Рассказ Орангутана
Рассказ Орангутана
Возможно, заявление о наших давних связях с Африкой было поспешным. Что если наши предки покинули Африку около 20 млн лет назад и поселились в Азии, а 10 млн лет назад вернулись в Африку?
Если так, то современные человекообразные обезьяны, включая тех, которые в итоге оказались в Африке, происходят от линии, мигрировавшей из Африки в Азию. Гиббоны и орангутаны – потомки эмигрантов, оставшихся в Азии. Поздние их потомки вернулись в Африку, где раннемиоценовые человекообразные обезьяны уже вымерли, и уже там породили горилл, шимпанзе, бонобо – и нас.
С этим вполне согласуются данные о дрейфе континентов и колебаниях уровня моря. Как раз в нужные моменты в Аравийском море имелись перешейки, соединявшие Азию с Африкой. В пользу этой теории говорит и принцип парсимонии, то есть экономии: хороша та теория, которая многое объясняет, постулируя малое. (Как я не раз говорил, по этому критерию теория естественного отбора Дарвина является, пожалуй, лучшей на свете.) В данном случае речь о том, чтобы до минимума снизить число предполагаемых миграционных событий. С этой точки зрения теория о том, что наши предки все время обитали в Африке (без миграций), на первый взгляд экономнее теории о том, что наши предки сначала переселились из Африки в Азию (первая миграция), а потом вернулись в Африку (вторая миграция).
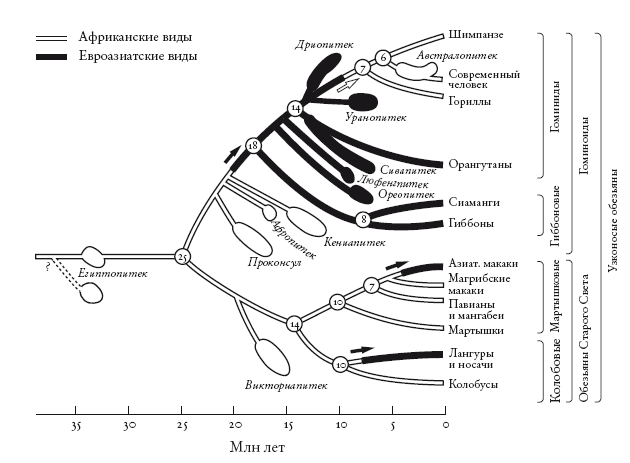
Филогенетическое древо африканских и азиатских обезьян. Расширения обозначают даты, полученные на основе анализа ископаемых, а линии, соединяющие их с древом, построены методом парсимонии. Stewart and Disotell [273].
Но это частный случай: мы рассмотрели лишь собственную линию и проигнорировали остальных человекообразных обезьян, особенно многие ископаемые виды. Стюарт и Дисотелл провели пересчет миграционных событий, в том числе необходимых для объяснения распределения человекообразных обезьян, включая вымерших. Для этого нужно сначала построить древо, отметив на нем все виды, о которых достаточно данных. Следующий шаг – указать для каждого вида, жил он в Африке или в Азии. На диаграмме Стюарта и Дисотелла азиатские ископаемые отмечены черным цветом, африканские – белым. Там представлены не все известные ископаемые, а лишь те, для которых можно определить положение. Стюарт и Дисотелл не забыли и обезьян Старого Света, которые отделились от человекообразных около 25 млн лет назад (самое наглядное различие между ними состоит в том, что первые сохранили хвосты). Миграционные события указаны стрелками.
С учетом этих ископаемых теория переселения в Азию и обратно оказывается экономнее теории о том, что наши предки все время жили в Африке. Оставим хвостатых обезьян, которые, согласно обеим теориям, участвовали в двух волнах миграции из Африки в Азию, и рассмотрим человекообразных. Теория переселения требует всего двух волн миграции:
1. Популяция человекообразных обезьян переселилась из Африки в Азию около 20 млн лет назад. От нее произошли все азиатские человекообразные обезьян, включая современных гиббонов и орангутанов.
2. Популяция человекообразных обезьян вернулась в Африку. От нее произошли современные африканские человекообразные обезьяны, включая нас.
При этом теория постоянного обитания в Африке для объяснения расселения человекообразных обезьян требует шести миграций. И все они направлены из Африки в Азию:
1. Гиббоны (около 18 млн лет назад).
2. Ореопитек (около 16 млн лет назад).
3. Люфенгпитек (около 15 млн лет назад).
4. Сивапитек и орангутан (около 14 млн лет назад).
5. Дриопитек (около 13 млн лет назад).
6. Уранопитек (около 12 млн лет назад).
Разумеется, эти подсчеты верны лишь при условии, что Огюарт и Дис-отелл, опиравшиеся на анатомические данные, верно построили древо. Они, например, считают, что ближайший из всех ископаемых родственник современных африканских человекообразных обезьян – это уранопитек. Его линия ответвляется непосредственно перед линией африканских человекообразных обезьян. Все следующие родственники, согласно оценкам Стюарт и Дисотелла, – азиаты (дриопитек, сивапитек и другие). Если же авторы неверно интерпретировали анатомические данные (например, если ближайший родственник современных африканских человекообразных обезьян – на самом деле ископаемый африканский кениапитек), подсчет миграционных событий надо провести заново.
Сама генеалогическая схема тоже построена по принципу парсимонии. Но мы не пытаемся уменьшить число миграционных событий, необходимых для теории, и игнорируем географию. Вместо этого мы хотим минимизировать число анатомических совпадений (конвергентная эволюция), положенных в основу теории. Так мы получаем древо, не учитывающее географические события, и, чтобы подсчитать миграционные события, накладываем географические данные (черная и белая маркировка на диаграмме). И приходим к выводу: с наибольшей вероятностью современные африканские человекообразные обезьяны – гориллы, шимпанзе и люди – пришли из Азии.
А теперь любопытное замечание. Ричард Клейн из Стэнфордского университета в учебнике по эволюции человека приводит обзор анатомических данных основных ископаемых. Клейн, сравнивая азиатского уранопитека и африканского кениапитека, задается вопросом, кто из них сильнее похож на нашего близкого предка (или родственника) австралопитека. И делает вывод: австралопитек сильнее похож на уранопитека, и если бы уранопитек жил в Африке, он вполне мог быть предком человека. Однако “на основе географических и морфологических данных”, пишет Клейн, кениапитек более подходящий кандидат. Видите? Клейн предполагает, что африканские человекообразные обезьяны просто не могли произойти от азиатского предка, даже если анатомические данные указывают на это. Таким образом, он подсознательно предпочитает географическую парсимонию анатомической. Из соображений анатомической парсимонии уранопитек к нам ближе, чем кениапитек. Однако, хотя об этом не говорится открыто, географическая парсимония берет верх. Стюарт и Дисотелл утверждают, что при рассмотрении географического распространения всех известных ископаемых анатомическая и географическая парсимонии согласуются. То есть географические данные не противоречат утверждению Клейна, на основе анатомических данных признавшему: уранопитек ближе к австралопитеку.
Возможно, этот спор далек от разрешения. Не так-то просто манипулировать анатомической и географической парсимониями. Статья Стюарт и Дисотелла вызвала бурную реакцию: в научных журналах появлялись и восторженные, и разгромные отклики. Мне кажется, что сегодня, оценив все доступные данные, мы должны отдать предпочтение теории “переселения в Азию и обратно”. Два миграционных события – меньше, чем шесть. Кроме того, мне кажется, между позднемиоценовыми азиатскими обезьянами и “нашей” линией африканских обезьян (например австралопитеками и шимпанзе) есть немалое сходство. Таким образом, теория представляет собой компромисс. Исходя из этого, я назначаю рандеву № 3 (и № 4) не в Африке, а в Азии.
Мораль “Рассказа Орангутана” неоднозначна. Когда ученый выбирает между двумя теориями, на первом месте всегда принцип парсимонии. Однако не всегда ясно, как ее оценивать. Наличие хорошей родословной бывает необходимо для дальнейших рассуждений в рамках эволюционной теории. Но чтобы построить хорошее древо, нужен опыт.
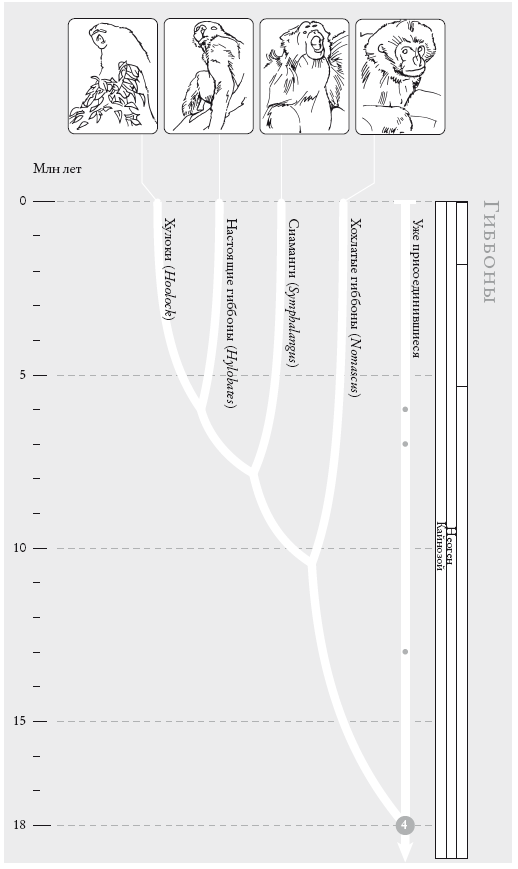
ГИББОНЫ. Двенадцать видов гиббонов сейчас объединяют в четыре группы. Порядок ветвления внутри них остается спорным (см. “Рассказ Гиббона”).
На рис. (слева направо): восточный хулок (Hoolock leuconedys), быстрый гиббон (Hylobatesagilis), сиаманг (Symphalangus syndactylus), желтощекий номаск (Nomascus gabriellae).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Рассказ Евы
Рассказ Евы Есть впечатляющая разница между «родословными деревьями генов» и «родословными деревьями людей». В отличие от человека, который происходит от двух родителей, у гена есть только один родитель. Каждый из Ваших генов должен был быть получен или от Вашей матери,
Рассказ Секвойи
Рассказ Секвойи Люди спорят о том, какое одно место в мире Вы должны посетить прежде, чем умрете. Мой кандидат – лес Muir Woods, несколько севернее моста «Золотые Ворота». Или, если Вы считаете, что слишком поздно, я не могу вообразить лучшего места, чтобы быть похороненным (вот
Рассказ Миксотрихи
Рассказ Миксотрихи Mixotricha paradoxa означает «парадоксальный организм с разными жгутиками», и через мгновение мы увидим почему. Это микроорганизм, который живет в кишечнике австралийского термита, «термита Дарвина», Mastotermes darwiniensis. Приятно, хотя не обязательно для местных
Рассказ Taq
Рассказ Taq Достигнув нашего самого древнего свидания, собрав в нашем странствии всю жизнь, которую знаем, мы имеем возможность рассмотреть ее разнообразие. На самом глубинном уровне разнообразие жизни является химическим. Профессии, которыми заняты наши
Рассказ Земледельца
Рассказ Земледельца Аграрная революция началась около 10 тыс. лет назад, на исходе последнего ледникового периода, между Тигром и Евфратом – в регионе, прозванном “плодородным полумесяцем”. Другие очаги земледелия возникли (скорее всего, независимо) в Китае, долине Нила
Рассказ Stw 573
Рассказ Stw 573 Не думаю, что есть смысл придумывать причины, в силу которых ходить на двух ногах – это здорово. Будь так, шимпанзе делали бы то же самое, не говоря уже о других животных. Нет причины, по которой бег на двух или четырех конечностях быстрее или удобнее.
Рассказ Орангутана
Рассказ Орангутана Возможно, заявление о наших давних связях с Африкой было поспешным. Что если наши предки покинули Африку около 20 млн лет назад и поселились в Азии, а 10 млн лет назад вернулись в Африку?Если так, то современные человекообразные обезьяны, включая тех,
Рассказ Айе-айе
Рассказ Айе-айе Один британский политик как-то описал конкурента (который позднее стал лидером партии) как человека, “в котором есть что-то ночное”. При взгляде на айе-айе складывается именно такое впечатление, и это не случайно: она ведет целиком ночной образ жизни. Из
Рассказ Шерстокрыла
Рассказ Шерстокрыла Шерстокрыл из Юго-Восточной Азии мог бы рассказать нам, каково это – парить во воздуху ночного леса. Но для нас, пилигримов, у него приготовлена гораздо более приземленная история. Мораль ее в том, что составленная нами схема с сопредками, рандеву и
Рассказ Мыши
Рассказ Мыши Из тысяч грызунов домашняя мышь (Mus musculus) может поведать кое-что особенное: ни один другой вид млекопитающих, кроме нашего собственного, мы не изучали так интенсивно. Именно на мышах, а не на пресловутых морских свинках держатся медицинские, физиологические и
Рассказ Бобра
Рассказ Бобра “Фенотип” – это то, что находится под влиянием генов. В целом под фенотипом понимают все, что относится к телу. Но есть тонкости, вытекающие из этимологии. Phamo по-гречески означает “показывать”, “обнаруживать”, “выявлять”, “выражать”, “раскрывать”.
Рассказ Гиппопотама
Рассказ Гиппопотама В школе, изучая греческий, я узнал, что “гиппос” означает “лошадь”, а “потамос” – “река”. Выходило, что гиппопотамы – “речные лошади”. Позднее, забросив греческий и занявшись зоологией, я узнал, что гиппопотамы не имеют отношения к лошадям. В
Рассказ Тюленя
Рассказ Тюленя В большинстве естественных популяций число самцов и самок примерно одинаково. На то есть эволюционные причины, и о них рассказал замечательный специалист по статистике и эволюционной генетике Рональд Э. Фишер. Представьте себе популяцию, количество в
Рассказ Броненосца
Рассказ Броненосца С зоологической точки зрения Южная Америка сродни Мадагаскару: как и Мадагаскар, она откололась от Африки – но не с восточной, а с западной стороны. Произошло это примерно тогда же или чуть позднее. Как и Мадагаскар, в течение почти всего времени
Рассказ Утконоса
Рассказ Утконоса Старое латинское название утконоса – Ornithorhynchus paradoxus [“парадоксальный птицеклюв”]. Когда его обнаружили, он показался ученым настолько нелепым, что присланный в музей экземпляр сочли чучелом из сшитых вместе частей тела млекопитающего и птицы. Кое-кто
Рассказ Taq
Рассказ Taq Итак, мы встретили практически все существующие формы жизни. Теперь можно окинуть взглядом открывшееся разнообразие. На самом глубоком уровне разнообразие жизни является химическим. Профессии, которыми владеют наши пилигримы, охватывают широкий диапазон