Происхождение и эволюция человекообразных обезьян
Происхождение и эволюция человекообразных обезьян
Примерно на рубеже олигоцена и миоцена (23 млн лет назад), или чуть раньше (см. рис. 2) происходит разделение дотоле единого ствола узконосых обезьян на две ветви: церкопитекоидов, или собакоподобных (Cercopithecoidea) и гоминоидов, т. е. человекообразных (Hominoidea). Это разделение, по всей видимости, во многом было обусловлено тем, что часть узконосых (предки церкопитекоидов) перешла к питанию листьями, тогда как другая часть (предки гоминоидов) сохранила верность фруктовой диете. Различия в меню сказались, в частности, на строении зубов, что чрезвычайно важно для палеонтологов, поскольку именно зубы составляют бо?льшую часть ископаемых находок. Поверхность жевательных зубов церкопитекоидов имеет характерный, только им присущий узор, образуемый четырьмя бугорками. На зубах же человекообразных обезьян пять округлых бугорков, разделённых У-образной бороздкой — так называемый «узор дриопитека» (рис. 5).
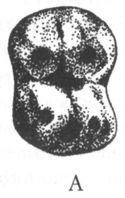

Рис. 5. Поверхность коренных зубов церкопитекоидов (А) и гоминоидов (Б)
Церкопитекоидов, представленных единственным, но очень многочисленным семейством мартышкообразных, часто называют низшими узконосыми обезьянами, а гоминоидов — высшими. Помимо особенностей формы зубов, гоминоидов отличает от низших узконосых обезьян также отсутствие хвоста, более короткое (по отношению к конечностям), плоское и широкое туловище и, наконец, специфическое строение плечевого сочленения, обеспечивающее большую свободу вращения верхних конечностей в разных плоскостях. По-видимому, все перечисленные признаки были приобретены ранними гоминоидами в результате приспособления к таким способам передвижения по деревьям, которые требуют вертикального и хотя бы частично выпрямленного положения тела. Это лазание с опорой на нижние конечности, а также так называемая брахиация, т. е. перенос или перебрасывание тела с ветки на ветку с помощью верхних конечностей (рис. 6). Для низших обезьян ни то, ни другое, в общем, не свойственно, и они, в отличие от человекообразных, даже по ветвям передвигаются, как правило, на четырёх конечностях, подобно всем остальным млекопитающим от белки до леопарда.
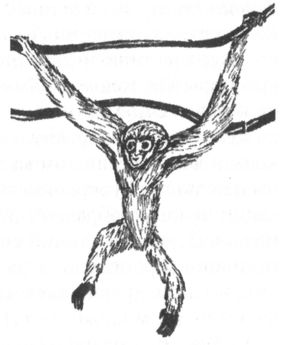
Рис. 6. Гиббоны — классические брахиаторы
Одно время некоторые исследователи полагали, что церкопитекоиды и гоминоиды разделились ещё в раннем олигоцене, и что уже проплиопитеки и египтопитеки, жившие примерно 30–35 млн лет назад, должны рассматриваться как гоминоиды. Действительно, зубы этих обезьян, найденные в Файюмской впадине, несут хорошо выраженный узор дриопитека, но вот кости их черепа и скелета по своему строению ближе к аналогичным костям церкопитекоидов. Такая мозаичность признаков позволяет видеть в этих родах более или менее близкое подобие той предковой формы, от которой произошли церкопитекоиды и гоминоиды. К сожалению, огромный временной интервал, захватывающий весь поздний олигоцен, всё ещё остаётся практически неохарактеризованным ископаемым материалом, и поэтому представить себе сколько-нибудь детально процесс расхождения двух ветвей узконосых обезьян пока невозможно.
Одно время в качестве наиболее ранней формы гоминоидов предположительно рассматривали род камояпитек (Kamoyapithecus), выделенный по находкам на позднеолигоценовом местонахождении Лосидок в северной Кении. Благодаря своему залеганию между двумя хорошо датированными калийаргоновым методом слоями базальта, нижний из которых имеет возраст 27,5±0,3 млн лет, а верхний 24,2±0,3 млн лет, эти находки имеют надёжную хронологическую привязку. Однако они всё же слишком малочисленны и фрагментарны, чтобы их с полной уверенностью можно было определить как останки человекообразной обезьяны. Более представительный материал, проливающий свет на ранние этапы эволюции гоминоидов, происходит с ряда местонахождений на западе Кении, но даже древнейшее из них — Месва Бридж — примерно на 3 млн лет моложе, чем Лосидок.
Сейчас, благодаря находкам в Африке и Евразии, известно около 30 родов миоценовых гоминоидов, но предполагается, что этот материал и наполовину не отражает их действительного разнообразия. По некоторым оценкам, число родов, существовавших в названный период, могло быть в пять раз больше, и те из них, которые имеют критическое значение для понимания филогенетических взаимоотношений разных групп внутри надсемейства человекообразных, ещё не открыты.[3] Так это или нет, но представления о филогенезе гоминоидов — как ископаемых, так и современных — пока и в самом деле далеки от ясности.
С середины 60-х гг. XX в. для построения генеалогического древа отряда приматов (а также многих других групп животных) стали использовать информацию, содержащуюся в макромолекулах белков и особенно нуклеиновых кислот. Принцип, лежащий в основе применяемых для этого методов, отчасти сродни тому, на котором базируются радиоизотопные способы датирования. Если в последних в качестве основы расчётов используется примерно одинаковая для больших промежутков времени скорость распада радиоактивных элементов (например, C14 — радиоактивного углерода), то в первых аналогичную роль играют так называемые нейтральные точечные мутации. Такие мутации, хотя и ведут к изменению последовательности нуклеотидов ДНК, не имеют, как предполагается, значения для естественного отбора и распределяются во времени (конечно, речь идёт о достаточно длительных его отрезках) более или менее равномерно. Если это так, то, сравнивая посредством различных, весьма изощрённых, методик строение молекул ДНК у разных групп организмов, можно судить о степени их родства (чем оно ближе, тем меньше должно быть различий), а при известной скорости мутирования даже и о примерном времени расхождения от общего предка. Конечно, биомолекулярные методы филогенетических исследований нельзя считать абсолютно надёжными и самодостаточными, и в этой области имеется ещё множество нерешённых проблем. Однако, как показывает опыт, в том, что касается эволюции приматов, биомолекулярный и палеонтологический анализ дают в общем, как правило, достаточно близкие результаты.
Сопоставление последовательностей нуклеотидов в молекулах ДНК, взятых у современных церкопитековых и человекообразных обезьян, говорит, по мнению большинства специалистов, о том, что эволюционные пути этих групп разошлись где-то в интервале от 22 до 28 млн лет назад. Таким образом, палеонтологические и молекулярные данные, взятые вместе, позволяют считать, что самостоятельная филогенетическая история надсемейства гоминоидов, в состав которого из ныне живущих приматов входят человек и человекообразные обезьяны (шимпанзе, горилла, орангутанг, гиббон, сиаманг), началась около 25 млн лет назад (рис. 4).
До недавнего времени в составе надсемейства гоминоидов было принято выделять три семейства: гилобатид (Hylobatidae), представленных гиббоном и сиамангом, понгид (Pongidae), к которым относили роды орангутанга (Pongo), гориллы (Gorilla) и шимпанзе (Pan), и гоминид (Hominidae), т. е. человека и его прямоходящих предков. Эта классификация основывалась на внешних анатомических признаках, прежде всего таких, как пропорции конечностей, особенности строения клыков и коренных зубов и т. п. Широкое использование биомолекулярных методов в систематике, однако, показало, что необходима перегруппировка принятых к настоящему времени таксонов. В частности, выяснилось, что орангутанг в генетическом плане отстоит от африканских человекообразных обезьян (горилла и шимпанзе) дальше, чем последние от человека, и должен быть выделен в особое семейство. Кроме того, появились данные, говорящие о том, что генетическая дистанция между человеком и шимпанзе может быть даже меньше, чем между шимпанзе и гориллой, и, если это так, то, значит, необходимы и соответствующие изменения в систематике.
Практически нет сомнений в том, что появились гоминоиды в Африке, и почти 10 млн лет их история оставалась связанной исключительно с этим континентом. Если не считать упоминавшиеся выше спорные материалы из Лосидок, то самые ранние гоминоиды, найденные на нижнемиоценовых местонахождениях Восточной Африки, относятся к роду проконсул (Proconsul) (рис. 7). Правда, существует точка зрения, согласно которой проконсул тоже не являлся ещё собственно гоминоидом, но и её сторонники допускают, что какой-то из видов этого рода вполне мог быть общим предком всех позднейших человекообразных обезьян.
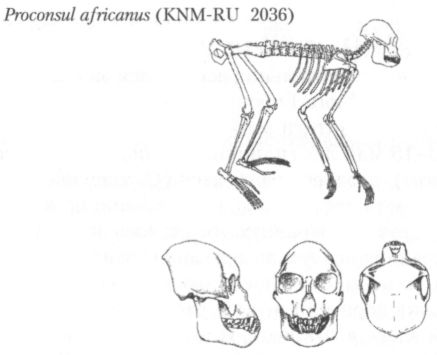
Рис. 7. Реконструкция скелета и черепа проконсула
В конце раннего миоцена в Африке жили уже представители нескольких родов гоминоидов: дендропитеки, микропитеки, афропитеки, турканопитеки и др., но филогенетическое значение этих форм неясно. Трудно сказать, имела ли какая-то из них непосредственное отношение к родословной современных горилл или шимпанзе. По размеру тела африканские раннемиоценовые гоминоиды варьировали от очень мелких, до 3 кг весом (Micropithecus clarki), до крупных (Proconsul major, Turkanapithecus heseloni), весящих около 100 кг, как самка современной гориллы, а их рацион включал главным образом фрукты и молодые листья. Все перечисленные формы вели преимущественно древесный образ жизни, а при передвижении по земле оставались четвероногими.[4] Единственным исключением из последнего правила был, возможно, ореопитек, или, точнее, вид Oreopithecus bamboli, но он обитал не в Африке, а в Европе, и не в начале, а в конце миоцена. Изучение костных останков ореопитека, найденных в Италии в отложениях возрастом 8–9 млн лет, привело ряд палеонтологов к предположению, что это существо, когда оно оказывалось на земле, предпочитало использовать для ходьбы не четыре, а две ноги.
В среднем миоцене, когда между Африкой и Евразией установилась сухопутная перемычка, (16–17 млн лет назад), ареал обитания гоминоидов значительно расширился за счёт включения в него территорий юга Европы и Азии. Наиболее древние ископаемые представители этой группы в Европе имеют возраст около 13–15 млн лет (плиопитек (Pliopithecus), дриопитек (Dryopithecus), позднее уранопитек (Ouranopithecus)), а в Азии порядка 12 млн лет. Однако, если в Азии, по крайней мере на её юго-восточных окраинах, им удалось основательно закрепиться, сохранившись там до сего дня (орангутанги, гиббоны, сиаманги), то в Европе условия оказались менее подходящими, и, пережив короткий период расцвета, к концу миоцена гоминоиды здесь вымирают. В отложениях возрастом моложе 7 млн лет их останков в Европе не найдено. В Африке в рассматриваемый период времени (от 15 до 5 млн лет назад) также отмечается значительное сокращение числа известных видов гоминоидов, но, несмотря на это, именно она всё же остаётся местом основных событий в их эволюции. О важнейших из этих событий, непосредственно связанных с происхождением человека, и пойдёт речь в следующих главах.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава II Эволюция или происхождение (родословная) живых существ
Глава II Эволюция или происхождение (родословная) живых существ Нам надлежит обсудить здесь этот вопрос, ибо в последнее время создалась невероятная путаница, благодаря смешению гипотез с фактами, между тем как мы желаем воздвигнуть наши предположения не на гипотезах, а
Глава 3. Происхождение жизни: химическая эволюция
Глава 3. Происхождение жизни: химическая эволюция Ничтожное ничто — начало всех начал. Теодор Рётке, "Вожделение" Теория химической эволюции — современная теория происхождения жизни — также опирается на идею самозарождения. Однако в основе ее лежит не внезапное (de novo)
Происхождение и эволюция австралопитеков
Происхождение и эволюция австралопитеков В настоящее время большинство антропологов считают, что род Homo берет свое начало от группы австралопитеков (хотя следует сказать, что некоторые ученые отрицают этот путь). Сами австралопитеки эволюционировали из дриопитековых
Часть I Происхождение и эволюция собак: комменсализм
Часть I Происхождение и эволюция собак: комменсализм Где я только ни бывал, повсюду мне встречались безнадзорные собаки, которые кормятся на улице, задворках, свалках. Они обычно небольшие, причем довольно схожи друг с другом по размерам и внешнему виду: весом редко больше
Участь человекообразных
Участь человекообразных Семейная форма брачных отношений — далеко не столбовой путь приматов, того отряда, к которому мы имеем честь принадлежать. У многих из них один самец спаривается с несколькими самками, причем зачастую «вся любовь» к самке после спаривания
Брачные отношения у человекообразных
Брачные отношения у человекообразных А как же наши ближайшие сородичи? В семейном отношении они мало похожи на человека. Орангутаны живут на деревьях, самцы не дерутся из-за самок и не заботятся ни о них, ни о детенышах, которые к четырем годам уходят жить в отдельные
ИНТЕЛЛЕКТ ЧЕЛОВЕКООБРАЗНЫХ ОБЕЗЬЯН [ 42 ]
ИНТЕЛЛЕКТ ЧЕЛОВЕКООБРАЗНЫХ ОБЕЗЬЯН[ 42 ] Что такое интеллект, разумность — стародавняя, тысячелетняя тема психологии, но до сих пор остающаяся полной для нее неопределенностью. Это я должен заключить по крайней мере из книги Келера об интеллекте человекообразных обезьян,
[СУЩНОСТЬ РАЗУМА У ЧЕЛОВЕКООБРАЗНЫХ ОБЕЗЬЯН И ОШИБОЧНОЕ ТОЛКОВАНИЕ КЕЛЕРА] [ 55 ]
[СУЩНОСТЬ РАЗУМА У ЧЕЛОВЕКООБРАЗНЫХ ОБЕЗЬЯН И ОШИБОЧНОЕ ТОЛКОВАНИЕ КЕЛЕРА][ 55 ] Акад. И. П. Павлов. — ...Теперь у меня есть две постоянные темы: с одной стороны, об обезьянах, с другой стороны, о господине Шеррингтоне. Обезьяны связаны с Келером. Может быть, лучше сказать, что, с
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а
9.5. Структура, происхождение и эволюция круговоротов и трофических цепей
9.5. Структура, происхождение и эволюция круговоротов и трофических цепей Жизнь со времени своего возникновения сформировалась как цепной процесс. Что касается трофических цепей, то, как мы упоминали ранее, они образовались «с конца», т. е. с редуцентов — организмов,
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а
9.5. Структура, происхождение и эволюция круговоротов и трофических цепей
9.5. Структура, происхождение и эволюция круговоротов и трофических цепей Жизнь со времени своего возникновения сформировалась как цепной процесс. Что касается трофических цепей, то, как мы упоминали ранее, они образовались «с конца», т. е. с редуцентов — организмов,
6. Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens
6. Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens Делались попытки оценить интеллект обезьян, человекообразных обезьян и гоминид до Homo sapiens, основываясь на теории Пиаже о развитии интеллекта у детей. Согласно теории Пиаже, дети проходят четыре стадии
Кооперация самцов и самок в сообществах у современных африканских человекообразных обезьян и у предков человека
Кооперация самцов и самок в сообществах у современных африканских человекообразных обезьян и у предков человека Разнообразие типов иерархических структур у современного человека можно рассматривать как результат эволюционного развития социальных структур у ранних