Рассказ Онихофоры
Рассказ Онихофоры
Если современная зоология и признает нечто похожее на миф о внезапном зарождении жизни, то это, безусловно, “кембрийский взрыв”. Кембрий – первый период фанерозойского эона, начавшегося около 545 млн лет назад. Именно в кембрийском периоде животная и растительная жизнь – такая, какой мы ее знаем, – стала появляться в окаменелостях. Все окаменелости старше кембрия представляют собой либо незначительные следы, либо загадки. Однако в кембрийском периоде на свет появился шумный зверинец, предвещающий современность. Неожиданность, с которой многоклеточные ископаемые появляются в начале кембрия, напоминает взрыв.
Креационисты очень любят “кембрийский взрыв” – видимо, потому, что их скудному воображению он представляется чем-то вроде палеонтологического сиротского приюта, населенного безродными таксонами: животными без прошлого, которые будто в одночасье возникли из ничего. С другой стороны, романтически настроенные зоологи тоже любят “кембрийский взрыв”. Они видят в нем своего рода Аркадию, эпоху зоологической невинности, в которой жизнь безумствовала, развиваясь в бешеном эволюционном темпе. То была безгрешная вакханалия жизненных форм, предваряющая скучный прагматизм, преобладающий в современном мире. В книге “Расплетая радугу” я процитировал выдающегося биолога (возможно, впрочем, он уже изменил свое мнение):
Вскоре после того, как были изобретены многоклеточные формы, прозвучал великий эволюционный взрыв. Кажется, можно физически ощутить, как многоклеточная жизнь с радостью торопится попробовать все варианты в диком танце беспечного исследования.
Если и есть животное, которое больше чем кто-либо еще символизирует такое представление о кембрии, то это галлюцигения (Hallucigenia). Это животное (Саймон Конвей-Моррис, описавший ее, выбрал это название намеренно) было сначала реконструировано вверх тормашками. Вот почему на рисунке галлюцигения стоит на невозможных ходулях-зубочистках. Согласно современной, перевернутой реконструкции, одинокий ряд “щупалец” вдоль спины представляет собой ноги. Но как же она ходила – неужто балансировала, как на канате? Нет, ископаемые, обнаруженные в Китае, указывают на то, что у галлюцигении был и второй ряд ног. И на современной реконструкции это животное выглядит вполне приспособленным к жизни. Сегодня галлюцигения уже не кажется “таинственным чудом” с неясными, давно утраченными родственными связями. Вместе с некоторыми другими кембрийскими ископаемыми теперь ее ориентировочно относят к типу лопастеногих (Lobopodia). Среди современных представителей этого типа – перипатус и другие онихофоры (см. рандеву № 26).
В те времена, когда кольчатых червей было принято считать близкими родственниками членистоногих, онихофор нередко называли “промежуточным звеном” между этими двумя группами. Это не совсем верно, если задуматься о том, как идет эволюция. Сейчас кольчатых червей относят к спиральным (Lophotrochozoa), а онихофор (Onychophora) с членистоногими (Arthropoda) – к линяющим (Ecdysozoa). Перипатуса с его древними связями разместили среди современных пилигримов, и именно он расскажет нам о “кембрийском взрыве”.
Онихофоры широко распространены в тропиках, особенно в Южном полушарии. Изображенная здесь онихофора Peripatopsis moseleyi обитает в Южной Африке. Все современные онихофоры живут на суше, в опавших листьях и других влажных местах, где охотятся на мелких животных: улиток, червей, насекомых и так далее. В кембрийском периоде галлюцигения и другие далекие предки перипатуса и перипатопсиса жили, как и все остальные животные, в море.
Связь галлюцигении с современными онихофорами до сих пор спорна. Нельзя забывать, что между полуистершимся сплющенным ископаемым в камне и красочной реконструкцией на странице всегда есть место для фантазии. Высказано даже предположение, что галлюцигения – это не целое животное, а лишь часть некоего другого животного. Такие примеры известны. На ранних художественных реконструкциях кембрийской фауны можно увидеть плавающее медузоподобное существо, похожее на консервированные кольца ананаса. Как выяснилось позднее, эти существа представляли собой части челюстного аппарата хищного аномалокариса (Anomalocaris). Другие кембрийские ископаемые, например Aysheaia, очень похожи на морские версии перипатуса, и это дает перипатусу дополнительное право на рассказ о кембрии.
Большая доля окаменелостей любой эпохи представляет собой остатки твердых частей животных: костей позвоночных, панцирей членистоногих, раковин моллюсков или плеченогих. Но существует три кембрийских местонахождения – в Канаде, в Гренландии и в Китае, – где из-за необычных условий сохранились и следы мягких тканей животных (лагерштетты). Это сланцы Берджес в Британской Колумбии, Сириус-Пассет на севере Гренландии и Чэнцзян в Южном Китае. Сланцы Берджес были открыты в 1909 году, однако получили известность лишь 80 лет спустя благодаря книге “Чудесная жизнь” Стивена Джея Гулда. Отложение Сириус-Пассет на севере Гренландии было обнаружено в 1984 году, однако пока оно изучено хуже двух указанных. В том же году Хоу Сяньгуан обнаружил окаменелости в Чэнцзяне. Доктор Хоу участвовал в подготовке прекрасно иллюстрированной монографии “Кембрийские окаменелости Чэн-цзяна (Китай)”, изданной в 2004 году, – к счастью для меня, это случилось непосредственно перед тем, как эта книга вышла в печать.
Окаменелости Чэнцзяна датируются 525 млн лет. Их возраст примерно равен возрасту Сириус-Пассет и на 10–15 млн лет превышает возраст сланцев Берджес. Однако все эти знаменитые залежи включают сходную фауну. В них встречается большое число лопастеногих, более или менее похожих на морские версии перипатуса. Есть морские водоросли, губки, черви, а также плеченогие, довольно похожие на современных, и загадочные животные с неясными родственными связями. Там находят много членистоногих, включая ракообразных, трилобитов и других животных, отдаленно напоминающих ракообразных и трилобитов, но, возможно, принадлежащих к обособленным группам. В Чэнцзяне и Берджес найдены крупные (иногда более метра в длину), судя по всему, хищные аномалокарисы и родственные им животные. Ученые пока точно не знают, кем они были – возможно, дальними родственниками членистоногих, – но, судя по всему, это были весьма эффектные животные. Некоторые “таинственные чудеса” открыты в сланцах Берджес, но не обнаружены в Чэнцзяне – например пятиглазая опабиния (Opabmia).
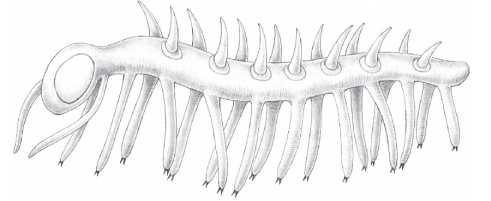
Галлюцигения (Hallucihenia).
Фауна Сириус-Пассет включает красивое животное халкиерию (Halkieria). Сначала ее считали древним моллюском, однако Саймон Конвей-Моррис, описавший много странных кембрийских животных, видит в ней сходство с тремя главными типами беспозвоночных: моллюсками, плеченогими и кольчатыми червями. Мне это нравится: он противоречит тому мистическому благоговению, которое зоологи испытывают по отношению к большим типам (см. вкладку). Если мы относимся к эволюции серьезно, то должны понимать, что по мере движения вспять во времени и приближения к точкам дивергенции типы неизбежно становятся все сильнее схожими и связанными между собой. Было бы, напротив, странно, если бы не существовало такого древнего животного, которое объединяет кольчатых червей, плеченогих и моллюсков. Обратите внимание на раковины на рисунке (см. вкладку).
Как мы увидели на рандеву № 22, в Чэнцзяне встречаются ископаемые, очень похожие на настоящих позвоночных, предшествующих похожей на ланцетника пикайе из сланцев Берджес и другим кембрийским хордовым. Согласно традиционным зоологическим представлениям, позвоночные не могли возникнуть так рано. Однако миллокунмингия (Myllokunmigia), известная по полусотне экземпляров из Чэнцзяна, очень похожа на типичных бесчелюстных рыб, которые возникли, как считалось, лишь 50 млн лет спустя, в середине ордовикского периода. Сначала были описаны два новых рода: миллокунмингия (Myllokunmingia) – как считалось, она родственна миногам, и хайкоуихтис (Haikouichthys) (к сожалению, назван он не в честь японской стихотворной формы), предположительно родственный миксинам. Сегодня некоторые специалисты по систематике объединяют их в один вид – Myllokunmingia fengjiaoa. Эта спорная ревизия статуса хайкоуихтиса – хороший пример того, как сложно бывает разобраться в очень древних ископаемых. Здесь приведена фотография миллокунмингии, а также рисунок, сделанный с помощью камеры-люциды. Меня восхищает терпение ученых, реконструирующих древних животных.
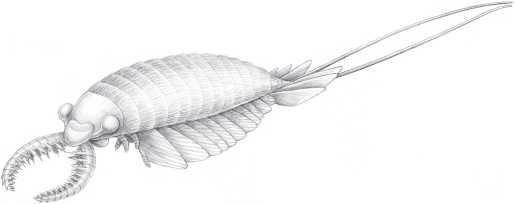
Аномалокарис (Anomalocaris saron).
Продление истории позвоночных до середины кембрийского периода говорит в пользу идеи “взрыва”. Действительно, большинство основных типов животных впервые появляется в палеонтологической летописи в узком временном промежутке – в кембрии. Это, конечно, не значит, что до кембрия никаких представителей этих типов не существовало. Просто они, как правило, не превращались в окаменелости. Как это объяснить? На этот счет есть три основных гипотезы (и различные их комбинации). Все они напоминают гипотезы относительно “взрывного” распространения млекопитающих после исчезновения динозавров.
“Взрыва” не было. Согласно этой точке зрения, “взрыв” касался лишь способности к фоссилизации, а не эволюции. Типы появились задолго до кембрийского периода, их предки жили сотни миллионов лет. Такое представление поддерживают некоторые молекулярные биологи, которые с помощью “молекулярных часов” датировали ключевых предков этих типов. Грегори Рэй, Джеффри Левинтон и Люси Шапиро в известной статье 1996 года установили, что общий предок позвоночных и иглокожих жил около миллиарда лет назад, а общий предок позвоночных и моллюсков – еще на 200 млн лет раньше, то есть был более чем вдвое старше “кембрийского взрыва”. Метод “молекулярных часов” вообще имеет свойство отодвигать даты таких ветвлений далеко в докембрий – то есть гораздо дальше, чем хочется большинству палеонтологов. Согласно этой точке зрения, до кембрия ископаемые по неизвестным причинам сохранялись плохо. Возможно, дело в нехватке легко подвергающихся фоссилизации твердых тканей, например раковин, панцирей и костей. Ведь сланцы Берджес и Чэн-цзян необычны именно тем, что там встречаются окаменелые мягкие ткани. Возможно, докембрийские животные, несмотря на свое разнообразие и сложную организацию, были просто слишком мелкими, чтобы фоссилизоваться. В пользу этой версии говорит существование некоторых типов мелких животных, которые вообще не оставили окаменелостей после кембрия и известны нам только благодаря “живым ископаемым”. Так почему мы надеемся на обнаружение докембрийских ископаемых? Как бы то ни было, некоторые из найденных докембрийских ископаемых, включая эдиакарскую фауну и окаменелые отпечатки следов и нор, указывают на то, что докембрийские многоклеточные существовали.
Замедленный “взрыв”. Общие предки различных типов действительно жили довольно близко друг к другу по времени. Однако от окаменелостей времен “взрыва” их отделяет более чем несколько десятков миллионов лет. Отложения Чэнцзяна (525 млн лет) на первый взгляд кажутся довольно близкими к предполагаемому сопредку, который жил около 590 млн лет назад. Но на самом деле их разделяет ни много ни мало 65 млн лет – это столько же, сколько прошло с исчезновения динозавров до сегодняшнего дня, а ведь за это время появились все многочисленные группы млекопитающих, которые мы видим сейчас. Даже 10 млн лет – это очень много, учитывая взрывное видообразование, о котором говорилось в “Рассказе Галапагосского вьюрка” и “Рассказе Цихлиды”. Конечно, проще думать, что два древних ископаемых, принадлежащих сегодня к двум современным типам, в древности так же сильно различались, как и современные представители этих типов. Однако не нужно забывать, что у этих современных представителей было полмиллиарда лет, чтобы разойтись. Не обязательно кембрийский таксономист, лишенный груза накопленных за полмиллиарда лет знаний, поместил бы эти два ископаемых в отдельные типы. Возможно, он решил бы, что различия между ними существуют лишь на уровне отрядов – ведь он не знает, что их потомкам суждено разойтись настолько, что они будут принадлежать к разным типам.

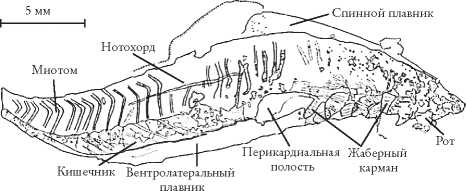
Никто и не думал, что позвоночные животные настолько древние. Ископаемая миллокунмингия (Myllokunmingia fengjiaoa), Чэнцзян. D.-G. Shu et al. [264].
Мгновенный “взрыв”. Эта точка зрения, на мой взгляд, бредовая. Или, выражаясь дипломатично, крайне нереалистичная и легкомысленная. Однако мне все же придется уделить ей внимание, потому что в последнее время она необъяснимо популярна – видимо, благодаря излишне романтичным зоологам.
Согласно этому взгляду, новые типы возникли внезапно, благодаря “взрыву” макромутаций. Вот изречения в остальном достойных ученых, которые я приводил в книге “Расплетая радугу”.
Похоже, что легкость, с которой совершались эволюционные скачки, приведшие к основным конструктивным новшествам – основа новых типов, – по каким-то причинам была утрачена после кембрийского периода. Как будто главный двигатель эволюции отчасти утратил свою мощность… Таким образом, эволюция кембрийских организмов могла совершать большие скачки, в том числе на уровне типов, однако позднее она стала сдержаннее и делала лишь короткие скачки в масштабе классов.
А вот слова ученого, о котором мы упоминали в начале рассказа:
В самом начале процесса ветвления мы обнаруживаем множество мутаций, которые довольно далеко отстоят от основного ствола и друг от друга. Такие виды имеют достаточно морфологических различий, чтобы их можно было рассматривать как основателей типов. Эти основатели также дают ответвления, однако не с помощью длинных скачков, а образуя ветви от основателя каждого типа к дочерним видам – основателям классов. По мере движения подходящие варианты образуются во все более близком соседстве, поэтому основатели отрядов, семейств и родов появляются последовательно.
Это все равно что садовник, посмотрев на старый дуб, озадаченно заметил бы:
Не странно ли, что за много лет на этом дереве не появилось ни одной новой толстой ветви? Сейчас на нем почему-то вырастают только тонкие веточки!
А вот цитата, автора которой я укажу: она из книги Эндрю Паркера “Во мгновение ока”, напечатанной уже после выхода “Расплетая радугу”. Книга Паркера в основном посвящена интересной и оригинальной теории автора, согласно которой “кембрийский взрыв” был вызван внезапным появлением у животных глаз. Однако прежде чем приступить к обсуждению теории, Паркер пишет о третьей версии мифа о “кембрийском взрыве”, принимая ее за чистую монету. Сначала он формулирует миф, причем в самой “взрывной” версии, о которой мне приходилось слышать:
Около 544 млн лет назад существовало три типа животных с разнообразными внешними формами, а 538 млн лет назад их было уже 38 – столько же, сколько сегодня.
Паркер поясняет, что не имеет в виду чрезвычайно быструю градуалистическую эволюцию за 6 млн лет – это был бы крайний вариант нашей второй гипотезы, да и то едва приемлемый. Не говорит он и то, что сказал бы я: вблизи точки расхождения два типа (точнее – предки эти типов) вряд ли сильно отличались бы друг от друга. Ведь они должны были пройти последовательные стадии пары видов, затем пары родов и так далее – пока не стали настолько различны, что их стало возможным отнести к типам. Паркер, судя по всему, считает, что все 38 типов полностью сформировались 538 млн лет назад в результате макромутационного прыжка:
Тридцать восемь типов животных эволюционировало на Земле. Таким образом, имели место тридцать восемь монументальных генетических событий, которые привели к возникновению тридцати восьми планов строения тела.
Монументальными генетическими событиями, конечно, пренебрегать не стоит. Мутация контролирующих генов из различных семейств Hox (см. “Рассказ Дрозофилы”), безусловно, может привести к потрясающему эффекту. Однако монументальное монументальному рознь. Дрозофила с парой ног вместо антенн – это очень сильный ход, но и в этом случае вопрос о выживании остается открытым. На то есть причина.
Мутантное животное с определенной долей вероятности может стать лучше вследствие этой новой мутации. “Лучше” в данном случае значит “лучше, чем родительский тип без мутации”. Однако родитель не мог быть так уж плох: по меньшей мере, он оказался достаточно хорош, чтобы оставить потомство. Очевидно, чем незначительнее мутация, тем с более высокой вероятностью она окажется полезной. “Очевидно” было любимым словом выдающегося специалиста по статистике и биологии Рональда Э. Фишера, и он нередко употреблял его по отношению к вещам, которые простым смертным были вовсе не очевидны. В данном случае, однако, понять Фишера довольно легко. Возьмем простой метрический признак, например длину бедра, которая варьирует в одном измерении, то есть может быть на несколько миллиметров больше или меньше.
Представьте себе ряд мутаций возрастающего масштаба. В начале ряда нулевая мутация. Она по определению не хуже, чем родительская копия гена, которая должна быть достаточно хороша, чтобы, по меньшей мере, обеспечить воспроизводство. Теперь представьте случайную незначительную мутацию: например такую, в результате которой нога становится на миллиметр длиннее или короче. Исходя из того, что родительский ген не безупречен, у мутации, которая бесконечно мало отличается от родительской версии, есть 50-процентный шанс стать лучше и 50-процентный – стать хуже. Она будет лучше, если результат ее – шаг в правильном направлении, и хуже, если результат – шаг в противоположном от родительского состояния направлении. Однако очень крупная мутация, скорее всего, окажется хуже родительской версии гена, даже если это будет шаг в верном направлении. Вообразите крайний случай: во всем нормального человека, однако с бедрами двухметровой длины.
Аргументы Фишера более общие. Когда мы говорим о макрому-тационных скачках, выводящих группу организмов на территорию нового типа, мы имеем дело не с простыми количественными параметрами вроде длины ног. Поэтому нам нужна другая версия этого аргумента. Как я уже говорил, есть гораздо больше способов погибнуть, чем уцелеть. Представьте себе математический ландшафт, населенный всеми возможными животными. Я называю его математическим, потому что у него сотни измерений, и он включает почти бесконечно широкий диапазон всех мыслимых монстров, а также и (относительно) небольшое число животных, когда-либо в реальности существовавших. То, что Паркер называет “монументальным генетическим событием”, эквивалентно макромутации огромного масштаба, действующей не в одном измерении, как в примере с бедром, а в сотнях одновременно. Таков масштаб изменений, с которыми мы столкнемся, если, подобно Паркеру, будем настаивать на мгновенном переходе от одного типа к другому.
В многомерном ландшафте, населенном всеми возможными животными, реальные существа суть островки жизнеспособности, разделенные океаном гротескного уродства. Начав с любого “острова”, можно постепенно эволюционировать: тут изменим ногу, там укоротим рога или перекрасим перья. Эволюция – это траектория через многомерное пространство, где каждому шагу соответствует тело, способное к выживанию и размножению не хуже, чем родительский тип, соответствующий предыдущему шагу. С течением времени траектория достаточной длины приведет от жизнеспособной отправной точки к жизнеспособной конечной точке, которая будет настолько сильно отличаться от исходной, что мы признаем ее новым типом – например, моллюсками. А другая траектория, начавшись из той же отправной точки, может через жизнеспособные промежуточные точки привести к другому жизнеспособному конечному пункту, который мы признаем другим типом – например, кольчатыми червями. Нечто подобное, должно быть, происходило в каждой точке ветвления, приводящей к появлению пары животных от общего предка.
Вот к чему я веду. Случайная мутация, дающая сразу новый тип, окажет такое влияние на сотню измерений одновременно, что нужно быть исключительно везучим, чтобы оказаться на другом островке жизнеспособности, а не посреди океана нежизнеспособности.
Креационисты неудачно уподобляют естественный отбор урагану, который проносится по свалке и случайно собирает “Боинг-747”. Они упускают из виду последовательную, кумулятивную природу естественного отбора. Но сравнение со свалкой соответствует гипотетическому внезапному появлению нового типа. Эволюционный шаг такого масштаба, как, например, моментальный переход от земляного червя к улитке, действительно является таким же невероятным, как ураган на свалке.
Итак, теперь мы можем с уверенностью отклонить третью гипотезу. Остаются две – или некий компромисс между ними. Я в этом случае предпочту не делать выводов до появления дополнительных данных. Сейчас все больше ученых признает, что показания “молекулярных часов” отодвигали основные точки ветвления на сотни миллионов лет назад, в докембрий. С другой стороны, тот факт, что ископаемые большинства типов почти не встречаются в докембрийских отложениях, не должен заставлять нас думать, что все эти типы быстро эволюционировали. Пример с ураганом указывает на то, что кембрийские ископаемые наверняка имели непрерывную цепочку предков. Эти предки существовали, но по каким-то причинам оказались не способны к фоссилизации. На первый взгляд идея, что множество животных 100 млн лет могло оставаться невидимым, кажется невероятнее, чем если бы они оставались невидимыми лишь 10 млн лет. По этой причине некоторым больше нравится гипотеза замедленного “кембрийского взрыва”. С другой стороны, чем внезапнее взрыв, тем труднее поверить, что все это многообразие могло появиться за такое короткое время. Этот аргумент – палка о двух концах, и он не может помочь нам сделать выбор между двумя гипотезами.
Многоклеточная жизнь присутствует в палеонтологической летописи еще до Чэнцзяна и Сириус-Пассет. Около 20 млн лет ранее, на границе кембрия и докембрия, появляется множество микроскопических ископаемых, которые сильнее всего похожи на раковинки. Их так и называют: “мелкораковинная фауна”. Для большинства палеонтологов стало неожиданностью, когда некоторые “раковинки” оказались чешуйками (склеритами) панциря лопастеногих – родственников онихофор. Это означает, что расхождение между группами первичноротых произошло еще до “кембрийского взрыва”.
Есть и более древние свидетельства разнообразия животных. За 20 млн лет до кембрия, в эдиакарском периоде позднего докембрия, была повсеместно распространена загадочная эдиакарская фауна (по названию возвышенности Эдиакара в Южной Австралии, где ее впервые обнаружили). Трудно сказать, что собой представляли эти животные, однако то были одни из первых существ, от которых остались окаменелости. Некоторые, возможно, были губками. Другие напоминают медуз. Есть похожие на актиний или на морские перья (это перьевидные родственники актиний). Некоторые похожи на червей или слизняков и, возможно, имели настоящую двустороннюю симметрию. Большинство же из них совершенно загадочны. Например, что думать о дикинсонии (Dickmsonia)? Это коралл? Червь? Гриб? (См. вкладку.) Или нечто совсем не похожее на все, что мы знаем? К эдиакарской фауне относится очень похожее на головастика ископаемое из Австралии. Формально оно еще не описано, но предполагают, что относится оно к хордовым (это, напомню, тип, к которому принадлежат позвоночные). Это было бы потрясающе. Но время покажет. Зоологи признают, что эдиакарская фауна пока не слишком помогает проследить родословную большинства современных животных.
Существуют также отпечатки окаменелостей, которые ученые считают следами или норами докембрийских животных. Это указывает на существование достаточно крупных ползающих животных. Но, к сожалению, следы почти ничего не говорят о том, каковы были эти животные. В Доушаньто (Китай) найдены еще более древние, главным образом микроскопические, окаменелости. Они похожи на эмбрионов, хотя неясно, какие животные из этих эмбрионов вырастали. Еще древнее дискообразные отпечатки из Северо-Западной Канады (600–610 млн лет). Эти животные, пожалуй, еще загадочнее эдиакарской фауны.
Мне бы хотелось привести оценки возраста для каждого из 39 рандеву в этой книге. Большую часть можно датировать вполне уверенно с помощью окаменелостей и “молекулярных часов”, откалиброванных с помощью таких окаменелостей. Однако очевидно: чем глубже в прошлое мы заходим, тем чаще окаменелости нас подводят. Мы уже не можем надежно откалибровать “молекулярные часы” и входим в область, не поддающуюся датировке. Для полноты картины я все же привел примерные датировки жизни сопредков №№ 23–39. Последние данные, мне кажется, поддерживают (хотя бы отчасти) гипотезу о замедленном “взрыве”. Прежде я склонялся скорее к тому, что никакого взрыва не было вообще. Когда мы получим новые данные (на что я очень надеюсь), я не удивлюсь, если они отодвинут нас еще глубже в докембрий, где мы будем искать общих предков современных типов. Или, возможно, новые данные укажут нам на мгновенный “взрыв” и на то, что общие предки основных типов животных сменяли друг друга на коротком временном отрезке в 20, даже 10 млн лет в начале кембрия. И в этом случае я уверен, что даже если мы правильно отнесем двух кембрийских животных к различным типам на основании их сходства с современными животными, в кембрии они были гораздо ближе друг к другу, чем их нынешние потомки. Кембрийские зоологи поместили бы их не в отдельные типы, а лишь, например, в подклассы.
Я не удивился бы, увидев, что любая из этих двух гипотез доказана. Сам я не хочу брать на себя этот риск. Но я скорее съем свою шляпу, чем поверю, что кто-то докажет третью гипотезу. У нас есть все основания думать, что эволюция в кембрийском периоде была по сути такой же, что сейчас. Что касается рассуждений о распрямившейся после кембрия пружине эволюции и новых типах, внезапно появившихся в райских лучах рассвета беззаботного экспериментаторства, то я ответственно заявляю, что все это – глупость.
Спешу добавить, что я не имею ничего против поэзии кембрия. Но предпочитаю версию Ричарда Форти, изложенную в прекрасной книге “Жизнь: неавторизованная биография”:
Представляю, как однажды вечером стою на кембрийском берегу – примерно так, как стоял на берегу архипелага Шпицберген, когда впервые задумался о биографии жизни. Прибой был бы почти таким же. Там, где море встречается с сушей, лежала бы полоса чуть липких округлых строматолитовых матов, оставшихся от обширных зарослей докембрия. Ветер дул бы в спину с красных равнин без признаков жизни, и по ногам меня хлестали бы острые песчинки. А на грязном песке у ног я видел бы следы червя – знакомые узкие извивающиеся бороздки. Я видел бы следы, оставленные спрятавшимися ракообразными… И не слышал бы ничего, кроме ветра и прибоя.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Рассказ Евы
Рассказ Евы Есть впечатляющая разница между «родословными деревьями генов» и «родословными деревьями людей». В отличие от человека, который происходит от двух родителей, у гена есть только один родитель. Каждый из Ваших генов должен был быть получен или от Вашей матери,
Рассказ Секвойи
Рассказ Секвойи Люди спорят о том, какое одно место в мире Вы должны посетить прежде, чем умрете. Мой кандидат – лес Muir Woods, несколько севернее моста «Золотые Ворота». Или, если Вы считаете, что слишком поздно, я не могу вообразить лучшего места, чтобы быть похороненным (вот
Рассказ Taq
Рассказ Taq Достигнув нашего самого древнего свидания, собрав в нашем странствии всю жизнь, которую знаем, мы имеем возможность рассмотреть ее разнообразие. На самом глубинном уровне разнообразие жизни является химическим. Профессии, которыми заняты наши
Рассказ Земледельца
Рассказ Земледельца Аграрная революция началась около 10 тыс. лет назад, на исходе последнего ледникового периода, между Тигром и Евфратом – в регионе, прозванном “плодородным полумесяцем”. Другие очаги земледелия возникли (скорее всего, независимо) в Китае, долине Нила
Рассказ Stw 573
Рассказ Stw 573 Не думаю, что есть смысл придумывать причины, в силу которых ходить на двух ногах – это здорово. Будь так, шимпанзе делали бы то же самое, не говоря уже о других животных. Нет причины, по которой бег на двух или четырех конечностях быстрее или удобнее.
Рассказ Гиббона[12]
Рассказ Гиббона[12] На рандеву № 4 мы встречаем крупную группу пилигримов. И теперь могут возникнуть проблемы с установлением родства. (Чем дальше, тем затруднительнее это сделать.) Существует двенадцать видов гиббонов, принадлежащих к четырем основным группам. Это Bunopithecus
Рассказ Ревуна[18]
Рассказ Ревуна[18] Новые гены появляются не из воздуха. Они образуются при дупликации прежних генов. После этого каждый из них идет своим путем, приобретая мутации и подвергаясь действию естественного отбора и дрейфа генов. Обычно мы не видим, как это происходит, но,
Рассказ Айе-айе
Рассказ Айе-айе Один британский политик как-то описал конкурента (который позднее стал лидером партии) как человека, “в котором есть что-то ночное”. При взгляде на айе-айе складывается именно такое впечатление, и это не случайно: она ведет целиком ночной образ жизни. Из
Рассказ Шерстокрыла
Рассказ Шерстокрыла Шерстокрыл из Юго-Восточной Азии мог бы рассказать нам, каково это – парить во воздуху ночного леса. Но для нас, пилигримов, у него приготовлена гораздо более приземленная история. Мораль ее в том, что составленная нами схема с сопредками, рандеву и
Рассказ Мыши
Рассказ Мыши Из тысяч грызунов домашняя мышь (Mus musculus) может поведать кое-что особенное: ни один другой вид млекопитающих, кроме нашего собственного, мы не изучали так интенсивно. Именно на мышах, а не на пресловутых морских свинках держатся медицинские, физиологические и
Рассказ Бобра
Рассказ Бобра “Фенотип” – это то, что находится под влиянием генов. В целом под фенотипом понимают все, что относится к телу. Но есть тонкости, вытекающие из этимологии. Phamo по-гречески означает “показывать”, “обнаруживать”, “выявлять”, “выражать”, “раскрывать”.
Рассказ Гиппопотама
Рассказ Гиппопотама В школе, изучая греческий, я узнал, что “гиппос” означает “лошадь”, а “потамос” – “река”. Выходило, что гиппопотамы – “речные лошади”. Позднее, забросив греческий и занявшись зоологией, я узнал, что гиппопотамы не имеют отношения к лошадям. В
Рассказ Тюленя
Рассказ Тюленя В большинстве естественных популяций число самцов и самок примерно одинаково. На то есть эволюционные причины, и о них рассказал замечательный специалист по статистике и эволюционной генетике Рональд Э. Фишер. Представьте себе популяцию, количество в
Рассказ Броненосца
Рассказ Броненосца С зоологической точки зрения Южная Америка сродни Мадагаскару: как и Мадагаскар, она откололась от Африки – но не с восточной, а с западной стороны. Произошло это примерно тогда же или чуть позднее. Как и Мадагаскар, в течение почти всего времени
Эпилог к рассказу Онихофоры
Эпилог к рассказу Онихофоры Большую часть книги я беззаботно разбрасывался датировками рандеву и даже опрометчиво оценивал сопредков как прародителей в том или ином поколении. Мои датировки главным образом основаны на ископаемых. Однако допущение при таком
Рассказ Taq
Рассказ Taq Итак, мы встретили практически все существующие формы жизни. Теперь можно окинуть взглядом открывшееся разнообразие. На самом глубоком уровне разнообразие жизни является химическим. Профессии, которыми владеют наши пилигримы, охватывают широкий диапазон