой ездой, постоянно меняющихся эукариот.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
5.1. Главная ячейка жизни — клетка
5.1. Главная ячейка жизни — клетка Определение жизни с позиций функционального подхода (метаболизм, размножение, расселение в пространстве) можно дать в следующей форме [Печуркин, 1982]: это открытая система, развивающаяся на основе матричного автокатализа под влиянием
Как клетка получает и использует энергию
Как клетка получает и использует энергию Чтобы жить, надо работать. Эта житейская истина вполне приложима к любым живым существам. Все организмы: от одноклеточных микробов до высших животных и человека — непрерывно совершают различные типы работы. Таковы движение, то
Бактериальная клетка в цифрах
Бактериальная клетка в цифрах Благодаря биофизике — одной из отраслей науки, с которой мы уже познакомились в начале этой главы, — были получены весьма интересные данные. Возьмем, например, шаровидную бактериальную клетку диаметром 0,5 мкм. Поверхность такой клетки
Клетка — элементарная частица жизни
Клетка — элементарная частица жизни Эти беглые замечания о способах выработки энергии в клетках многоклеточного организма и в бактериальных клетках акцентируют весьма существенные различия в важнейших аспектах их жизнедеятельности. Несходны эти два класса клеток и
Сложная машина — иммунитет. Но чем сложнее машина, тем более вероятна ее поломка. Так ли это?
Сложная машина — иммунитет. Но чем сложнее машина, тем более вероятна ее поломка. Так ли это? — Сложная эта машина — иммунитет. Врожденные дефекты и старость — Да, не простая. — Но ведь чем сложнее машина, тем более вероятна ее поломка. — Это
7.1. Клетка – структурно-функциональная единица жизни
7.1. Клетка – структурно-функциональная единица жизни Клеточная форма организации свойственна всем живым организмам. Если не касаться вопроса о приложимости понятия «живое» к вирусам (тем более что ответ, пожалуй, следует дать отрицательный), то клетку нужно признать
Глава 2 КЛЕТКА – ОСНОВНАЯ ЕДИНИЦА НЕРВНОЙ ТКАНИ
Глава 2 КЛЕТКА – ОСНОВНАЯ ЕДИНИЦА НЕРВНОЙ ТКАНИ Головной мозг человека состоит из огромного количества разнообразных клеток. Клетка – основная единица биологического организма. Наиболее просто организованные животные могут иметь всего одну клетку. Сложные организмы
ГРУДНАЯ КЛЕТКА
ГРУДНАЯ КЛЕТКА Форма грудной клетки изменяется в зависимости от конституционального типа собаки, степени ее развития и возраста. Грудная клетка, вмещающая дыхательные органы, сердце и главнейшие кровеносные сосуды, должна быть объемистой. Объем груди обусловлен длиной,
Клетка-ловушка
Клетка-ловушка Тебе понадобятся: клетка-ловушка, приманка (зерна, сыр, хлеб, колбаса), доска или черепицаДлительность опыта: 1–2 дня.Время проведения: поздняя осень — ранняя весна. Твои действия:• Купи клетку-ловушку любого типа или сделай ее сам. Для этого возьми
5.2. Кишечная клетка
5.2. Кишечная клетка Схема кишечной клетки представлена на рис. 26. Известно, что численность кишечных клеток составляет 1010, а соматических клеток взрослого человека— 10 15. Следовательно, одна кишечная клетка обеспечивает питание около 100 000 других клеток. Такая
Глава 2. Клетка
Глава 2. Клетка ТЕМЫ• История изучения клетки. Клеточная теория• Химический состав клетки• Строение эукариотической и прокариотической клеток• Реализация наследственной информации в клетке• ВирусыУдивительный и загадочный мир окружает нас, жителей планеты,
10. Эукариотическая клетка. Цитоплазма. Органоиды
10. Эукариотическая клетка. Цитоплазма. Органоиды Вспомните!Каковы основные положения клеточной теории?Какие выделяют типы клеток в зависимости от расположения генетического материала?Назовите известные вам органоиды клетки. Какие функции они выполняют?В § 4 мы уже
12. Прокариотическая клетка
12. Прокариотическая клетка Вспомните!В чём заключаются принципиальные отличия в строении прокариотических и эукариотических клеток?Какова роль бактерий в природе?Разнообразие прокариот. Царство прокариот в основном представлено бактериями, наиболее древними