Глава 27. Регуляция и взаимосвязь метаболизма
Глава 27. Регуляция и взаимосвязь метаболизма
Для нормального функционирования организма должна осуществляться точная регуляция потока метаболитов по анаболическим и катаболическим путям. Все сопутствующие химические процессы должны протекать со скоростями, отвечающими требованиям организма как единого целого в условиях окружающей среды. Генерация АТФ, синтез макромолекул, транспорт, секреция, реабсорбция и другие процессы должны чутко реагировать на изменения в окружении, в котором находится клетка, орган или весь организм. Клеточный метаболизм основан на принципе максимальной экономии. Клетка потребляет в каждый данный момент как раз такое количество питательных веществ, которое позволяет ей удовлетворять свои энергетические нужды. Такая высокая организация и скоординированность метаболизма достигается с помощью регуляторных механизмов. Эти механизмы достаточно разнообразны.
Различают несколько уровней регуляции метаболизма:
1. Молекулярный.
2. Клеточный.
3. Органный (тканевой).
4. Организменный.
По времени достижения регуляторного эффекта различают быструю регуляцию (действующую в течение секунд и минут) и медленную регуляцию (в течение часов и суток).
Основными регуляторными механизмами являются:
1. Регуляция на уровне мембран.
2. Регуляция с участием циклических нуклеотидов и других вторичных посредников.
3. Регуляция количества ферментов.
4. Регуляция ферментативной активности.
5. Гормональная регуляция.
Регуляция на уровне мембран может осуществляться посредством нескольких механизмов. Во-первых, это избирательная проницаемость мембран для различных метаболитов и ионов. Во-вторых, способность мембран фиксировать гормоны с помощью рецепторов. В-третьих, ферментативная активность мембран. На уровне мембран реализуются, по крайней мере частично, такие регуляторные факторы, как доступность субстратов и коферментов, удаление продуктов реакции.
Циклические нуклеотиды и другие вторичные посредники участвуют в реализации действия целого ряда гормонов.
Регуляция количества ферментов.
Концентрация любого фермента определяется соотношением скоростей его синтеза и распада. Скорость синтеза белков-ферментов регулируется с помощью механизмов, общих для регуляции синтеза других белков. Влияние регуляторных факторов может интегрально проявляться в виде репрессии или индукции синтеза фермента. Данный механизм относится к медленному типу регуляции метаболизма.
Регуляция активности ферментов.
Это один из наиболее разнообразных методов регуляции метаболизма. Он может реализоваться по целому ряду механизмов, которые подробно изложены в главе 4.
Аллостерическая регуляция метаболических путей
Аллостерические регуляторы бывают, как правило, двух типов:
1. Конечные продукты цепей последовательных реакций, регулирующие свой синтез по принципу обратной связи.
2. АТФ, АДФ, АМФ, НАД+ и НАДН•+Н+. Эти соединения хотя и не являются конечными продуктами самих метаболических путей, но образуются в результате их протекания и оказывают регуляторное влияние на поточную скорость. АТФ служит активатором ферментов, действующих в направлении синтеза биополимеров и аккумуляции энергии и является ингибитором реакций катаболизма. АДФ, а иногда и АМФ играют обратную роль – они активируют пути катаболизма, обеспечивающие их превращение в АТФ, ингибируют процессы анаболизма, связанные с потреблением АТФ, в котором клетки испытывают недостаток. НАД+ в этом смысле ведет себя подобно АМФ, НАДН+Н+ выступает в том же качестве, что и АТФ.
Как правило, аллостерические ферменты занимают место в начале мультиферментной последовательности реакций и катализируют ту её стадию, которая лимитирует скорость всего процесса в целом. Обычно роль такой стадии играет практически необратимая реакция. В некоторых случаях аллостерический фермент одного метаболического пути специфическим образом реагирует на промежуточные или конечные продукты другого. Благодаря этому достигается необходимая координация различных метаболических путей, направленная на обеспечение конкретных функций или процессов. Например, при мышечном сокращении возрастает скорость утилизации АТФ, необходимой для его энергообеспечения. При этом компенсаторно увеличивается скорость гликолиза с помощью регуляторных механизмов в соответствии с понижением уровня АТФ. В результате активации гликолиза увеличивается скорость наработки ацетил-КоА, являющегося субстратом цикла трикарбоновых кислот. Активация цикла трикарбоновых кислот приводит к наработке повышенных количеств НАДН+Н+, который вовлекается в цепь тканевого дыхания, активность которой при этом также увеличивается. Это приводит к ресинтезу АТФ и пополнению её пула, сниженного в результате мышечного сокращения.
Взаимосвязь метаболизма
Метаболизм в целом не следует понимать как сумму обменов белков, нуклеиновых кислот, углеводов и липидов. В результате взаимодействия обменов отдельных классов органических соединений возникает единая система метаболических процессов, представляющая собой качественно новое образование. Обмены важнейших структурных мономеров живых систем – аминокислот, моносахаридов (глюкозы), жирных кислот, мононуклеотидов тесно взаимосвязан. Эта взаимосвязь осуществляется через так называемые ключевые метаболиты, которые служат общим звеном на путях распада или синтеза мономеров. Взаимосвязь обменов отдельных классов органических соединений особенно хорошо выражена в процессах их взаимного превращения, хотя и не сводится только к этому. Примером такого взаимопревращения может являться прирост массы тела за счет отложения подкожного жирового слоя при избыточном потреблении углеводной пищи. К ключевым метаболитам, которые служат узловыми моментами взаимосвязи метаболизма относятся пируват, глицерофосфат, ацетил-КоА, некоторые метаболиты цикла трикарбоновых кислот (Рис. 27.1).
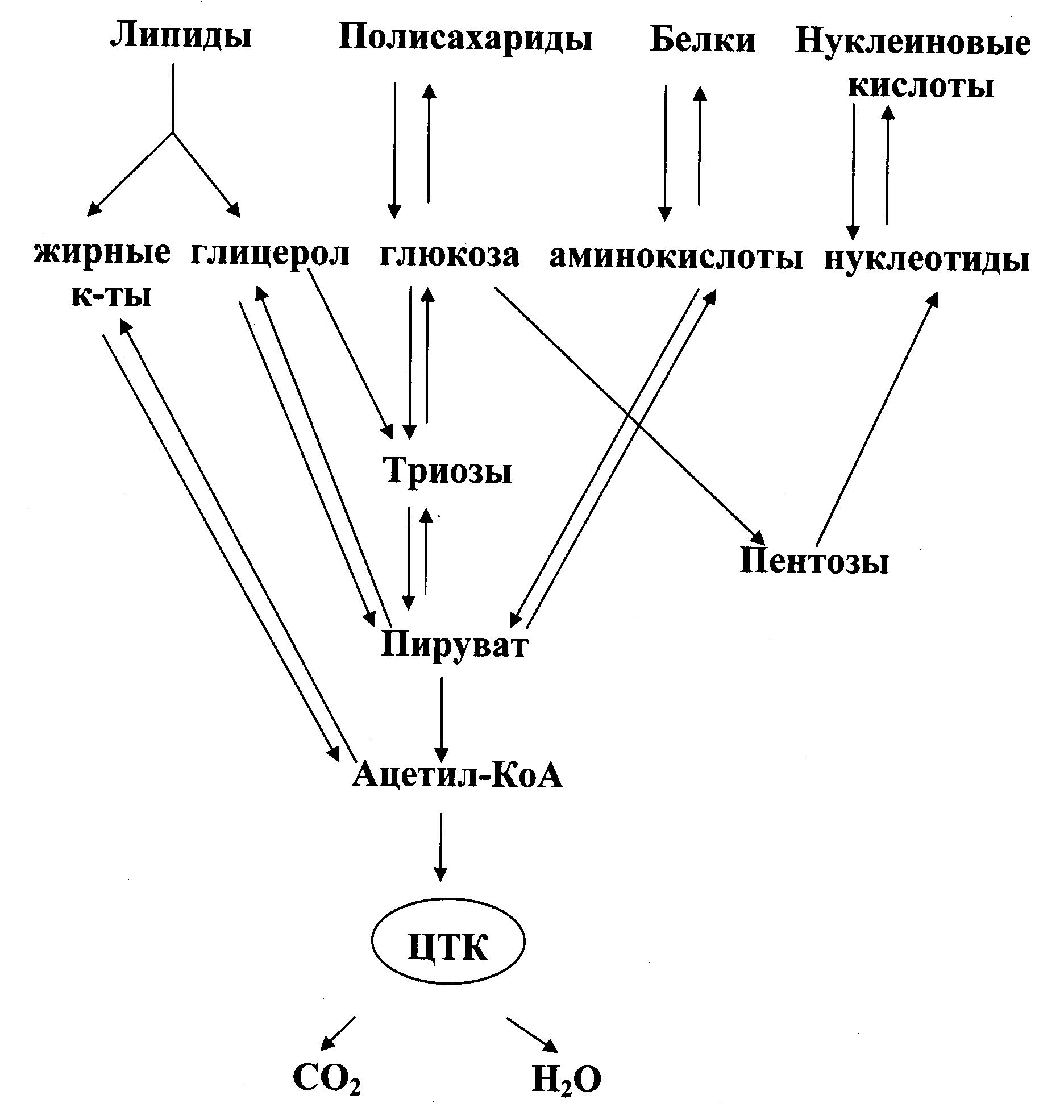
Рис. 27.1. Взаимосвязь метаболизма различных классов органических соединений.
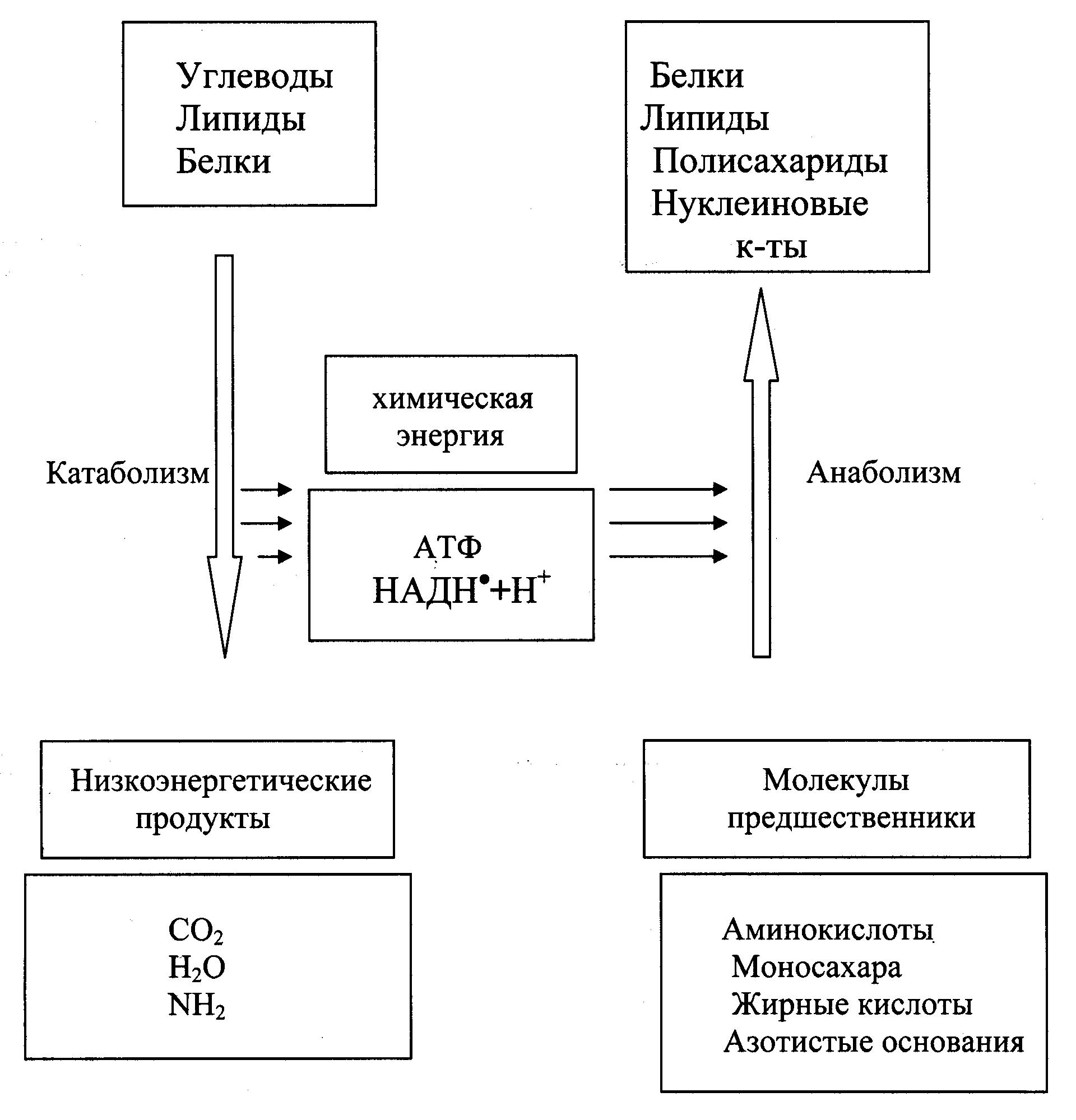
Рис. 27.2. Энергетические взаимосвязи между катаболическими и анаболическими путями.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава XIII Регуляция экспрессии генов
Глава XIII Регуляция экспрессии генов О проблеме регуляции экспрессии генов мы в этой книге говорим фактически во всех главах, рассматривая ее с разных сторон. Существует такое, может быть несколько одностороннее, определение развития: «Понять развитие — это значит
Взаимосвязь наук
Взаимосвязь наук «Недопустимо, чтобы молодые ученые забывали о том, что новые открытия обнаруживают тенденцию возникать в пограничной зоне между различными науками, где одна дисциплина примыкает к другой. Если бы я понимал это раньше, я был бы гораздо лучшим
7. Виды метаболизма бактерий
7. Виды метаболизма бактерий В процессе метаболизма выделяют два вида обмена:1) пластический (конструктивный):а) анаболизм (с затратами энергии);б) катаболизм (с выделением энергии);2) энергетический обмен (протекает в дыхательных
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы Пер. А. НеизвестногоВ предыдущей главе мы обсудили возможные сценарии возникновения клеток и (будем надеяться) достигли
Глава 4. Регуляция активности ферментов. Медицинская энзимология
Глава 4. Регуляция активности ферментов. Медицинская энзимология Способы регуляции активности ферментов:1. Изменение количества ферментов.2. Изменение каталитической эффективности фермента.3. Изменение условий протекания реакции.Регуляция количества
Глава 17. Пути метаболизма глюкозы
Глава 17. Пути метаболизма глюкозы Глюкоза является основным метаболитом и транспортной формой углеводов в организме человека и животных. Источниками глюкозы являются углеводы пищи, гликоген тканей и процесс глюконеогенеза в печени и корковом веществе почек. Для
Взаимосвязь метаболизма
Взаимосвязь метаболизма Метаболизм в целом не следует понимать как сумму обменов белков, нуклеиновых кислот, углеводов и липидов. В результате взаимодействия обменов отдельных классов органических соединений возникает единая система метаболических процессов,
Особенности метаболизма в форменных элементах крови
Особенности метаболизма в форменных элементах крови Эритроциты:1. Зрелые эритроциты лишены ядра, поэтому в клетке не синтезируются белки. Эритроцит почти целиком заполнен гемоглобином.2. Эритроциты не имеют митохондрий, поэтому в клетке не протекают реакции ЦТК, ЦТД,
Глава 32. Особенности метаболизма в нервной ткани
Глава 32. Особенности метаболизма в нервной ткани Человеческий мозг – это самая сложная из всех известных живых структур. Нервной системе и, в первую очередь, головному мозгу принадлежит важнейшая роль в координации поведенческих, биохимических, физиологических
Глава 7 Генетическая регуляция развития
Глава 7 Генетическая регуляция развития У природы столько дел в этом мире, ей приходится создавать такую массу разнообразнейших творений, что по временам она и сама не в силах разобраться во всех тех различных процессах, которыми она одновременно занимается. Уилки
Глава 1 Взаимосвязь функций организма
Глава 1 Взаимосвязь функций организма Психические явления – это следствия химических процессов в организме? – ДА!Психические явления можно объяснить химическими процессами? –