Рассказ Миноги
Рассказ Миноги
Причина, по которой миноге выпало рассказывать этот рассказ, будет выявлена в конце. Это – повторение темы, с которой мы сталкивались прежде: есть отдельный взгляд на происхождение и родословную с точки зрения гена, удивительно независимый от представления, которое мы получаем, когда думаем о генеалогических деревьях более традиционным способом.
Отдельный взгляд на родословную с точки зрения гена.
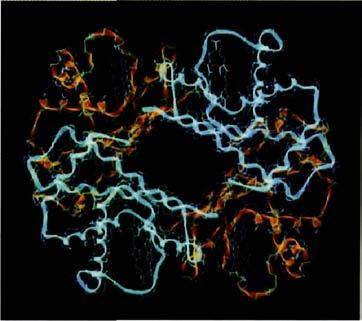
Компьютерное графическое представление молекулы гемоглобина, показывающее две альфа-цепи глобина синим цветом и две бета-цепи глобина желтым.
Гемоглобин известен как жизненно важная молекула, которая несет кислород к нашим тканям и придает нашей крови ее захватывающий цвет. Человеческий взрослый гемоглобин – в действительности соединение четырех цепей белка, названных глобинами, закрученных друг вокруг друга. Их последовательности ДНК показывают, что четыре цепи глобина близко связаны друг с другом, но не идентичны. Две из них называют альфа-глобинами (каждая соединяет цепочкой 141 аминокислоту), и две – бета-глобинами (каждая соединяет 146 аминокислот). Гены, кодирующие альфа-глобины, находятся на нашей 11 хромосоме; а кодирующие бета-глобины – находятся на хромосоме 16. На каждой из этих хромосом располагается подряд кластер генов глобинов вперемежку с некоторым количеством мусорной ДНК, которая никогда не транскрибируется. Альфа-кластер на хромосоме 11 содержит семь генов глобина. Четыре из них – псевдогены – выведенные из строя версии альфы с ошибками в их последовательности, никогда не переводимые в белок. Два истинных альфа-глобина используются во взрослом возрасте. Последний глобин называют дзэтой, и он используется только в эмбрионах. В бета кластере на хромосоме 16 есть шесть генов, некоторые из которых повреждены, а один используется только в эмбрионе. Гемоглобин взрослого, как мы видели, содержит две альфа и две бета цепи, обвитые друг вокруг друга, чтобы сформировать красиво функционирующий узел.
Не обращайте внимания на все эти сложности. Вот очаровательный момент. Тщательный побуквенный анализ показывает, что различные виды генов глобина – буквально кузены друг друга, члены семьи. Но эти дальние родственники все еще сосуществуют в Вас и во мне. Они все еще сидят бок о бок со своими кузенами в каждой клетке каждого бородавочника и каждого вомбата, каждой совы и каждой ящерицы.
В масштабе целых организмов, конечно, все позвоночные животные – также кузены друг друга. Дерево эволюции позвоночных – генеалогическое дерево, с которым мы все знакомы, его точки ветвления представляют события видообразования – раскол видов на дочерние виды. В обратном направлении они – пункты свиданий, которые перемежают это странствие. Но есть другое генеалогическое дерево, занимающее ту же шкалу времени, ветви которого представляют не события видообразования, а события дупликации гена в геномах. И образец ветвления дерева глобина выглядит очень отличающимся от образца ветвления генеалогического дерева, если мы чертим его обычным, ортодоксальным способом, с видами, ветвящимися, чтобы сформировать дочерние виды. Не существует единственного эволюционного дерева, в котором виды делятся и дают начало дочерним видам. У каждого гена есть свое собственное дерево, своя собственная хроника расщеплений, свой собственный перечень близких и дальних родственников.
Около дюжины различных глобинов в Вас и во мне перешли к нам через все родословные наших позвоночных предков. Приблизительно пятьсот миллионов лет назад у бесчелюстных рыб, возможно, похожих на миногу, предковый ген глобина случайно раскололся на два, обе копии остались в различных частях генома той рыбы. Потом было две копии этого гена, в различных частях генома всех потомков животного. Одной копии было предопределено дать начало альфа-группе на том, что, в конечном счете, станет хромосомой 11 в нашем геноме, другой – бета группе на нашей теперешней хромосоме 16. Нет никакого смысла пытаться догадаться на какой хромосоме любой из них располагался у промежуточных предков. Местоположения опознаваемых последовательностей ДНК, фактически ряд хромосом, на которые разделен геном, перетасованы и изменены с удивительно беспечной энергией. Системы нумерации хромосом, поэтому, не являются общими для всех групп животных.
Прошли века, были дальнейшие дупликации, и, несомненно, также некоторые делеции. Приблизительно 400 миллионов лет назад предковый альфа-ген был дуплицирован снова, но на сей раз две копии оставались близкими соседями друг друга в кластере на одной и той же хромосоме. Одной из них было суждено стать дзэтой наших эмбрионов, другая стала альфа-генами глобина взрослых людей (дальнейшие ветвления дали начало атрофированным псевдогенам, о которых я упоминал). Подобная история произошла с бета-ветвью, но с дупликациями в другие моменты геологической истории.
Теперь интересный момент. Учитывая, что раскол между альфа- и бета-группой имел место пятьсот миллионов лет назад, конечно, не только наши, человеческие геномы свидетельствуют о расколе и обладают как альфа-, так и бета-генами в различных своих частях. Мы должны ожидать увидеть тот же раскол, если посмотрим на геномы любых других млекопитающих, птиц, рептилий, амфибий или костных рыб – до нашего общего со всеми ими предка, жившего меньше чем 500 миллионов лет назад. Везде, где он был исследован, это ожидание оказалось правильным. Наша самая большая надежда на обнаружение позвоночного животного, которое не разделяет с нами древний альфа/бета раскол, было бы бесчелюстной рыбой, такой как минога или миксина, поскольку они – наши самые отдаленные кузены среди сохранившихся позвоночных животных. Они – единственные сохранившиеся позвоночные животные, чей общий предок с остальными является достаточно древним, чтобы он мог предшествовать альфа/бета расколу. Без сомнения, эти бесчелюстные рыбы – единственные известные позвоночные животные, у которых отсутствует альфа/бета разделение. Другими словами, Свидание 22 является настолько древним, что оно предшествовало расколу между альфа- и бета-глобином.
Нечто подобное «Рассказу Миноги» могло быть сказано для каждого из наших генов, поскольку все они, если вернуться достаточно далеко, обязаны своим происхождением расколу некоторого древнего гена. И нечто подобное этой всей книге могло быть написано для каждого гена. Мы произвольно выбрали, что это должно быть человеческим путешествием, и мы определили свои вехи как пункты свиданий с другими линиями, что означает в прямом направлении события видообразования, в которых наши человеческие предки откалываются от других. Я уже отмечал, что мы могли точно так же начать свое путешествие с современного дюгоня или современного черного дрозда и перечислить другой ряд копредков на пути назад к Кентербери. Но я теперь выскажу более радикальное соображение. Мы могли также описать обратное странствие для любого гена.
Мы могли бы проследить за странствием альфа-гемоглобина, или цитохромома-c, или любого другого упомянутого гена. Свидание 1 было бы вехой, в которой наш выбранный ген последний раз был дуплицирован, создав свою копию в другом месте в геноме. Свидание 2 было бы предыдущим случаем дупликации, и так далее. Каждое из свиданий происходило бы в некотором отдельном животном или растении, так же как «Рассказ Миноги» определил кембрийскую бесчелюстную рыбу как вероятный сосуд, где произошел раскол между бета- и альфа- гемоглобином.
Точка зрения гена на эволюцию продолжает привлекать наше внимание.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Миграции каспийской миноги
Миграции каспийской миноги В 1911–1912 годах я подробно проследил миграцию каспийской миноги. До того времени такие наблюдения над этой чрезвычайно скрытной рыбой не производились. Оказалось, что минога начинает входить из моря в реку осенью при температуре воды 10–11° и
Рассказ Евы
Рассказ Евы Есть впечатляющая разница между «родословными деревьями генов» и «родословными деревьями людей». В отличие от человека, который происходит от двух родителей, у гена есть только один родитель. Каждый из Ваших генов должен был быть получен или от Вашей матери,
Рассказ Миксотрихи
Рассказ Миксотрихи Mixotricha paradoxa означает «парадоксальный организм с разными жгутиками», и через мгновение мы увидим почему. Это микроорганизм, который живет в кишечнике австралийского термита, «термита Дарвина», Mastotermes darwiniensis. Приятно, хотя не обязательно для местных
Рассказ Taq
Рассказ Taq Достигнув нашего самого древнего свидания, собрав в нашем странствии всю жизнь, которую знаем, мы имеем возможность рассмотреть ее разнообразие. На самом глубинном уровне разнообразие жизни является химическим. Профессии, которыми заняты наши
Рассказ Stw 573
Рассказ Stw 573 Не думаю, что есть смысл придумывать причины, в силу которых ходить на двух ногах – это здорово. Будь так, шимпанзе делали бы то же самое, не говоря уже о других животных. Нет причины, по которой бег на двух или четырех конечностях быстрее или удобнее.
Рассказ Айе-айе
Рассказ Айе-айе Один британский политик как-то описал конкурента (который позднее стал лидером партии) как человека, “в котором есть что-то ночное”. При взгляде на айе-айе складывается именно такое впечатление, и это не случайно: она ведет целиком ночной образ жизни. Из
Рассказ Шерстокрыла
Рассказ Шерстокрыла Шерстокрыл из Юго-Восточной Азии мог бы рассказать нам, каково это – парить во воздуху ночного леса. Но для нас, пилигримов, у него приготовлена гораздо более приземленная история. Мораль ее в том, что составленная нами схема с сопредками, рандеву и
Рассказ Мыши
Рассказ Мыши Из тысяч грызунов домашняя мышь (Mus musculus) может поведать кое-что особенное: ни один другой вид млекопитающих, кроме нашего собственного, мы не изучали так интенсивно. Именно на мышах, а не на пресловутых морских свинках держатся медицинские, физиологические и
Рассказ Бобра
Рассказ Бобра “Фенотип” – это то, что находится под влиянием генов. В целом под фенотипом понимают все, что относится к телу. Но есть тонкости, вытекающие из этимологии. Phamo по-гречески означает “показывать”, “обнаруживать”, “выявлять”, “выражать”, “раскрывать”.
Рассказ Гиппопотама
Рассказ Гиппопотама В школе, изучая греческий, я узнал, что “гиппос” означает “лошадь”, а “потамос” – “река”. Выходило, что гиппопотамы – “речные лошади”. Позднее, забросив греческий и занявшись зоологией, я узнал, что гиппопотамы не имеют отношения к лошадям. В
Рассказ Тюленя
Рассказ Тюленя В большинстве естественных популяций число самцов и самок примерно одинаково. На то есть эволюционные причины, и о них рассказал замечательный специалист по статистике и эволюционной генетике Рональд Э. Фишер. Представьте себе популяцию, количество в
Рассказ Броненосца
Рассказ Броненосца С зоологической точки зрения Южная Америка сродни Мадагаскару: как и Мадагаскар, она откололась от Африки – но не с восточной, а с западной стороны. Произошло это примерно тогда же или чуть позднее. Как и Мадагаскар, в течение почти всего времени
Рандеву № 22 Миноги и миксины
Рандеву № 22 Миноги и миксины Рандеву № 22 состоялось в теплых морях раннего кембрия, около 530 млн лет назад. Сопредок № 22, по моей приблизительной оценке, был нашим прародителем в 240-миллионном поколении. Полномочными посланниками первоначальной эпохи позвоночных
Рассказ Миноги
Рассказ Миноги Сейчас речь снова пойдет о том, что генные деревья могут сильно отличаться от генеалогических схем, построенных традиционным способом.Гемоглобин – это жизненно важная молекула, которая переносит кислород к нашим тканям и придает крови яркий цвет.
Рассказ Taq
Рассказ Taq Итак, мы встретили практически все существующие формы жизни. Теперь можно окинуть взглядом открывшееся разнообразие. На самом глубоком уровне разнообразие жизни является химическим. Профессии, которыми владеют наши пилигримы, охватывают широкий диапазон