Глава 3. Фотосинтез
Глава 3. Фотосинтез
Представьте себе планету без фотосинтеза. Начнем с того, что она не была бы зеленой. Изумрудный цвет Земли — символ торжества растений и водорослей, а в конечном счете — их зеленых пигментов, улавливающих свет для фотосинтеза. Первейший из этих пигментов — великий преобразователь хлорофилл, умеющий ловить лучи света и превращать их в кванты химической энергии, за счет которой живут как растения, так и животные.
Планета, вероятно, не была бы и голубой: лазурь небес и морских пейзажей невозможна без чистого неба и воды, очищенных от тумана и пыли кислородом. А без фотосинтеза свободного кислорода бы не было.
Более того, возможно, что на такой планете не было бы и океанов. Без кислорода в атмосфере не может быть озона, а без озона мало что сдерживало бы обжигающий жар ультрафиолетовых лучей. Они “раскалывают” молекулы воды, образуя кислород и водород. Но кислород при этом образуется медленно, он не накапливался бы в атмосфере, а реагировал с железом, входящим в состав минералов и окрашивающим их в ржаво-рыжий цвет. А водород, самый легкий из всех газов, ускользал бы из поля тяготения и рассеивался в космосе. Этот процесс может быть медленным, но он неумолим и приводит к тому, что океаны словно истекают кровью, сочащейся в космос. Ультрафиолетовое излучение стоило Венере ее океанов, а возможно, и Марсу тоже.
Так что нам потребуется не так уж много воображения, чтобы представить себе планету без фотосинтеза. Она могла бы быть похожа на Марс — пыльную рыжую пустыню без океанов и без явных признаков жизни. Жизнь, разумеется, возможна и без фотосинтеза, и многие астробиологи ищут ее на Марсе. Но даже если под камнями Марса и найдутся немногочисленные спрятавшиеся там бактерии или следы жизни будут обнаружены в толще марсианского льда, сама планета все-таки мертва. Она пребывает почти в полном равновесии — верный признак безжизненности. Марс никак не спутать с Геей.
Кислород — это ключ к жизни на планете. Будучи всего лишь отходами фотосинтеза, именно это вещество делает планету живой. Кислород выделяется в ходе фотосинтеза с такой скоростью, что планета не успевает его поглощать. Когда вся пыль и все железо из минералов, вся сера в воде и весь метан в воздухе, все, что может быть окислено, окисляется, свободный кислород продолжает поступать в океаны и в атмосферу. Насыщая их, он останавливает процесс потери воды. Выделяемый из воды водород неизбежно вновь сталкивается с кислородом, не успевая проложить себе дорогу в космос. Их быстрая реакция вновь приводит к образованию воды, падающей с небес в виде дождя, и останавливает потерю воды океанами. А когда в атмосфере накапливается достаточно кислорода, в ней образуется озоновый слой, который сдерживает обжигающие ультрафиолетовые лучи и делает планету более пригодной для жизни.
Кислород не только оберегает жизнь на планете: он заряжает все живое энергией, позволяя существам достигать крупных размеров. Бактерии прекрасно обходятся без кислорода: эти несравненные мастера электрохимии способны осуществлять реакции между чуть ли не любыми веществами, понемногу добывая для себя энергию. Но общее количество энергии, которую можно получать путем брожения или осуществляя реакции между, например, метаном и сульфатами, ничтожно по сравнению с энергией кислородного дыхания — в буквальном смысле кислородного горения пищи, в ходе которого она полностью окисляется до углекислого газа и водяного пара. Ничто другое не в состоянии давать энергию на все нужды многоклеточной жизни. Все животные и растения не могут жить без кислорода — по крайней мере, на каком-то этапе своих жизненных циклов. Единственное известное мне исключение — микроскопический (но многоклеточный) круглый червь, каким-то образом умудряющийся существовать в неподвижных бескислородных глубинах Черного моря. Так что мир без свободного кислорода мог бы быть лишь микромиром — в том смысле, что был бы населен лишь микроскопическими организмами.
Кислород и другими путями способствует приобретению живыми существами крупных размеров. Представьте себе пищевую цепь. Хищники высшего звена питаются менее крупными животными, которые, в свою очередь, питаются насекомыми, а те — более мелкими насекомыми, которые питаются грибами и травой. Пищевые цепи из пяти-шести звеньев встречаются не так уж редко. При переходе к каждому следующему звену тратится энергия, поскольку КПД ни у одной формы дыхания не может быть стопроцентным. КПД кислородного дыхания составляет около 40 %, в то время как КПД большинства других форм дыхания (в которых вместо кислорода используется, например, железо или сера) менее 10%. Это значит, что если не использовать кислород, количество доступной энергии будет сокращаться до 1% от исходного всего за пару переходов между звеньями пищевой цепи, в то время как при кислородном дыхании сокращение до 1% будет занимать пять пежий цвет. А водород, самый легкий из всех газов, ускользал бы из поля тяготения и рассеивался в космосе. Этот процесс может быть медленным, но он неумолим и приводит к тому, что океаны словно истекают кровью, сочащейся в космос. Ультрафиолетовое излучение стоило Венере ее океанов, а возможно, и Марсу тоже.
Так что нам потребуется не так уж много воображения, чтобы представить себе планету без фотосинтеза. Она могла бы быть похожа на Марс — пыльную рыжую пустыню без океанов и без явных признаков жизни. Жизнь, разумеется, возможна и без фотосинтеза, и многие астробиологи ищут ее на Марсе. Но даже если под камнями Марса и найдутся немногочисленные спрятавшиеся там бактерии или следы жизни будут обнаружены в толще марсианского льда, сама планета все-таки мертва. Она пребывает почти в полном равновесии — верный признак безжизненности. Марс никак не спутать с Геей.
Кислород — это ключ к жизни на планете. Будучи всего лишь отходами фотосинтеза, именно это вещество делает планету живой. Кислород выделяется в ходе фотосинтеза с такой скоростью, что планета не успевает его поглощать. Когда вся пыль и все железо из минералов, вся сера в воде и весь метан в воздухе, все, что может быть окислено, окисляется, свободный кислород продолжает поступать в океаны и в атмосферу. Насыщая их, он останавливает процесс потери воды. Выделяемый из воды водород неизбежно вновь сталкивается с кислородом, не успевая проложить себе дорогу в космос. Их быстрая реакция вновь приводит к образованию воды, падающей с небес в виде дождя, и останавливает потерю воды океанами. А когда в атмосфере накапливается достаточно кислорода, в ней образуется озоновый слой, который сдерживает обжигающие ультрафиолетовые лучи и делает планету более пригодной для жизни.
Кислород не только оберегает жизнь на планете: он заряжает все живое энергией, позволяя существам достигать крупных размеров. Бактерии прекрасно обходятся без кислорода: эти несравненные мастера электрохимии способны осуществлять реакции между чуть ли не любыми веществами, понемногу добывая для себя энергию. Но общее количество энергии, которую можно получать путем брожения или осуществляя реакции между, например, метаном и сульфатами, ничтожно по сравнению с энергией кислородного дыхания — в буквальном смысле кислородного горения пищи, в ходе которого она полностью окисляется до углекислого газа и водяного пара. Ничто другое не в состоянии давать энергию на все нужды многоклеточной жизни. Все животные и растения не могут жить без кислорода — по крайней мере, на каком-то этапе своих жизненных циклов. Единственное известное мне исключение — микроскопический (но многоклеточный) круглый червь, каким-то образом умудряющийся существовать в неподвижных бескислородных глубинах Черного моря. Так что мир без свободного кислорода мог бы быть лишь микромиром — в том смысле, что был бы населен лишь микроскопическими организмами.
Кислород и другими путями способствует приобретению живыми существами крупных размеров. Представьте себе пищевую цепь. Хищники высшего звена питаются менее крупными животными, которые, в свою очередь, питаются насекомыми, а те — более мелкими насекомыми, которые питаются грибами и травой. Пищевые цепи из пяти-шести звеньев встречаются не так уж редко. При переходе к каждому следующему звену тратится энергия, поскольку КПД ни у одной формы дыхания не может быть стопроцентным. КПД кислородного дыхания составляет около 40 %, в то время как КПД большинства других форм дыхания (в которых вместо кислорода используется, например, железо или сера) менее 10 %. Это значит, что если не использовать кислород, количество доступной энергии будет сокращаться до 1 % от исходного всего за пару переходов между звеньями пищевой цепи, в то время как при кислородном дыхании сокращение до 1 % будет занимать пять переходов. Это, в свою очередь, означает, что длинные пищевые цепи возможны только при кислородном дыхании. В силу самого устройства пищевых цепей хищникам сложно было бы существовать в бескислородном мире, потому что без кислорода хищничество как образ жизни просто не окупается.
Хищничество же способствует увеличению животных в размерах, вызывая гонку вооружений между хищниками и их жертвами. От зубов защищают раковины и панцири, укрыться от чужих глаз помогает маскировка, но размерами можно устрашить и охотника, и дичь. Если есть кислород, то хищничество окупается, а если есть хищничество, то окупаются размеры. Так что кислород делает возникновение крупных организмов не только возможным, но и вероятным.
Он также помогает им расти. Белок, дающий телу животного прочность и упругость, называется коллаген. Это главный белок всех соединительных тканей, как обызвествленных (в костях, зубах и панцирях), так и “голых” (в связках, сухожилиях, хрящах и коже). Коллаген существенно превосходит по количеству все остальные белки млекопитающих, составляя целых 25 % общей массы белков в их организмах. Как у позвоночных, так и у беспозвоночных он служит ключевым компонентом всевозможных раковин, кутикул, панцирей и волокнистых тканей — “скотчем” и “клеем” всего животного мира. Коллаген состоит из довольно необычных “строительных блоков”, в которых для формирования перекрестных связей между соседними белковыми волокнами, придающими всей структуре прочность и упругость, требуется свободный кислород. Это означает, что крупные животные, защищенные панцирем или прочным скелетом, могли появиться лишь тогда, когда концентрация кислорода в атмосфере достигла достаточного уровня, чтобы обеспечивать синтез коллагена. Возможно, что этот фактор способствовал внезапному появлению крупных животных в палеонтологической летописи в начале кембрийского периода, около 550 миллионов лет назад — вскоре после сильного повышения концентрации кислорода в атмосфере планеты.
Потребность в кислороде для синтеза коллагена может показаться случайностью. Вероятно, вместо коллагена можно было использовать что-либо, для чего не требуется свободный кислород. Действительно ли кислород необходим, чтобы давать животным прочность, или этот ингредиент лишь случайно вошел в состав “строительного раствора” и остался в его рецепте навсегда? Мы не знаем наверняка, но примечательно, что высшим растениям для синтеза их структурной опоры — необычайно прочного полимера лигнина, придающего древесине гибкость и прочность, — тоже требуется кислород. Лигнин образуется посредством довольно беспорядочного химического процесса, в ходе которого свободный кислород участвует в формировании прочных поперечных связей между цепочками. Эти связи очень сложно разорвать, поэтому-то древесина и отличается такой твердостью и устойчивостью к разложению. Стоит убрать из древесины лигнин (как делают производители бумаги, которым приходится тщательно удалять это вещество из древесной стружки, чтобы изготавливать бумагу), и деревья рухнут под собственной тяжестью, не в силах выдержать даже легкий ветерок.
Так что без кислорода не было бы ни крупных животных, ни высоких растений, ни хищников, ни голубого неба, ни, возможно, океана: вообще ничего, кроме пыли и бактерий. Невозможно представить себе более ценные отходы, чем кислород. Причем эти отходы не только ценны, но и маловероятны. Фотосинтез вполне мог возникнуть в ходе эволюции — у нас, на Земле, или на Марсе, или где угодно еще во Вселенной — в такой форме, что никакого свободного кислорода в результате этого процесса не образовывалось бы. А это почти неизбежно обрекло бы жизнь на вечное пребывание на бактериальном уровне сложности. Если так и происходило на других планетах, мы можем быть единственными разумными существами во Вселенной, населенной бактериями.
Одна из причин, почему кислород мог никогда не начать накапливаться в атмосфере, связана с дыханием. Фотосинтез и дыхание — процессы однотипные, но противоположные. В самых общих чертах фотосинтез — это синтез органических веществ из двух несложных веществ — углекислого газа и воды — за счет энергии солнечного света. А в процессе дыхания мы сжигаем органические вещества (пищу), выделяя углекислый газ и воду обратно в атмосферу, и получаем энергию, за счет которой живем. Вся наша энергия — это энергия солнечного света, освобождаемая нами из плена, в котором она пребывает в пище.
Фотосинтез и дыхание противоположны не только как химические процессы, но и как факторы глобального масштаба. Если бы не было дыхания, то есть никакие животные, грибы и бактерии не сжигали бы растительную пищу, то весь углекислый газ был бы давно высосан из атмосферы и преобразован в биомассу. Тогда жизнь остановилась бы, за исключением того немногого, что смогло бы существовать за счет медленного самопроизвольного разложения органики и за счет углекислого газа, выделяемого вулканами. Но ничего подобного не происходит. А происходит вот что: в процессе дыхания все органические вещества, откладываемые растениями, сжигаются. В геологических масштабах времени кажется, что растения исчезают в дыму. Из этого вытекает одно важное следствие. Весь кислород, выделяемый в атмосферу при фотосинтезе, может вновь поглощаться в процессе дыхания. В результате могло бы сложиться вечное, неизменное, ничем не нарушаемое равновесие, которое для любой планеты было бы смерти подобно. Единственный способ, позволяющий планете приобрести кислородную атмосферу, а значит и единственный способ избежать судьбы Марса и не превратиться в пыльную красную пустыню, состоит в том, чтобы сохранять определенную часть растительной биомассы нетронутой, неподвластной стихиям и неуязвимой для хитроумных изобретений жизни, позволяющих разлагать биомассу, добывая из нее энергию. А для этого ее необходимо хоронить.
Именно это и происходит. Откладываемая растительная биомасса хоронится в виде каменного и бурого угля, нефти, природного газа и пыли, образуя породы, спрятанные глубоко в недрах земли. Согласно результатам революционных исследований геохимика Роберта Бернера, работавшего в Йельском университете, в земной коре залегает примерно в 26 тысяч раз больше "мертвого” органического углерода, чем содержится во всем живом в биосфере. Каждый атом углерода есть нечто обратное молекуле атмосферного кислорода. На всякий атом углерода, который мы добываем из-под земли и сжигаем как топливо, из атмосферы выхватывается одна молекула кислорода и превращается снова в углекислый газ, что приводит к серьезным, хотя и непредсказуемым последствиям для климата. К счастью, нам никогда не удастся истощить запасы атмосферного кислорода, сжигая ископаемое топливо, даже если мы вызовем настоящую климатическую катастрофу: подавляющее большинство органического углерода хоронится в виде микроскопического детрита в таких горных породах, как сланцы, сжигать которые в промышленных масштабах невозможно или, по крайней мере, экономически невыгодно. До сих пор, несмотря на все наши попытки сжечь все известные запасы ископаемого топлива, мы снизили концентрацию кислорода в атмосфере лишь на 0,001 %1.
Но этот огромный запас захороненного органического углерода не формируется постоянно: он откладывался на протяжении прошедших геологических эпох лишь периодически. В норме на Земле сохраняется положение, очень близкое к равновесию, при котором дыхание сводит на нет результаты фотосинтеза (а эрозия сводит на нет результаты отложения органики), так что в общем счете почти никакого захоронения углерода не происходит. Поэтому концентрация кислорода в атмосфере и остается на уровне примерно 21 % уже не один десяток миллионов лет. Но в редких случаях, в геологической древности, дела обстояли совсем по-другому. Наверное, самый впечатляющий пример — это каменноугольный период (карбон), около трехсот миллионов лет назад, когда по воздуху летали стрекозы размером с чайку, а по лесам шныряли многоножки длиною в метр. Эти гиганты были обязаны своим существованием исключительно высокой скорости происходившего в то время отложения углерода (так сформировались огромные запасы каменного угля, благодаря которым период и получил свое название — каменноугольный). За время отложения углерода на дне болот уровень кислорода в атмосфере подскочил до 30 %, давая некоторым существам (а именно животным, поглощающим кислород не путем активной вентиляции легких, а за счет пассивной диффузии в пронизывающих тело трахеях или на поверхности кожи) возможность достигать размеров гораздо больших, чем обычно2.
Чем была обусловлена беспрецедентная скорость отложения углерода в каменноугольный период? Почти наверняка — целым набором случайных факторов: взаимное расположение континентов, влажный климат, наличие обширных затапливаемых равнин, а также (что, возможно, особенно важно) появление в ходе эволюции лигнина, благодаря чему возникли большие деревья и другие крепкие растения, способные заселять обширные площади суши. Лигнин, который грибам и бактериям трудно разлагать даже сегодня, вскоре после своего появления был, судя по всему, и вовсе никому не по зубам. Поэтому его никто и не разлагал, добывая энергию, и он в огромных количествах оставался нетронутым, в то время как выделенный при его синтезе кислород насыщал атмосферу.
В истории Земли было и два других эпизода, когда сочетание случайных геологических факторов приводило к существенному повышению уровня кислорода в атмосфере. Оба эти эпизода были, возможно, связаны с глобальными оледенениями (предполагаемыми так называемой гипотезой “Земли-снежка”). Первое сильное повышение уровня кислорода в земной атмосфере произошло около 2,2 миллиарда лет назад, сразу вслед за периодом бурных геологических сдвигов и глобального оледенения. Второй период глобального оледенения (примерно с восьмисот до шестисот миллионов лет назад), судя по всему, тоже привел к повышению уровня кислорода. Эти глобальные бедствия, по-видимому, сказывались на равновесии фотосинтеза и дыхания, а также отложения осадочных пород и эрозии. Когда великие ледники таяли и начинали лить дожди, минералы и питательные вещества (железо, нитраты и фосфаты), соскобленные льдом с горных пород, смывались в океан, где вызывали бурное “цветение” водорослей и фотосинтетических бактерий, подобное “цветению воды”, к которому сегодня приводит использование удобрений, но, вероятно, гораздо большее по масштабам. Этот смыв должен был не только вызывать “цветение” фотосинтезирующих организмов, но и способствовать их захоронению: пыль, грязь и песок, попадавшие в океан, смешивались с “цветущими” бактериями и оседали на дне, приводя к беспрецедентному отложению углерода. А это, в свою очередь, приводило к тому, что уровень насыщенности атмосферы кислородом в масштабе планеты возрастал всерьез и надолго.
Итак, судя по всему, становление кислородной атмосферы на нашей планете было во многом случайным. Это впечатление лишь усилится, если принять во внимание отсутствие каких-либо изменений на протяжении других, весьма продолжительных периодов. С двух миллиардов примерно до одного миллиарда лет назад (в период, который геологи называют “скучным миллиардом” лет) на Земле, похоже, не произошло почти ничего примечательного. Концентрация кислорода в течение этого периода оставалась постоянной и довольно низкой, как, впрочем, бывало и в другие периоды, длившиеся сотни миллионов лет. Застой был нормой, но эпизоды геологических пертурбаций порой вносили серьезные изменения. Подобные геологические факторы могут работать и на других планетах, но, судя по всему, для совпадения обстоятельств, необходимого для накопления кислорода, требуется движение литосферных плит и вулканическая активность. Предположение, что фотосинтез мог давным-давно возникнуть на Марсе, не выходит за рамки возможного, но эта небольшая планета с ее угасающей вулканической активностью не могла поддерживать геологические потоки, которые позволили бы кислороду накапливаться. Поэтому если фотосинтез и мог на ней возникнуть, впоследствии он должен был повсеместно прекратиться.
Но есть и вторая, еще более важная причина, почему фотосинтез не обязательно должен приводить к формированию на планете кислородной атмосферы. Сам фотосинтез мог вообще не прийти к использованию воды в качестве сырья. Травы, деревья, водоросли — все они фотосинтезируют принципиально одинаково, выделяя кислород. Этот процесс называют кислородным фотосинтезом. Но если мы отступим на несколько шагов назад, к бактериям, то окажется, что есть и другие опции. Некоторые сравнительно примитивные бактерии используют для фотосинтеза не воду, а растворенное железо или сероводород. Если нам кажется, что такое сырье не годится для фотосинтеза, то только оттого, что мы привыкли к своему кислородному миру (продукту кислородного фотосинтеза) настолько, что нам трудно представить себе условия, которые были на Земле в древнейшие времена, когда фотосинтез впервые возник.
Нам также трудно оценить суть парадоксального, но на самом деле простого механизма фотосинтеза. Приведу пример, который, как я подозреваю (возможно, несправедливо), отражает общепринятое понимание фотосинтеза. Это отрывок из очаровательной книги Примо Леви “Периодическая система", опубликованной в 1975 году и объявленной “лучшей научно-популярной книгой всех времен” по результатам голосования, проводившегося в 2006 году среди публики (я тоже принял участие) в лондонском Королевском институте:
Углерод проникает внутрь листа, сталкиваясь там с множеством других (в данном случае бесполезных) молекул углерода и кислорода, присоединяется к большой и сложной молекуле, благодаря которой активизируется и одновременно получает важнейшее послание небес в виде сверкающего пучка солнечных лучей, после чего в одно мгновенье, как насекомое в лапах паука, лишается кислорода и соединяется с водородом, а также (есть такая версия) с фосфором, включаясь в цепь жизни (не важно, длинную или короткую).
Заметили ошибку? На самом деле их даже две, и очень жаль, что Леви их допустил, ведь в химическом механизме фотосинтеза по-настоящему удалось разобраться лет за сорок до сочинения его книги. “Сверкающий пучок солнечных лучей” не активирует молекулу углекислого газа: ее с тем же успехом можно активировать и среди ночи. Более того, она и не может активироваться светом, даже на самом что ни на есть солнцепеке. Кроме того, углерод не лишается кислорода в одно мгновение: кислород упорно отказывается расставаться с углеродом. Леви в своем рассказе исходит из распространенного, но ошибочного представления, будто кислород, выделяемый при фотосинтезе, берется из углекислого газа. Но это не так. Он происходит из воды, а это совсем другое дело. Осознать это — значит сделать первый шаг на пути к пониманию истоков фотосинтеза.
Кроме того, это значит сделать первый шаг на пути к решению энергетических и климатических проблем человечества.
Пучки солнечной энергии, используемые при фотосинтезе, разбивают молекулы воды на водород и кислород. Это та же реакция, которая идет в масштабах всей планеты, когда океан теряет под ударами ультрафиолета улетающий в космос водород. Чего удалось добиться фотосинтезирующим организмам (и чего до сих пор не можем добиться мы сами) — это получить катализатор, способный отнимать водород от молекул воды с минимальными затратами энергии, за счет мягкого видимого света, а не обжигающего ультрафиолетового. До сих пор все попытки людей изобрести нечто подобное заканчивались тем, что на “раскалывание” молекул воды тратилось больше энергии, чем ее можно было получить. Когда нам удастся повторить достижение фотосинтезирующих организмов, создав простой катализатор, аккуратно выбивающий из воды атомы водорода, энергетический кризис уже не будет нам страшен. Мы станем жечь водород, удовлетворяя все свои потребности в энергии, а отходами его горения будет чистая вода: никакого загрязнения, никакого углеродистого следа, никакого глобального потепления. Но это отнюдь не простая задача, поскольку молекула воды — поразительно стойкая комбинация атомов. Это видно по земным океанам: даже во время страшнейших штормов, когда волны без устали бьются о скалы, вода не распадается на кислород и водород. Вода — одновременно и самое распространенное, и самое труднодоступное сырье на нашей планете. Современному мореплавателю может прийтись по душе идея использовать в качестве топлива воду и солнечный свет. Но как это сделать? Об этом можно спросить у скоплений сине-зеленых “водорослей”, плывущих по волнам.
Стой же самой проблемой столкнулись в свое время и давние предки этих “водорослей” — современных цианобактерий. Они были единственной на нашей планете формой жизни, которой повезло найти способ расщеплять воду. Причем, как ни странно, цианобактерии стали расщеплять воду потому же, почему другие бактерии расщепляют сероводород или окисляют железо: им были нужны электроны. На первый взгляд, добывать электроны из воды — последнее дело. Но это не так.
По существу идея фотосинтеза проста: главное — электроны. Стоит добавить к углекислому газу несколько электронов, плюс несколько протонов, чтобы уравновесить заряды, и мы как по волшебству получим сахар. Сахара (углеводы) — одна из разновидностей органических веществ. Это и есть те цепи жизни, о которых писал Примо Леви. Именно из них в итоге и получается вся наша пища. Но откуда брать электроны? Если использовать немного солнечной энергии, их можно брать почти везде. В случае знакомого нам кислородного фотосинтеза они берутся из воды, но все же гораздо легче отнимать их у других соединений, не столь стойких, как вода. Если отнять электроны у сероводорода, то вместо кислорода, выделяемого в атмосферу, мы получим выпадающую в осадок серу — библейский жупел. Если отнять их у железа, растворенного в океане в виде заряженных частиц — ионов двухвалентного железа, — мы получим ржаво-рыжие соединения трехвалентного железа, выпадающие в осадок, образуя минералы. Возможно, именно так в свое время образовывались железистые кварциты, залегающие по всему миру и составляющие крупнейшие сохранившиеся запасы низкокачественной железной руды.
В современном, богатом кислородом мире эти формы фотосинтеза встречаются редко — просто потому, что необходимое для них сырье (сероводород и ионы двухвалентного железа) трудно найти в освещенной солнцем, хорошо аэрируемой воде. Но когда Земля была еще молода и свободного кислорода на ней почти не было, именно у этих веществ легче всего было брать электроны, и океаны были прямо-таки насыщены ими. Отсюда следует парадокс, решение которого имеет принципиальное значение для понимания корней фотосинтеза. Зачем переключаться с богатого и удобного источника электронов на нечто гораздо менее удобное — на воду, при расщеплении которой к тому же получаются отходы в виде кислорода — токсичного газа, который мог причинять серьезный вред клеткам любых выделявших его бактерий? Дело не может быть в том, что воды все равно еще больше, чем двух других веществ. Ведь для ее использования требуется не только энергия солнца, но и очень эффективный катализатор, а у эволюции нет дара предвидения. Неважно здесь и то, что кислородный фотосинтез преобразил нашу планету: планете это безразлично. Так что же за давление среды или что за мутации могли привести к такому новшеству?
Лежащий на поверхности ответ, который можно найти в учебниках, состоит в том, что запасы сырья истощились: жизнь обратилась к воде, потому что других подходящих альтернатив не осталось, точно так же, как нам, возможно, предстоит обратиться к воде, когда закончится ископаемое топливо. Но этот ответ не может быть правильным: из геологической летописи явственно следует, что кислородный фотосинтез возник задолго до того, как запасы другого сырья истощились, — более чем за миллиард лет. Жизнь вовсе не была загнана в угол.
Другой ответ, который начал вырисовываться лишь недавно, скрыт в механизме самого фотосинтеза, и этот ответ гораздо красивее. Он объединяет случай с необходимостью и проливает свет простоты на одну из самых запутанных и сложных проблем происхождения.
У растений добыча электронов осуществляется в хлоропластах. Это миниатюрные зеленые структуры, которые можно найти в клетках любого листа или травинки и которые своим цветом и делают листья и траву зелеными. Их название связано с названием пигмента, который, в свою очередь, делает зелеными сами хлоропласты. Этот пигмент — хлорофилл. Он участвует в фотосинтезе, отвечая за поглощение солнечной энергии. Молекулы хлорофилла встроены в причудливую систему мембран, расположенную внутри каждого хлоропласта. Эти мембраны образуют большие стопки уплощенных дисков (ничто так не напоминающих, как инопланетную электростанцию из фантастического фильма), связанных друг с другом трубками, переброшенными через промежутки между дисками под разными углами и на разной высоте. В дисках и осуществляется великое дело фотосинтеза — извлечение электронов из воды.
Как ни сложно добывать электроны из воды, растениям это удается. Участвующие в этом комплексы белков и пигментов так велики (по молекулярным меркам), что их размеры можно сравнить с размерами небольшого города. Все они объединены в два больших комплекса — так называемые фотосистемы I и II. Каждый хлоропласт содержит тысячи таких фотосистем. Их задача состоит в том, чтобы улавливать лучи света и синтезировать с их помощью органические вещества. На то, чтобы разобраться в механизме их работы, потребовалось больше половины столетия и целый ряд экспериментов из числа самых красивых и остроумных в истории науки. Здесь, к сожалению, не место их обсуждать3. Сейчас нам нужно сосредоточиться на том, что в итоге удалось узнать и что это может сказать нам об изобретении фотосинтеза.
Сердце механизма фотосинтеза называют Z-схемой. Она в равной степени восхищает и приводит в ужас студентов, изучающих биохимию. Эту схему, которая представляет собой описание “энергетического профиля” фотосинтеза, впервые нарисовал блистательный и при этом скромный английский ученый Робин Хилл в 1960 году. Хилл не особенно распространялся о своей работе. Сотрудники его лаборатории не хотели донимать его расспросами, и поэтому даже они были удивлены, когда в одном из номеров журнала “Нейчур” за 1960 год была опубликована его гипотеза: они плохо представляли себе, над чем именно он трудился. На самом деле открытие Z-схемы было основано не на результатах собственных экспериментов Хилла (хотя они тоже были исключительно важны), а на других, уже известных и довольно загадочных экспериментальных наблюдениях. Главным из них был один интересный факт из области термодинамики. Как выяснилось, в число продуктов фотосинтеза входят не только новые молекулы органических веществ, но и АТФ — “энергетическая валюта” жизни. Удивительно, но выход АТФ оказался, судя по всему, строго сопряжен с выходом органики: чем больше органических веществ получалось в результате фотосинтеза, тем больше получалось АТФ (и наоборот: если органики синтезировалось меньше, то меньше синтезировалось и АТФ). Солнце как будто кормило растения двумя бесплатными обедами одновременно. Замечательно, что Робин Хилл, отталкиваясь от одного этого факта, сумел угадать суть всего механизма фотосинтеза. Говорят, что гений - это способность прежде всех узреть очевидное4.
И все же (что весьма характерно для Хилла) предложенный им термин “Z-схема”, при всей лаконичности, может ввести в заблуждение. Букву I на самом деле следовало бы повернуть на 90°, чтобы получилась буква Л/, которая более точно отражала бы энергетический профиль фотосинтеза. Представьте себе левую палочку буквы N как вертикально восходящую реакцию, для осуществления которой требуется приток энергии. Косая черта, идущая вниз, будет, таким образом, означать нисходящую реакцию, приводящую к выделению энергии, которую можно улавливать и запасать в форме АТФ. Наконец, правая палочка означает вторую восходящую реакцию, требующую затрат энергии.
Две участвующие в фотосинтезе системы, фотосистема I и фотосистема II, располагаются в двух нижних точках буквы N. Фотон света падает на первую фотосистему и забрасывает электрон на более высокий энергетический уровень, после чего энергия электрона поэтапно снижается вновь, проходя несколько небольших молекулярных стадий, в ходе которых она используется для синтеза АТФ. Спустившись на низкий энергетический уровень, электрон попадает во вторую фотосистему, где еще один фотон снова забрасывает его на более высокий энергетический уровень. С этого уровня электрон наконец передается на молекулу углекислого газа, с чего и начинается синтез молекулы сахара. Разобраться в этом процессе помогает рисунок Ричарда Уокера, на котором процесс изображен в виде трюка ярмарочных силачей, один из которых бьет по планке молотом, забрасывая металлический шар на вершину высокого помоста, В данном случае удар молота дает шару энергию, необходимую, чтобы забросить его наверх, а в случае фотосинтеза энергия фотона солнечного света делает то же самое с электроном.
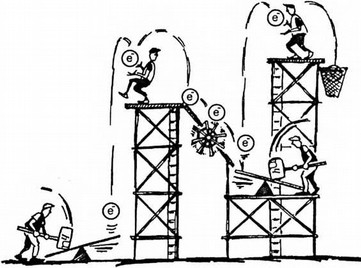
Рисунок Ричарда Уокера, изображающий принцип работы Z-схемы. Энергия фотона (показана в виде удара молота) забрасывает электрон на более высокий энергетический уровень. Часть энергии, выделяемой во время обратного спуска этого электрона, используется для осуществления различных клеточных процессов. Затем второй фотон забрасывает электрон на еще более высокий энергетический уровень. Там его ловит высокоэнергетическая молекула (НАДФ), которая затем реагирует с молекулой углекислого газа, образуя молекулу органического вещества.
Z-схема (или, если угодно, N-схема) кажется на удивление запутанным механизмом, но на то есть весомые технические причины. Совместить отнятие электронов у воды с преобразованием углекислого газа в сахар каким-либо иным способом почти невозможно в силу химических ограничений. Это связано с природой явления передачи электронов, а именно — с химическим сродством электронов с различными соединениями. Молекулы воды, как мы убедились, очень стойки: они обладают высоким сродством к своим электронам. Чтобы отнять электрон у воды, нужно потянуть с огромной силой. Иными словами, нужен очень мощный окислитель. Этим мощным окислителем служит жадная форма хлорофилла, своего рода молекулярный мистер Хайд, возникающий из кроткого доктора Джекилла при поглощении высокоэнергетических фотонов5. Но что хорошо тянет, то обычно не столь же хорошо толкает. Вещество, крепко хватающее электроны, в силу химических причин неохотно их отдает, точно так же, как человеконенавистник мистер Хайд или какой-нибудь алчный скряга не будет склонен к внезапным приступам щедрости, заставляющим его делиться своими богатствами. Точно так же обстоят дела и с этой формой хлорофилла. Активированная светом, она приобретает колоссальную силу, позволяющую ей оттягивать электроны от воды, но ей очень трудно после этого оттолкнуть их от себя и передать дальше. Говоря научным языком, это мощный окислитель, но слабый восстановитель.
С углекислым газом тоже проблема. Его молекулы тоже очень стойки и не имеют химического желания, чтобы их пичкали новыми электронами. Они готовы принимать электроны, и то неохотно, лишь от могучих толкателей, то есть, по-научному, сильных восстановителей. А для этого требуется другая форма хлорофилла, которая очень хорошо умеет толкать, но плохо умеет тянуть. Это уже не алчный скряга, а скорее навязчивый уличный торговец, старающийся всеми правдами и неправдами “толкнуть” свой сомнительный товар. Активированная светом молекула этой формы хлорофилла может “сплавлять” электроны своему партнеру в этой сомнительной торговле — молекуле НАДФ, которая тоже всячески стремится от них избавиться, в итоге передавая их молекуле углекислого газа6.
Так что на использование в фотосинтезе двух фотосистем есть особые причины. Тут нет ничего особенно удивительного. Гораздо сложнее ответить на вопрос о том, как эти сложные взаимосвязанные системы могли возникнуть в ходе эволюции. На самом деле в процессе фотосинтеза задействованы даже не две, а пять частей. Первая — это кислород-выделяющий комплекс, своего рода молекулярная “орехоколка”, располагающая молекулы воды так, чтобы раскалывать их одну за другой, отнимая электроны и выделяя в качестве отходов кислород. Затем идет фотосистема II (в названиях здесь получилась некоторая путаница: в силу исторических причин две фотосистемы были пронумерованы в обратном порядке), которая, если активировать ее светом, превращается в молекулярного мистера Хайда и отбирает эти электроны у кислород-выделяющего комплекса. Затем идет электрон-транспортная цепь, по которой электроны передаются дальше, как мяч, передаваемый через поле игроками в регби. Электрон-транспортная цепь использует энергию, выделяемую при движении электронов вниз по энергетическому профилю, чтобы синтезировать немного АТФ, а затем передает те же электроны фотосистеме I. Еще один фотон снова забрасывает электроны на высокий энергетический уровень, где их прибирает к рукам “навязчивый торговец” — молекула НАДФ, сильный толкатель электронов, которому только и нужно снова от них избавиться. А затем, наконец, идет молекулярный аппарат, необходимый для активации углекислого газа и преобразования его в сахар. Работа “навязчивого торговца”, запущенного фотосистемой I и преобразующего углекислый газ в сахар, идет за счет химической, а не световой энергии, за что ее и называют темновой фазой фотосинтеза. Эту особенность Примо Леви совсем упустил из виду.
Все пять систем работают последовательно, отнимая электроны у воды и навязывая их углекислому газу. Это необычайно сложный способ “колоть орехи”, но, судя по всему, данный “орех” только так и можно расколоть. Большой эволюционный вопрос состоит в том, как все эти сложные и взаимосвязанные системы могли возникнуть и организоваться едва ли не единственно возможным способом, который делает возможным кислородный фотосинтез.
Слово “факт” нередко вызывает у биологов содрогание, потому что из всякого правила в биологии может быть множество исключений. Но один факт, касающийся кислородного фотосинтеза, почти точно соответствует действительности: кислородный фотосинтез возник в ходе эволюции лишь однажды. Хлоропласт — структуру, в которой происходит фотосинтез, — можно найти во всех фотосинтезирующих клетках всех растений и водорослей. Хлоропласты вездесущи и явно родственны друг другу. Все они хранят одну общую историческую тайну. Ключ к их прошлому кроется в их размерах и в форме: они похожи на бактерий, живущих внутри более крупной клетки-хозяина. Дело в том, что они и вправду произошли от бактерий, о чем свидетельствует также присутствие во всех хлоропластах собственных кольцевых молекул ДНК. Эти молекулы реплицируются перед каждым делением хлоропластов и передаются дочерним хлоропластам точно так же, как при делении бактерий подобные молекулы передаются дочерним бактериальным клеткам. Последовательности букв в ДНК хлоропластов не только подтверждают родство хлоропластов с бактериями, но и выдают их ближайших современных родственников — цианобактерий. И последнее, не менее важное обстоятельство: Z-схема фотосинтеза растений со всеми пятью ее составляющими, хотя и в более простом виде, свойственна и цианобактериям. Короче говоря, нет никаких сомнений в том, что предками хлоропластов были именно свободноживущие цианобактерии.

Типичный хлоропласт (из листа свеклы Beta vulgaris) в разрезе. Видны стопки мембранозных дисков (тилакоидов), где в ходе фотосинтеза расщепляется вода и выделяется кислород. Сходство с бактерией не случайно: предками хлоропластов были свободноживущие цианобактерии.
Цианобактерии, которых раньше называли поэтичным, но некорректным термином “сине-зеленые водоросли”, составляют единственную известную группу бактерий, представители которой способны расщеплять молекулы воды посредством кислородной формы фотосинтеза. Как именно некоторые из их числа поселились в более крупных клетках-хозяевах — тайна геологической древности, покрытая мраком. Несомненно, что это произошло более миллиарда лет назад. Предполагается, что однажды эти цианобактерии были поглощены, но не переварены (что бывает не так уж редко) и в итоге оказались иначе полезны клетке-хозяину. Потомки таких клеток впоследствии основали два великих царства, водорослей и растений, определяющей чертой которых служит способность всех их представителей жить на солнце и в воде благодаря фотосинтетическому аппарату, унаследованному от бактерий, некогда поселившихся внутри их предков.
Так что вопрос о происхождении фотосинтеза сводится к вопросу о происхождении цианобактерий — единственной разновидности бактерий, представителям которой удалось решить проблему расщепления воды. А это один из самых спорных и, более того, по-прежнему неразрешенных вопросов биологии.
До самого конца прошлого тысячелетия большинству исследователей представлялись убедительными, хотя и досадными, выводы, основанные на замечательных открытиях, которые сделал Билл Шопф, энергичный профессор палеобиологии из Калифорнийского университета в Лос-Анджелесе. С 8о-х годов Шопф нашел и изучил целый ряд древнейших ископаемых остатков жизни на Земле — возрастом около трех с половиной миллиардов лет. Нужно пояснить, что здесь имеется в виду под ископаемыми остатками. Шопф нашел цепочки микроскопических минеральных капсул, очень похожих на бактерий и примерно соответствующих им по размерам. Изучив их тонкую структуру, Шопф первоначально объявил их ископаемыми цианобактериями. Эти крошечные микроокаменелости были часто связаны с чем-то похожим на ископаемые строматолиты. Живые строматолиты представляют собой минерализующиеся столбики, постепенно нарастающие слой за слоем и достигающие в высоту около метра; их образуют процветающие сообщества бактерий, покрывающих в виде налета откладываемые под ними минеральные слои. Наружные, живые слои современных строматолитов обычно кишат цианобактериями. Это позволило Шопфу утверждать, что обнаруженные им ископаемые формы могут служить дополнительным свидетельством древности происхождения цианобактерий. Предвидя возможные возражения, Шопф также показал, что эти предполагаемые ископаемые содержат остатки органического углерода, состав которого, судя по всему, указывал на его происхождение из живых организмов, причем не только древних, но и фотосинтети- ческих. В общем, по словам Шопфа, цианобактерии или некие очень похожие на них существа уже появились три с половиной миллиона лет назад — всего через несколько сотен миллионов лет после окончания великого обстрела астероидами, которым были отмечены самые первые годы нашей планеты, образовавшейся вскоре после возникновения всей Солнечной системы.

Живые строматолиты в лагуне Хамелин-Пул в заливе Шарк-Бей на западе Австралии. Вода в лагуне примерно вдвое солонее, чем в открытом океане. Это создает условия, непригодные для существования животных, питающихся бактериальными пленками, таких как брюхоногие моллюски, и дает колониям цианобактерий возможность процветать.
Мало кто обладал достаточной квалификацией, чтобы оспорить предложенную Шопфом интерпретацию этих древних ископаемых, но тех немногих, кто обладал, судя по всему, его аргументы убедили. Однако некоторые другие исследователи, может, не столь квалифицированные, были настроены более скептически. Не так-то просто было совместить концепцию раннего появления цианобактерий (которые предположительно должны были выделять в виде отходов кислород, как это делают современные цианобактерии) с появлением первых геологических признаков кислорода в атмосфере, отмечаемым гораздо больше миллиарда лет спустя. Может, еще важнее было то, что сложность Z-схемы вызывала у большинства биологов недовольство идеей, что кислородный фотосинтез мог возникнуть в ходе эволюции так быстро. Им представлялось, что по-настоящему древними скорее могли быть другие, более простые формы фотосинтеза. Так что большинство ученых в целом согласилось с тем, что исследованные Шопфом ископаемые были бактериями (может быть, фотосинтетическими), однако по-прежнему сомневалось, что это были действительно цианобактерии — главный шедевр бактериальной жизни.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Фотосинтез без хлорофилла
Фотосинтез без хлорофилла Биофизик Ю. Владимиров рассказал мне однажды, что лет двадцать назад академик А. Красновский спросил как-то своих учеников: — Какой самый простой признак фотосинтеза? — Присутствие хлорофилла, — дружно ответили его молодые коллеги. Догма о
ГЛАВА I
ГЛАВА I Задачи обработки земли.Обработка земли преследует две главные, на первый взгляд противоположные, цели: во-первых, она должна стремиться создать такое соотношение благоприятствующих росту растений условий, при котором можно было бы получить самую богатую
ГЛАВА XII.
ГЛАВА XII. Очистка почвы от сорных трав.Целью очистки почвы от сорных трав является такое ее состояние, при котором после посева не могло бы расти никакое другое растение, кроме посеянного. Если мы позволим сорным травам расти на ниве, то они будут отнимать у почвы
ГЛАВА XIV.
ГЛАВА XIV. Обработка земли под озимь.Весною 1898 г. посев начался у меня очень поздно — 21 марта по старому стилю. Была очень сухая весна, и нетрудно было предвидеть, что почва ссохнется, как кирпич, так что мелко вспахать землю под озимь будет очень трудно.Поэтому тотчас же
ГЛАВА XV.
ГЛАВА XV. Обработка земли под яровые хлеба.Обработку земли под яровые хлеба я начинаю тотчас после уборки озими . Только при соблюдении этого условия можно рассчитывать на самый обильный урожай.Поля, поросшие сорными травами и покрытые густым жнивьем, я вспахиваю
ГЛАВА XVI.
ГЛАВА XVI. Посев.Как нужно сеять, чтобы получить самый обильный урожай? Чтобы ответить на этот вопрос, нам нужно будет припомнить те условия, при которых растения развиваются в желательном для земледельца направлении. Эти условия, о которых мы говорили во второй главе,
Что такое фотосинтез и какое значение он имеет для жизни на Земле?
Что такое фотосинтез и какое значение он имеет для жизни на Земле? Фотосинтезом называют образование высшими растениями, водорослями, фотосинтезирующими бактериями сложных органических веществ, необходимых для жизнедеятельности как самих растений, так и всех других
2.4. Фотосинтез и хемосинтез
2.4. Фотосинтез и хемосинтез Основной путь, по которому практически вся энергия попадает в биосферу – это процесс фотосинтеза. Каждый год на Земле в процессе фотосинтеза образуется более 150 ? 109 тонн органических веществ. Вклад хемоавтотрофов (к ним относятся только
Бактериальный фотосинтез и хемосинтез
Бактериальный фотосинтез и хемосинтез Различные группы фотосинтезирующих бактерий осуществляют особую разновидность фотосинтеза – бактериальный фотосинтез. Бактерии имеют свои специфические фотосинтетические пигменты (в частности, различные бактериохлорофиллы),
Глава 2. ДНК
Глава 2. ДНК На стене паба “Орел” в Кембридже висит синяя мемориальная доска, установленная в 2003 году в честь пятидесятилетия одного случая, когда разговоры в этом пабе приняли не совсем обычный оборот. Во время обеда 28 февраля 1953 года два завсегдатая “Орла”, Джеймс
Глава 1. Вид
Глава 1. Вид ТЕМЫ• История эволюционных идей• Современное эволюционное учение• Происхождение жизни на Земле• Происхождение человекаВ настоящее время на нашей планете обитает несколько миллионов видов живых организмов, каждый из которых по своему уникален. Каким
17. Пластический обмен. Фотосинтез
17. Пластический обмен. Фотосинтез Вспомните!Какую часть метаболизма называют пластическим обменом?Какова роль зелёных растений в природе?В каких органоидах клетки осуществляется фотосинтез?Любой живой организм – открытая динамичная система, в которой постоянно