Рассказ Бобра
Рассказ Бобра
“Фенотип” – это то, что находится под влиянием генов. В целом под фенотипом понимают все, что относится к телу. Но есть тонкости, вытекающие из этимологии. Phamo по-гречески означает “показывать”, “обнаруживать”, “выявлять”, “выражать”, “раскрывать”. Фенотип – это внешнее, видимое проявление скрытого генотипа. Оксфордский словарь английского языка определяет его как “совокупность всех поддающихся наблюдению признаков индивида, рассматриваемых как следствие взаимодействия его генотипа с окружающей средой”, но предваряет это определение еще одно: “Тип организма, отличающийся от других поддающимися наблюдению признаками”.
Дарвин рассматривал естественный отбор как выживание и размножение определенных типов организмов за счет других, конкурирующих с ними. Под типами здесь не подразумеваются группы, породы или виды. Фраза “сохранение благоприятствуемых пород” из подзаголовка “Происхождения видов”, так часто понимаемая неверно, никоим образом не подразумевает породы в обычном смысле этого слова. Дарвин писал это еще до того, как генам дали имена, до того, как кто-то понял, что это такое, но, выражаясь современным языком, под “благоприятствуемыми породами” он имел в виду “носителей благоприятствуемых генов”.
Отбор направляет эволюцию лишь в той степени, в которой альтернативные типы обязаны генам своими различиями: если различия не наследуются, то избирательное выживание никак не влияет на будущие поколения. Для дарвиниста фенотипы – это проявления, по которым естественный отбор оценивает гены. Когда мы говорим, что хвост бобра уплощен, чтобы служить веслом, мы имеем в виду, что гены, фенотипическое проявление которых обеспечило уплощение хвоста, выживали благодаря преимуществам такого фенотипа. Отдельные бобры с плоскохвостым фенотипом выживали потому, что лучше плавали, а в них выживали отвечающие за это гены, которые затем передавались новым поколениям плоскохвостых бобров.
Вместе с этим выживали также гены, проявлявшиеся в острых резцах. Бобры получаются за счет перетасовок генов в генофонде. Гены выживали в ряду поколений боровых предков благодаря тому, что хорошо зарекомендовали себя в сотрудничестве с другими генами из генофонда бобра и смогли обеспечить фенотипы, преуспевающие в тех условиях жизни, в которых живет бобр.
В то же время в других генофондах сохранялись альтернативные кооперации генов, создающие тела, которые выживали, занимаясь другими “ремеслами”: тигриная кооперация, верблюжья кооперация, тараканья кооперация, морковная кооперация. Моя первая книга “Эгоистичный ген” вполне могла бы называться “Кооперативный ген”, причем в самой книге при этом не нужно было бы менять ни слова. Более того, это могло бы предотвратить некоторые недоразумения (отдельные литературные критики довольствуются чтением одного только заголовка). Эгоизм и кооперация – это две стороны дарвиновской медали. Каждый ген действует в своих интересах; кооперируясь с другими генами в генофонде, который тасуется в результате полового размножения и служит для гена окружающей средой, гены строят новые тела.
Но гены бобров создают фенотипы, сильно отличающиеся от фенотипов тигров, верблюдов и моркови. Фенотип бобра – это запруда, обусловленная фенотипом плотины. Запруда – расширенный фенотип, особый род фенотипа [37]. Это интересно не только само по себе, но и потому, что так легче понять, как развиваются фенотипы в обычном понимании этого слова. Мы увидим, что между расширенным фенотипом, таким как запруда, и обычным фенотипом, таким как уплощенный хвост, разница не так уж и велика.
Как можно использовать одно и то же слово – фенотип – для обозначения, с одной стороны, хвоста, а с другой – массива стоячей воды, образованного плотиной в русле реки? Дело в том, что и то, и другое – проявления генов бобра. И то, и другое эволюционировало, чтобы как можно лучше сохранять эти гены. И то, и другое связано с генами, которые они экспрессируют, сходными цепочками эмбриологических причинно-следственных связей.
Эмбриологические процессы, с помощью которых гены бобра создают хвосты, изучены пока не очень подробно, но мы понимаем, какого рода вещи там происходят. Гены в каждой клетке бобра ведут себя так, как будто “знают”, в какой они клетке. В клетках кожи те же гены, что и в костных клетках, однако в каждой ткани “включаются” разные гены (см. “Рассказ Мыши”). Во всех клетках бобрового хвоста гены ведут себя, будто “знают”, где они, и заставляют соответствующие клетки взаимодействовать так, что хвост в итоге становится плоским и лысым. Невероятно трудно выяснить, откуда гены “знают”, в какой части хвоста они находятся, но мы понимаем, как можно преодолеть эти трудности. Решение этих проблем и сами проблемы в общих чертах одинаковы, рассматриваем ли мы развитие тигриных лап, верблюжьего горба или морковных листьев.
Они примерно одинаковы и при развитии нейронных и нейрохимических механизмов, управляющих поведением. Копуля-тивное поведение бобров инстинктивно. Посредством выделения в кровь гормонов, а также с помощью нервов, контролирующих мышцы, которые двигают искусно сочлененными костями, мозг самца бобра дирижирует симфонией движений. Результат – точная координация действий с самкой. Можно с уверенностью сказать, что эта безупречная нейромышечная музыка усовершенствована поколениями естественного отбора. А это подразумевает отбор генов. В генофонде бобров выживали гены, фенотипические проявления которых в мозге, нервах, мышцах, железах, костях и органах чувств увеличивали шансы этих генов дожить до современности.
“Гены поведения” выживают точно так же, как и “гены костей” или “гены кожи”. Вы возразите, что нет генов поведения, а есть лишь гены нервов и мышц, из движений которых складывается поведение. Значит, вы еще одурманены грезами. Анатомические структуры не обладают особенным статусом по сравнению с поведенческими структурами в отношении “прямого” влияния генов. Гены “фактически” или “напрямую” отвечают только за белки и другие непосредственные биохимические реакции. Все остальное (будь то влияние на анатомические или поведенческие фенотипические проявления) – косвенное. Но провести границу между прямым и косвенным влиянием невозможно. В дарвинистском смысле важно только, что различия между генами приводят к различиям фенотипов. Естественному отбору интересны лишь различия. Примерно в том же смысле генетикам интересны лишь различия.
Помните “более тонкое” определение фенотипа: “Тип организма, отличающийся от других поддающимися наблюдению признаками”? Ключевое слово здесь – “отличающийся”. “Ген карих глаз” – это не ген, непосредственно кодирующий синтез коричневого пигмента. Да, так бывает, но дело не в этом. А в том, что обладание “геном карих глаз” обеспечивает различия в цвете глаз по сравнению с другой, альтернативной версией гена – “аллелем”. Причинно-следственные цепочки, приводящие в итоге к различиям между одним фенотипом и другим – скажем, между карими и голубыми глазами, – обычно длинны и извилисты. Ген производит белок, который отличается от белка, производимого альтернативным геном. Этот белок оказывает ферментативный эффект на клеточные химические процессы, которые влияют на икс, который влияет на игрек, который влияет на… длинную цепочку промежуточных факторов, которые влияют на… интересующий нас фенотип. Аллель обеспечивает различия при сравнении данного фенотипа с соответствующим фенотипом другого альтернативного аллеля, возникающим в конце длинной цепочки причинно-следственных связей, которую запускает этот аллель. Различия в генах приводят к различиям фенотипов. Изменения в генах приводят к изменениям фенотипов. В эволюции выбор аллелей происходит на основе различий их фенотипических проявлений.
Вот что хочет сказать нам бобр: сравнение фенотипов может происходить в любом месте причинно-следственной цепочки. Все промежуточные звенья представляют собой настоящие фенотипы, и любой может обеспечивать эффект, на основании которого и выбирается ген: он просто должен быть “заметен” для естественного отбора, причем не важно, заметен ли он при этом для нас. В цепочке нет “последнего звена”: не существует никакого итогового, конечного фенотипа. Любое следствие изменений аллелей, всегда и везде, какой бы длинной ни была цепочка причинно-следственных связей, представляет для естественного отбора законную добычу, пока он обеспечивает выживание данного аллеля относительно его конкурентов.
А теперь рассмотрим цепочку эмбриологических причинно-следственных связей, приводящую к тому, что бобры строят плотины. Строительство плотин – это сложный поведенческий стереотип, встроенный в мозг подобно хорошо настроенному часовому механизму. Или, если проследить историю часов до эпохи электроники, строительство плотин “зашито” в мозг. Я видел замечательный фильм о бобрах, которых запустили в необорудованную клетку без воды и древесины. Бобры впустую проигрывали все стереотипные движения, которые они обычно совершают в природе, где есть настоящие деревья и вода. Они помещали воображаемое дерево в воображаемую стену плотины, трогательно пытаясь построить призрачную стену из призрачных веток, и все это на твердом, сухом, плоском полу своей тюрьмы. Их можно пожалеть: они как будто отчаянно стараются использовать свой ненужный здесь механизм.
Такой мозговой механизм есть лишь у бобров. У других видов есть механизмы для копуляции, почесывания и драк. У бобров они тоже есть. Но лишь бобры имеют механизм для постройки плотин, который должен был постепенно эволюционировать у бобровых предков. А эволюционировал он потому, что запруды, создаваемые плотинами, очень полезны. Для чего именно, пока не слишком понятно, но ясно, что они полезны и нашим современникам-бобрам, а не только древним бобрам. Вероятнее всего, запруды образуют безопасные места для постройки хатки, недосягаемые для большинства хищников, и безопасные каналы для фуражировки. Какой бы ни была эта польза, она должна быть существенной, иначе бобры не тратили бы столько времени и сил на строительство плотин. Хочу снова обратить ваше внимание на то, что естественный отбор – это предсказательная теория. Дарвинист может уверенно предсказать, что если бы плотины были бесполезными, бобры-конкуренты, воздержавшиеся от их строительства, выживали бы чаще и передавали потомкам свою генетическую нелюбовь к плотинам. Тот факт, что бобры стремятся к строительству плотин, – веское доказательство того, что их предки извлекали из этого пользу.
Как и любая другая полезная адаптация, мозговой механизм строительства плотин должен был эволюционировать путем отбора генов. В системе проводки мозга, отвечающей за строительство плотин, должны были существовать генетические варианты, обеспечивавшие строительство плотин повышенного качества, которые с более высокой вероятностью выживали в генофонде бобров. То же происходит со всеми остальными дарвиновскими адаптациями. Но что из этого есть фенотип? На каком звене в цепочке причинно-следственных связей мы можем сказать: здесь генетические различия проявили свой эффект? Отвечу снова: на любом звене, где различие это заметно. Система проводки мозга? Да, наверняка. Клеточные химические процессы, которые в ходе эмбриологического развития создают такую систему? Конечно. А также поведение – симфония мышечных сокращений, которая и составляет поведение, – это самый настоящий фенотип. Различия в строительном поведении – это, без сомнения, проявления различий в генах. И, точно так же, следствия этого поведения вполне можно назвать фенотипом генов. Какие следствия? Плотины, конечно, а также запруды, поскольку они являются следствиями плотин. На различия между запрудами влияют различия между плотинами, как на различия между плотинами влияют различия между поведенческими паттернами, которые, в свою очередь, следствия различий между генами. Можно сказать, что свойства плотин или запруд – это настоящие фенотипические проявления генов.
Обычно биологи рассматривают фенотипические проявления гена как нечто, ограниченное кожей особи, несущей этот ген. Но из “Рассказа Бобра” видно, что это необязательно так. Фенотип гена может выходить за пределы организма. Птичьи гнезда – это расширенные фенотипы. Их формы и размеры, вычурные трубы и воронки, которые строят некоторые птицы, – дарвиновские адаптации, которые должны были эволюционировать за счет избирательного выживания альтернативных генов. Гены строительного поведения? Да. Гены, создающие систему проводки в мозге так, чтобы мозг умел строить гнезда правильной формы и размера? Да. Гены гнезд правильной формы и размера? Да. Но ведь гнезда строят из травы, веток или грязи, а не из клеток птицы? Да, но это неважно, когда мы говорим, влияют ли различия между генами на различия между гнездами. Если влияют, то гнезда – настоящие фенотипы генов. А различия между генами, несомненно, влияют на различия между гнездами, иначе как они совершенствовались бы в ходе естественного отбора?
Такие вещи, как гнезда или плотины (и запруды), – вполне очевидные примеры расширенных фенотипов (см. вкладку). Но есть и другие примеры, где логика немного более… ну, широкая. Например, можно сказать, что гены паразита фенотипически проявляются в телах хозяев. Это утверждение верно и тогда, когда паразиты не живут в телах хозяев, как, например, кукушки. Многие примеры коммуникации у животных – например, когда самец канарейки поет для самки и ее яичники растут, – можно переписать на языке расширенного фенотипа. Но это уведет нас слишком далеко от бобра, чей рассказ мы завершим так: в благоприятных условиях запруда может простираться на несколько километров, и это делает ее самым большим в мире фенотипом гена.
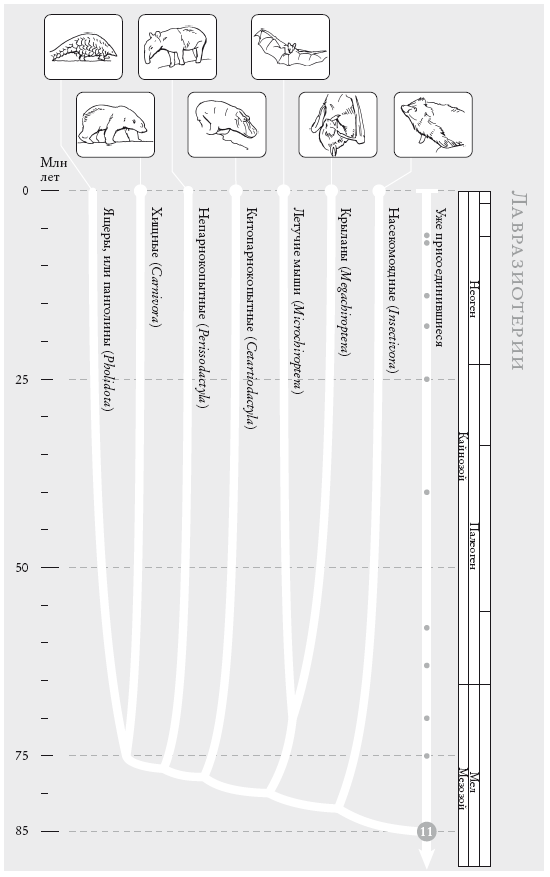
Лавразиотерии. В начале XXI века генетические исследования совершили переворот в систематике млекопитающих. Сейчас выделяют четыре основных группы плацентарных млекопитающих. Одна из них – группа, к которое относимся мы сами (она состоит в основном из грызунов и приматов). Ближайшая родственная нам группа включает ок. 2 тыс. видов лавразиотериев. Представленную здесь филогению лавразиотериев сторонники новой систематики млекопитающих считают достаточно надежной.
Нарис. (слева направо): саванный панголин, или степной ящер (Manis temminckii); белый медведь (Ursus maritimus); чепрачный тапир (Tapirus indicus); обыкновенный бегемот, или гиппопотам (Hippopotamus amphibius); австралийский ложный вампир (Macroderma gigas); индийская летучая лисица (Pteropus giganteus); обыкновенный, или европейский, еж (Erinaceus europaeus).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Рассказ Евы
Рассказ Евы Есть впечатляющая разница между «родословными деревьями генов» и «родословными деревьями людей». В отличие от человека, который происходит от двух родителей, у гена есть только один родитель. Каждый из Ваших генов должен был быть получен или от Вашей матери,
Рассказ Бобра
Рассказ Бобра «Фенотип» – это то, что находится под влиянием генов. В значительной степени он говорит все о теле. Но есть тонкости значения, которые вытекают из этимологии слова. Phaino в переводе с греческого – «показать», «обнародовать», «выявить», «обнаружить»,
Рассказ Миксотрихи
Рассказ Миксотрихи Mixotricha paradoxa означает «парадоксальный организм с разными жгутиками», и через мгновение мы увидим почему. Это микроорганизм, который живет в кишечнике австралийского термита, «термита Дарвина», Mastotermes darwiniensis. Приятно, хотя не обязательно для местных
Рассказ Taq
Рассказ Taq Достигнув нашего самого древнего свидания, собрав в нашем странствии всю жизнь, которую знаем, мы имеем возможность рассмотреть ее разнообразие. На самом глубинном уровне разнообразие жизни является химическим. Профессии, которыми заняты наши
СТРОИТЕЛЬНАЯ "РАБОТА" БОБРА
СТРОИТЕЛЬНАЯ "РАБОТА" БОБРА В ряде районов СССР сохранился, успешно расселяется и размножается замечательный зверь - речной бобр. Бобры живут иногда скрытно, подобно выдре, в береговых норах. Но местами встречаются и шалашеобразные, искусно сделанные жилища бобров -
Рассказ Stw 573
Рассказ Stw 573 Не думаю, что есть смысл придумывать причины, в силу которых ходить на двух ногах – это здорово. Будь так, шимпанзе делали бы то же самое, не говоря уже о других животных. Нет причины, по которой бег на двух или четырех конечностях быстрее или удобнее.
Рассказ Орангутана
Рассказ Орангутана Возможно, заявление о наших давних связях с Африкой было поспешным. Что если наши предки покинули Африку около 20 млн лет назад и поселились в Азии, а 10 млн лет назад вернулись в Африку?Если так, то современные человекообразные обезьяны, включая тех,
Рассказ Гиббона[12]
Рассказ Гиббона[12] На рандеву № 4 мы встречаем крупную группу пилигримов. И теперь могут возникнуть проблемы с установлением родства. (Чем дальше, тем затруднительнее это сделать.) Существует двенадцать видов гиббонов, принадлежащих к четырем основным группам. Это Bunopithecus
Рассказ Айе-айе
Рассказ Айе-айе Один британский политик как-то описал конкурента (который позднее стал лидером партии) как человека, “в котором есть что-то ночное”. При взгляде на айе-айе складывается именно такое впечатление, и это не случайно: она ведет целиком ночной образ жизни. Из
Рассказ Шерстокрыла
Рассказ Шерстокрыла Шерстокрыл из Юго-Восточной Азии мог бы рассказать нам, каково это – парить во воздуху ночного леса. Но для нас, пилигримов, у него приготовлена гораздо более приземленная история. Мораль ее в том, что составленная нами схема с сопредками, рандеву и
Рассказ Мыши
Рассказ Мыши Из тысяч грызунов домашняя мышь (Mus musculus) может поведать кое-что особенное: ни один другой вид млекопитающих, кроме нашего собственного, мы не изучали так интенсивно. Именно на мышах, а не на пресловутых морских свинках держатся медицинские, физиологические и
Рассказ Гиппопотама
Рассказ Гиппопотама В школе, изучая греческий, я узнал, что “гиппос” означает “лошадь”, а “потамос” – “река”. Выходило, что гиппопотамы – “речные лошади”. Позднее, забросив греческий и занявшись зоологией, я узнал, что гиппопотамы не имеют отношения к лошадям. В
Рассказ Тюленя
Рассказ Тюленя В большинстве естественных популяций число самцов и самок примерно одинаково. На то есть эволюционные причины, и о них рассказал замечательный специалист по статистике и эволюционной генетике Рональд Э. Фишер. Представьте себе популяцию, количество в
Рассказ Броненосца
Рассказ Броненосца С зоологической точки зрения Южная Америка сродни Мадагаскару: как и Мадагаскар, она откололась от Африки – но не с восточной, а с западной стороны. Произошло это примерно тогда же или чуть позднее. Как и Мадагаскар, в течение почти всего времени
Рассказ Утконоса
Рассказ Утконоса Старое латинское название утконоса – Ornithorhynchus paradoxus [“парадоксальный птицеклюв”]. Когда его обнаружили, он показался ученым настолько нелепым, что присланный в музей экземпляр сочли чучелом из сшитых вместе частей тела млекопитающего и птицы. Кое-кто
Рассказ Taq
Рассказ Taq Итак, мы встретили практически все существующие формы жизни. Теперь можно окинуть взглядом открывшееся разнообразие. На самом глубоком уровне разнообразие жизни является химическим. Профессии, которыми владеют наши пилигримы, охватывают широкий диапазон