Дрейф и отбор: кто кого?
Дрейф и отбор: кто кого?
Генетический дрейф царствует над нейтральными мутациями (аллелями), отбор — над полезными и вредными. Отбор, повышающий частоту полезных мутаций, называют положительным. Отбор, отбраковывающий вредные мутации, — отрицательным, или очищающим.
Впрочем, все так хорошо и просто только в больших популяциях. В маленьких ситуация сложнее, потому что отбор и дрейф — две главные движущие силы эволюции — начинают конкурировать друг с другом за контроль над слабовредными и слабополезными мутациями.
Посмотрим, как это происходит. Воспользуемся для этого той же моделью с мюмзиками. Изменим только характер мутации, которая привела к появлению аллеля А2. До сих пор мы считали, что мутация была нейтральной. Поскольку она была нейтральной, обладатели аллелей А1 и А2 имели одинаковую плодовитость. Давайте теперь предположим, что мутация была полезной, что она повысила плодовитость мюмзиков на 5 %. Это можно смоделировать так: пусть мюмзики с генотипом А1 рожают по 20 детенышей, а мюмзики с генотипом А2 — по 21. Начальная частота аллеля А2 пусть будет по-прежнему равна 0,5. Только теперь мы рассмотрим популяции с разной численностью (N).
На рисунке [см. ниже] показано, как будет меняться частота А2 в том случае, если его носители имеют пятипроцентное адаптивное преимущество. Мы видим, что в большой популяции (N = 5000) частота А2 неуклонно растет, приближаясь к единице. Так работает отбор в идеальных для него условиях, т. е. в больших популяциях, где влияние дрейфа на полезные и вредные аллели пренебрежимо мало. Форма у кривой довольно правильная, и это наводит на мысль, что ее можно описать какой-нибудь математической формулой. Это действительно так, но выводить формулу мы не будем, чтобы не утомить читателей (а любители математики могут сделать это самостоятельно)[19].
Перед нами, между прочим, важнейший эволюционный процесс — аллельное замещение, т. е. вытеснение более приспособленным аллелем менее приспособленного. Процесс идет не слишком быстро. В большой популяции, например состоящей из миллиона особей, для того чтобы зафиксировалась новая полезная мутация, дающая 5-процентное адаптивное преимущество, требуется около 560 поколений. А ведь 5 % — это серьезное преимущество. Такие мутации — редкость. Ждать, пока зафиксируется мутация с преимуществом в 1 %, придется уже 2800 поколений! Тем не менее в большой популяции отбор «чувствует» даже самую незначительную разницу в приспособленности. Это обеспечивает эффективную (пусть и медленную) фиксацию слабополезных мутаций и элиминацию слабовредных[20].
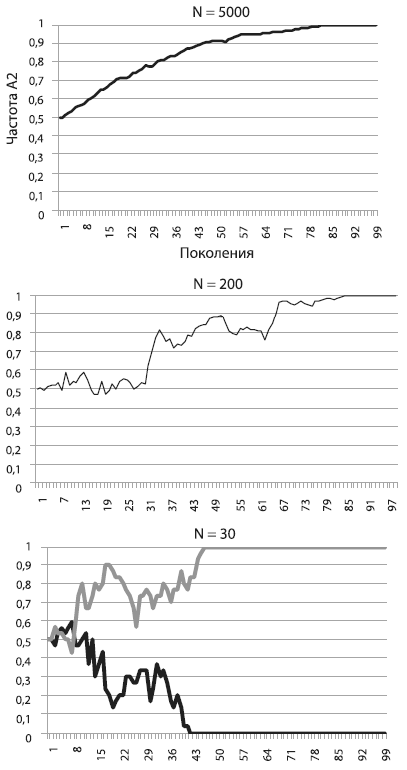
На среднем графике мы видим, какая судьба ждет такую же полезную мутацию, повышающую приспособленность на 5 %, в популяции меньшего размера (N = 200). Частота полезного аллеля и в этом случае росла и в конце концов достигла 1 (мутация зафиксировалась), но путь был труден и извилист. Пожалуй, нашей мутации могло и не повезти, ее шансы на фиксацию не были стопроцентными.
Наконец, на правом графике мы видим, что происходит с точно таким же полезным аллелем в совсем крошечной популяции (N = 30). На рисунке показаны результаты двух запусков модели. В одном случае мутация зафиксировалась, в другом — элиминировалась. Не правда ли, картинка похожа на результат работы дрейфа, а не отбора?
Так оно и есть. В этом заключается главный урок, который мы можем извлечь из наших экспериментов. Чем меньше популяция, тем слабее в ней власть отбора и тем могущественнее дрейф. В маленьких популяциях слабополезные и слабовредные мутации начинают вести себя фактически как нейтральные. Их частоты «случайно блуждают», пока не наткнутся на верхний или нижний порог. В маленьких популяциях, где царствует дрейф, небольшие различия в приспособленности становятся невидимыми для отбора. Поэтому слабовредная мутация запросто может зафиксироваться, а слабополезная — элиминироваться.
Хорошо это или плохо? В большинстве случаев, конечно, плохо. Потеря слабополезных мутаций мешает маленькой популяции приспосабливаться к меняющимся условиям. Бесконтрольное накопление слабовредных мутаций и вовсе может поставить ее на грань вымирания. Именно в этом, между прочим, видят биологи одну из причин того, что крупные животные вымирают в среднем чаще, чем мелкие. У крупных животных, таких как носороги или слоны, популяции не могут быть такими же большими, как у мышей или насекомых. Это снижает приспособляемость крупных животных.
Но у этой медали есть и обратная сторона. У маленьких популяций больше шансов выйти из «ловушки локального оптимума», т. е. сползти с невысокого пика ландшафта приспособленности и взобраться на другой, повыше. Ведь отбор гонит организмы вверх и только вверх. Если популяция велика и малейшее различие в приспособленности «заметно» для отбора, спуск по склонам становится невозможен. Однажды взобравшись на одинокий холм, большая популяция уже никогда с него не слезет. Что касается дрейфа, то он ведет организмы по ландшафту приспособленности хаотическим образом, не замечая подъемов и спусков. Если популяция невелика и дрейф силен, у организмов есть шанс иногда двигаться не только вверх, но и немного вниз (и в сторону). Спустившись в ложбинку, организмы могут «обнаружить», что отсюда есть другой, более перспективный подъем. Если, конечно, не вымрут раньше, чем на него наткнутся.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава 10 Следы, оставленные три с половиной миллиона лет назад! Дарт, Брум и современные исследователи. Дрейф континентов. Поименный список гоминидов. Люси и ее сородичи.
Глава 10 Следы, оставленные три с половиной миллиона лет назад! Дарт, Брум и современные исследователи. Дрейф континентов. Поименный список гоминидов. Люси и ее сородичи. Столь длительная сохранность доисторических следов у Лаетоли – случай фантастический, но не
ГЛАВА 3 Эволюция земной коры. Дрейф континентов и спрединг океанического дна. Мантийная конвекция
ГЛАВА 3 Эволюция земной коры. Дрейф континентов и спрединг океанического дна. Мантийная конвекция Горные породы, формирующие кору Земли, как мы помним, бывают изверженные — первичные, образовавшиеся при охлаждении и затвердевании магмы, и осадочные — вторичные,
Нейтральные мутации и генетический дрейф — движение без правил
Нейтральные мутации и генетический дрейф — движение без правил Ландшафт приспособленности — образ яркий и полезный, но, как и всякая модель, он несовершенен. Многие аспекты эволюционного процесса с его помощью отразить трудно или невозможно. Реальный ландшафт
Кого мы ищем?
Кого мы ищем? Во всей Сибири нет озера, похожего на чукотское озеро Эльгыгытгын, имеющего идеальную круглую форму. Озеро было открыто лишь в 1933 году. О нем местные жители сложили много легенд. В одной говорится, что в озере обитает огромное чудовище Калилгу со страшной
ДРЕЙФ ГЕНОВ
ДРЕЙФ ГЕНОВ Иногда эта концепция называется «эффект Сьюэлла — Райта», в честь предложивших ее двух популяционных генетиков. После того как Мендель доказал, что гены являются единицами наследственности, а Харди и Вайнберг продемонстрировали механизм их поведения,
ДРЕЙФ МАТЕРИКОВ
ДРЕЙФ МАТЕРИКОВ В 1912 году немецкий ученый Альфред Вегенер предположил, что около 200 миллионов лет назад все материки Земли составляли единый массив суши, который он назвал Пангеей. В последующие 200 миллионов лет Пангея разделилась на несколько материков, которые стали
Кого мы выбираем?
Кого мы выбираем? Где-то в ожиданье одиноком Томится одинокая душа, Но ждет она, надеется, — и к сроку С другой душою встретится, спеша. И воедино, словно два потока, Сольются, тихой нежностью дыша, И вечность распахнет свои врата, И тьма уж не вернется никогда. (1) Эдвин
Кого выбирают мужчины
Кого выбирают мужчины В ходе исследования, охватившего около 10 000 участников из 37 стран, ученые просили мужчин и женщин распределить 18 характеристик в порядке убывания их важности при выборе супруга. (29) На первое место представители обоих полов поставили любовь и
Кого выбирают женщины?
Кого выбирают женщины? Изучение 800 частных объявлений о знакомствах, размещенных в американских газетах и журналах, показало: в США женщины вдвое чаще, чем мужчины, ищут партнеров, способных помочь им материально. (48) Множество женщин-врачей, женщин-юристов, просто
Так кого же больше?
Так кого же больше? Если у тебя нет женщины, значит, у кого-то их две. Аркадий Давидович В бытовом обсуждении проблемы женского одиночества часто звучит мнение, что в означенном одиночестве виновата нехватка мужчин. И хотя строгие социологические исследования не
У кого учиться?
У кого учиться? Учиться всегда, всему и у всех бесполезно. Нужно знать, когда, чему и у кого учиться. Это знание и содержит программа импринтинга. Животные обучаются сами, обучаются в играх со сверстниками, обучаются у родителей и обучаются у взрослых. Программа такова, что
У кого где уши
У кого где уши У кузнечика и сверчка — в передних ножках; у саранчи уши в брюшке, в том месте, откуда растут задние ноги-ходули; у бабочек и мух — в основании крыльев; у мотыльков — в конце груди, начале брюшка; у муравьев, по-видимому, — в усиках; у некоторых жуков — тоже; у
ОТ КОГО ПРОИЗОШЛИ ДИНОЗАВРЫ?
ОТ КОГО ПРОИЗОШЛИ ДИНОЗАВРЫ? В последнее время часто можно слышать вопрос, уже ставший риторическим: отчего вымерли динозавры? И при всем разнообразии ответов почему-то совсем не звучит другой вопрос: а откуда же взялись на Земле эти самые динозавры?Ну это же скучно и
КТО ОТ КОГО «ПРОИЗОШЁЛ»?
КТО ОТ КОГО «ПРОИЗОШЁЛ»? Одно важное обстоятельство не должно уйти из поля нашего внимания: чтобы приспосабливаться, надо иметь тело, т. е. то, что будет приспосабливаться. Например, человеку тело дано отцом и матерью, у которых также были отец и мать, имевшие родителей.
Кто кого перехитрит
Кто кого перехитрит Мир полового размножения переменчив. Из-за наличия паразитов хозяину выгодно в каждом поколении генетически изменяться — делать именно то, к чему оно и приводит. Успешность генных комбинаций, защитивших родителей от паразитов — главная причина для
Кого больше?
Кого больше? В настоящее время генетиками и молекулярными биологами сделан огромный рывок вперед, значительно обогативший наши знания о процессах, лежащих в основе формирования мужского и женского организма. Но природа пола продолжает таить в себе массу загадок. Отчего,