3. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА
3. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА
Строение мозга у животных разных видов разнообразно. И хотя, как следует из рис. 19.3, родственные виды, например среди ракообразных или млекопитающих, имеют общие черты в строении мозга, между собой они мало похожи. Именно с появлением нервной системы организмы получили возможность быстрой адаптации к быстро меняющимся факторам среды, что, безусловно, давало преимущества в борьбе за существование. У предков млекопитающих, как и у современных рептилий, кора больших полушарий была очень слабо дифференцирована [Батуев, 1979]. Но на пути от рептилиеподобных предков млекопитающих до ныне живущих млекопитающих произошло значительное увеличение коры мозга по сравнению с другими структурами мозга (рис. 19.4), изменение количества (см. рис. 19.3 В), размеров и формы корковых областей, их модульного строения и связей [Kaas, 1987], произошло увеличение переднего мозга и, как предполагается [McLean, 1993; Povinelli, Preuss, 1995], перемещение главенствующей роли от стриарного комплекса сначала к лимбической системе, а затем к неокортексу и особенно к префронтальной области коры (см. рис. 19.3 Б).
Важной чертой эволюции млекопитающих является уже упомянутое увеличение неокортекса, при этом степень этого увеличения отличает приматов от остальных млекопитающих, а человека – от остальных приматов. В частности, соотношение площади неокортекса у мыши, макаки и человека составляет 1 : 100 : 1000 соответственно [цит. по: Rakic, 1995], а соотношение объёмов коры мозга и спинного мозга у крыс и у человека – 31 : 35 и 77 : 2, соответственно (см. рис. 19.4) [Swanson, 1995]. Такая экспансия неокортекса, как полагают, связана с модификацией регуляторных генов, определяющих митотическое деление клеток вентрикулярного (пролиферативного) слоя коры [Rakic, 1995]. При этом, по-видимому, кора развивалась под влиянием внутрикорковых или непрямых таламических подкорковых влияний [Krubitzer, 1995]. Такое развитие коры привело к возникновению новых мощных приспособительных функций [Innocenti, Kaas, 1995]. Можно предположить, что именно это явилось основой такого свойства нервной системы, которое У. Матурана обозначил как способность нервной системы «взаимодействовать со своими собственными внутренними состояниями, как если бы те были независимыми сущностями» [Матурана, 1995, с. 104]. В таком «взаимодействии» нервной системы со своими собственными состояниями может лежать ключ и к пониманию онтологической сути субъективных состояний – «переживаний», «мыслей», «образов», – а также языка и мышления.
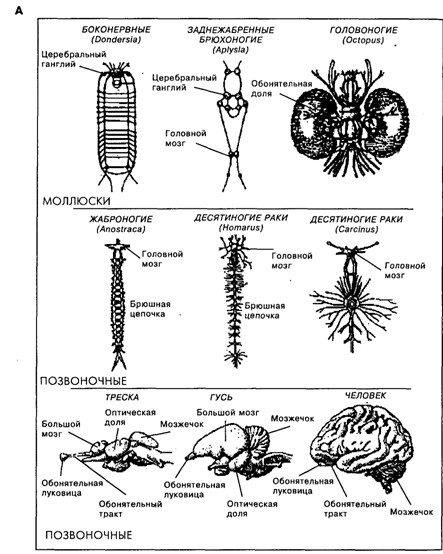
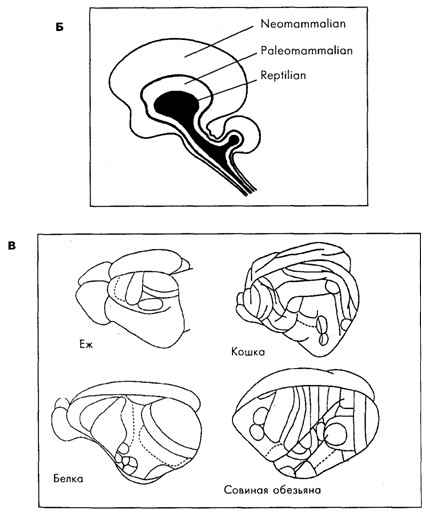
Рис. 19.3. А. Схематичное представление мозга животных, относящихся к линиям моллюсков, членистоногих и хордовых (масштабы разные)[по Кэндел, 1980]
Б. Условное представление триединой эволюции трёх основных мозговых ансамблей, приведших к образованияю переднего мозга у людей и у других развитых млекопитающих [McLean, 1993].
В . Корковые поля у примитивных млекопитающих (ёж), у млекопитающих с незначительно более развитой нервной системой (белка) и у двух млекопитающих с умеренно развитым мозгом (кошка и совиная обезьяна) [Kaas, 1987]

Рис. 19.4. Сравнение размеров разных отделов цетральной нервной системы крысы и человека. Пропорции между областями мозга соблюдены [Swonson, 1995]
Внутрикорковые процессы рассматриваются рядом авторов в качестве нейронной основы сознания [Маунткасл, 1981; Эделмен, 1981; Eccles, 1992]. Так, например, в концепциях В. Маунткасла и Дж. Эделмена сознание возникает на основе повторного входа «сигнализации» (см. гл. 11), при этом, согласно одним взглядам, данный процесс происходит в модульных распределённых сетях [Маунткасл, 1981], а согласно другим – в вырожденной группе нейронов [Эделмен, 1981]. По мнению же Дж. Экклса, сознание – побочный эффект, возникший в эволюции неокортекса, наблюдающийся только у млекопитающих и сводящийся к взаимодействию мозга и ментального мира в множестве пресинаптических окончаний корковых нейронов, которые образуют морфологическую единицу – дендрон [Eccles, 1992].
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.3. Экспериментальные эволюционные машины (ЭЭМ)
6.3. Экспериментальные эволюционные машины (ЭЭМ) В предыдущем параграфе мы упоминали о том, что непрерывный рост микробных популяций в проточных системах позволяет экспериментально изучать микроэволюционные переходы, т.е. шаги эволюции.Суть непрерывного процесса
КОГДА-ТО НАШИ ЭВОЛЮЦИОННЫЕ ПУТИ РАЗОШЛИСЬ
КОГДА-ТО НАШИ ЭВОЛЮЦИОННЫЕ ПУТИ РАЗОШЛИСЬ В области сексуальных и брачных отношений человекообразные обезьяны явно ушли от общих с человеком предков своими особыми путями. И общее у них здесь с человеком — это наличие менструальных циклов, заменивших характерную для
Эволюционные истоки морфологических особенностей человека
Эволюционные истоки морфологических особенностей человека Как с эволюционной точки зрения интерпретировать переход к прямохождению, исчезновение волос на теле, нетипичное для обезьян распределение жировой ткани, стремительное увеличение размеров головного мозга и
Эволюционные стратегии популяций
Эволюционные стратегии популяций В эволюционной экологии рассматриваются различные теоретические обобщения, показывающие межвидовые взаимоотношения. В результате взаимодействия у животных выработались собственные эволюционные стратегии. Одной из важнейших
Разрывы, недостающие звенья и эволюционные механизмы
Разрывы, недостающие звенья и эволюционные механизмы Отсутствие предковых форм или форм, промежуточных между отдельными ископаемыми видами, не является некой странной особенностью ранней истории Metazoa. Разрывы представляют собой общее явление и встречаются по всей
Эволюционные изменения в организации яиц со спиральным дроблением
Эволюционные изменения в организации яиц со спиральным дроблением Для многих зародышей со спиральным дроблением, в особенности для зародышей моллюсков и кольчецов, составлены очень точные карты, на которых указана дальнейшая судьба различных клеток. Разные типы
Эволюционные изменения в организации яиц хордовых
Эволюционные изменения в организации яиц хордовых Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от
1. ЕСТЕСТВЕННАЯ АГРЕССИВНОСТЬ И ЭВОЛЮЦИОННЫЕ МЕХАНИЗМЫ, ЕЕ ОГРАНИЧИВАЮЩИЕ
1. ЕСТЕСТВЕННАЯ АГРЕССИВНОСТЬ И ЭВОЛЮЦИОННЫЕ МЕХАНИЗМЫ, ЕЕ ОГРАНИЧИВАЮЩИЕ Три закона робототехники1. Робот не может причинить вред человеку или своим бездействием допустить, чтобы человеку был причинен вред.2. Робот должен повиноваться всем приказам, которые отдает
1.3. Существование самоотверженности и ее эволюционные преимущества
1.3. Существование самоотверженности и ее эволюционные преимущества Логике примитивного социал-дарвинизма противоречат факты массового героизма и самоотвержения, с существованием героической верности долгу, с самоуничтожением ради выполнения долга, с существованием
Когда-то наши эволюционные пути разошлись
Когда-то наши эволюционные пути разошлись Человекообразные в области сексуальных и брачных отношений явно ушли от общих с человеком предков своими особыми путями. Общее с человеком у них только наличие менструальных циклов, заменивших овуляцию раз в год, характерную
Глава 6. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ. ЭВОЛЮЦИОННЫЕ И БИОЛОГИЧЕСКИЕ АСПЕКТЫ
Глава 6. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ. ЭВОЛЮЦИОННЫЕ И БИОЛОГИЧЕСКИЕ АСПЕКТЫ При рассмотрении эволюции пищеварения мы сталкиваемся с несколькими на первый взгляд исключающими друг друга закономерностями. В самом деле, в ходе эволюции механизмы, участвующие в
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера С деятельностью живых систем связана та часть поверхности Земли, которая объединена под названием биосферы. Биосфера, являющаяся самой крупной экосистемой, представляет собой
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера С деятельностью живых систем связана та часть поверхности Земли, которая объединена под названием биосферы. Биосфера, являющаяся самой крупной экосистемой, представляет собой
Эволюционные предпосылки гомосексуальности
Эволюционные предпосылки гомосексуальности Гомосексуальное поведение — явление отнюдь не сугубо человеческое. Оно широко распространено у животных в естественной среде обитания. Гомосексуальные отношения можно часто наблюдать в группах холостяков у павианов
Эволюционные корни ритуала
Эволюционные корни ритуала Антропологи уже давно пришли к выводу, что ритуалы в древнейших человеческих сообществах способствовали выживанию, поддерживая в данном клане или племени чувство особой идентичности и общей судьбы[91]. Ритуал постоянно напоминал членам