Австралопитеки
Австралопитеки
Несмотря на открытие в последние годы нескольких форм гоминид, более примитивных и древних, чем австралопитеки, последние всё же остаются главными «поставщиками» сведений о самых ранних стадиях эволюционной истории человечества. В отложениях возрастом от 4 до 2 млн лет кости их довольно многочисленны, и каждый год приносит новые находки. Судя по распределению возраста этих находок и их расположению (см. табл. 1 и рис. 10), австралопитеки появились и обитали первоначально в восточной части Африки, и лишь в самом конце плиоцена, около 3 млн лет назад, проникли на южную оконечность этого континента, а также в его центральные районы. За пределами Африки достоверных находок костей австралопитеков неизвестно, хотя время от времени появляются сообщения об обнаружении таковых в Восточной Азии. Сообщения эти, однако, воспринимаются большинством специалистов скептически, поскольку материалы, на которых они основываются, слишком фрагментарны и не поддаются однозначной идентификации. В ряде случаев было убедительно показано, что кости из Китая или Индонезии, первоначально приписывавшиеся австралопитекам, на самом деле принадлежали различным восточноазиатским обезьянам, довольно далёким от гоминид.
С Австралией австралопитеки, вопреки звучанию своего имени, также ничего общего не имеют. В переводе с латыни australis («аустралис») и греческого ??????? («питекос») слово «австралопитек» означает просто «южная обезьяна». Такое название дал новому роду в 1924 г. выделивший его антрополог Раймонд Дарт, изучавший коллекции ископаемых костей в Йоханнесбургском университете. Южная — потому что необычный череп, попавший в руки Дарта, был найден на юге Африки (в пещере Таунг), а обезьяна, потому что объём мозговой полости черепа был слишком мал, чтобы считать его человеческим. По этой же причине год спустя, когда Дарт публично выступил с идеей, что открытое им существо было одним из древнейших предков человека, английские коллеги южноафриканского учёного, задававшие тогда тон в палеоантропологии, приняли его гипотезу в штыки. В те годы считалось, что, коль скоро людей отличает от животных прежде всего разум, то, значит, первым шагом нашей эволюции должно было стать развитие мозга. В пользу этого мнения свидетельствовал как будто и знаменитый череп из Пильтдауна в Англии, где вполне человеческая мозговая полость сочеталась с обезьяньей челюстью. Однако, несмотря на враждебный поначалу приём, череп из Таунга выдержал проверку временем, и сейчас все признают его принадлежность гоминиду, а вот пильтдаунская находка оказалось просто фальшивкой, ловкой подделкой. Одним из немногих антропологов, кто сразу же без колебаний признал правоту Дарта, был его соотечественник, палеонтолог Роберт Брум, прославивший к тому времени своё имя открытием многих десятков новых родов и видов вымерших рептилий. Рассказывают, что, будучи по характеру человеком очень экспансивным, он, прочтя об открытии йоханнесбургского коллеги, сразу же отправился к нему с визитом, и, вбежав в лабораторию, пал на колени перед лежавшим на столе черепом австралопитека. Интересно, что именно Бруму суждено было через несколько лет (в 1936 г.) обнаружить второй похожий череп, а затем, в 1947 г., и третий (рис. 11), что немало способствовало окончательному утверждению рода австралопитека (Australopithecus) в качестве одного из наиболее достоверных и древних предков человека.
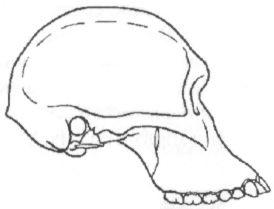
Рис. 11. Череп австралопитека (африканского) найденный Р. Брумом в пещере Штеркфонтейн в Южной Африке
Самой заметной и важной с точки зрения систематики особенностью ранних гоминид, сближающей их с людьми, является двуногость. Если для оррорина и сахельантропа наличие этого качества из-за скудости материалов всё ещё нельзя считать окончательно доказанным[14], то для австралопитеков, напротив, о нём можно говорить с полной уверенностью. Их анатомическая конструкция не оставляет сомнений в том, что они ходили не на четырёх, а на двух ногах. Прежде всего об этом свидетельствуют такие признаки, как короткий и широкий таз (рис. 12), сводчатая стопа, непротивопоставляемый большой палец ноги. О двуногости австралопитеков говорит и анализ их следов, сохранившихся в затвердевшем вулканическом пепле на местонахождении Лэтоли в Танзании (возраст следов от 3,2 до 3,6 млн лет). Ещё ряд признаков, прежде всего таких, как S-образный изгиб позвоночного столба и положение затылочного отверстия в центре (а не в задней части, как у обезьян) основания черепа, свидетельствуют о «стройности» австралопитека, выпрямленном положении его тела, что вполне соответствует выводу о прямохождении этого существа. В то же время, наряду с перечисленными особенностями, все виды рода австралопитека сохраняли в строении скелета и особенно конечностей некоторые черты, связанные ещё с древесным образом жизни, и, вполне вероятно, что немалую часть времени многие из них, действительно, проводили на деревьях.
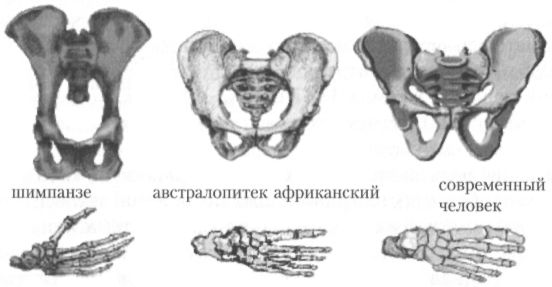
Рис. 12. Таз и стопа шимпанзе, австралопитека и современного человека. По строению этих частей скелета хорошо видно, насколько австралопитек ближе к людям, чем к человекообразным обезьянам
Помимо анатомических характеристик, являющихся результатом приспособления к передвижению на двух ногах, австралопитеков отличает от ископаемых и современных человекообразных обезьян ряд черт в строении зубной системы. Наряду с возросшей толщиной эмали, для них характерны относительно небольшие клыки и связанная с этим слабая выраженность или отсутствие диастемы, т. е. промежутка между клыками и предкоренными зубами в нижней челюсти, и между резцами и клыками в верхней, куда у обезьян входят клыки противоположной челюсти. Кроме того, очертания зубной дуги у австралопитеков, как правило, несколько более плавные, параболические, чем у обезьян, зубы которых образуют обычно два параллельных или почти параллельных ряда (рис. 13).
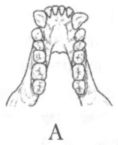
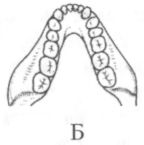
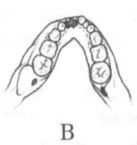
Рис. 13. Передняя часть нижней челюсти человекообразной обезьяны (А), австралопитека афарского (Б) и австралопитека африканского (В). Кроме различий в очертаниях зубной дуги, обращает на себя внимание также малый размер клыков на челюстях (Б) и (В) по сравнению с челюстью (А)
По абсолютному объёму мозговой полости (400–500 см3) австралопитеки в целом мало отличаются от шимпанзе, а гориллам даже несколько уступают. Это объясняется сравнительно небольшими размерами тела ранних гоминид, вес которых, согласно имеющимся реконструкциям, в большинстве случаев составлял от 30 до 50 кг (табл. 3). Что же касается относительной величины мозга, т. е. его веса или размера, взятых по отношению к весу или размерам тела, то по этому показателю австралопитеки, вероятно, хоть и не намного, но всё же превосходили всех человекообразных обезьян, как вымерших, так и современных.
Таблица 3. Австралопитецины: виды, время их существования, оценки массы и размеров тела. Для сравнения приведены также данные о массе тела шимпанзе (Pan troglodytes)
Вид Время существования (млн лет назад) Масса (кг) Рост (см) Мужчина Женщина Мужчина Женщина Pan troglodytes современность 49 41 — — Australopithecus anamensis 4,2–3,9 51 33 — — Australopithecus afarensis 3,9–3,0 45 29 151 105 Australopithecus africanus 3,0–2,4 41 30 138 115 Australopithecus garhi 2,5–? — — — — Paranthropus aethiopicus 2,7–2,2 — — — — Paranthropus boisei 2,3–1,2 49 34 137 124 Paranthropus robustus 1,9–1,4 40 32 132 110Самые древние останки австралопитеков происходят с местонахождений Канапои и Алиа Бэй в Кении. Здесь в 1994–97 гг. были найдены зубы и фрагменты челюстей, а также несколько костей скелета, не относящихся к черепу. Эти материалы, имеющие возраст от 4,2 до 3,9 млн лет, послужили для выделения вида австралопитек анаменсис (Australopithecus anamensis), представляющего собой наиболее раннюю и примитивную из всех известных сейчас форм рода австралопитека.
Видовое название этой формы происходит от слова «анам», означающего на языке местной народности «озеро». Это имя было выбрано потому, что Канапои, где была сделана самая первая находка, находится на берегу озера Туркана. Как и все представители рода австралопитеков, австралопитек анаменсис, несомненно, передвигался по земле без помощи верхних конечностей. Рацион его, в отличие от рациона человекообразных обезьян, включал в основном грубую, твёрдую растительную пищу, о чём свидетельствует возросшая — по сравнению с рамидусом и современными шимпанзе — толщина эмали на коренных зубах. Промежуточная хронологическая позиция анаменсиса и известные анатомические характеристики этого вида позволяют предположительно рассматривать его в качестве возможного связующего звена между ардипитеком, с одной стороны, и более поздними формами австралопитеков, с другой.
Систематика и филогенез австралопитеков давно находятся в числе наиболее активно обсуждаемых антропологами тем. В пределах этой группы выделяют сейчас до восьми видов: австралопитек анаменсис ( A. anamensis), австралопитек афарский (A. afarensis), австралопитек эфиопский (A. aethiopicus), австралопитек африканский (A. africanus), австралопитек бойсов (A. boisei), австралопитек робустус (A. robustus), австралопитек барелгазали (A. bahrelghazali), австралопитек гархи (A. garhi), а часто её делят ещё на два рода или подрода: собственно австралопитек (Australopithecus) и парантроп(Paranthropus). В один из этих родов (подродов) включают так называемые «грацильные» формы австралопитек афарский (A. afarensis), австралопитек африканский (A. africanus), австралопитек гархи (A. garhi), а в другой «массивные» парантроп бойсов (P. boisei), парантроп робустус (P. robustus), парантроп эфиопский (P. aethiopicus), причём разница между теми и другими заключается, главным образом, в величине челюстей и зубов. В качестве общего названия для австралопитеков и парантропов, как правило, используют термин «австралопитецины».
Недавние (1998–99) находки на местонахождении Ломекви в Кении (западное побережье оз. Туркана), включавшие довольно хорошо сохранившийся череп возрастом 3,5 млн лет, послужили основанием для выделения ещё одного рода гоминид, сосуществовавших с австралопитеками. Этот род, названный кениантроп (Kenyanthropus), представлен пока единственным видом Kenyanthropus platyops. От всех остальных известных сейчас своих современников кениантроп отличается прежде всего необычайно низкой степенью выступания нижней части лицевого скелета, сближаясь в этом отношении с более поздними гоминидами.[15] Эта особенность нового таксона отражена, кстати, в его видовом имени, которое образовано от греческих слов ?????? («платус») и ???? («опсис»), означающих, соответственно, «плоский» и «лицо».
Кстати, говоря о «видах» давно вымерших животных, следует помнить, что мы, собственно, не знаем, были ли они действительно видами в точном смысле этого слова. Дело в том, что главным критерием биологического вида, когда речь идёт об организмах с половым размножением, считается репродуктивная изоляция. Это значит, что в норме особи, принадлежащие к разным видам, либо не могут скрещиваться между собой, либо же не способны при скрещивании давать плодовитое (т. е. нестерильное) потомство. Понятно, что на ископаемых материалах выполнение последнего условия проверить невозможно, и потому приходится мириться с тем, что выделяемые по черепам, а то и по одним зубам палеонтологические виды могут не совпадать с видами биологическими. Строго говоря, нельзя исключить, что в каких-то случаях даже обладатели костей, относимых к разным родам (скажем, австралопитека и парантропа), при всех их внешних отличиях, на самом деле не были разделены барьером репродуктивной изоляции. И наоборот, этот барьер вполне мог существовать для некоторых особей, чьи останки принято включать в один вид.
Периоды существования большинства видов австралопитеков частично перекрываются. Единственным исключением в этом отношении является, возможно, вид австралопитек афарский (Australopithecus afarensis), описанный по находкам с танзанийского местонахождения Лэтоли, эфиопских местонахождений Феджедж, Билохдели, Мака и Хадар и кенийских местонахождений Кооби-Фора и Ломекви (рис. 14). Появившись примерно 4 млн лет назад, этот вид, насколько можно судить по имеющимся в настоящее время данным, оставался в течение следующего миллиона лет единственной формой австралопитеков, сосуществуя в течение какого-то времени лишь с кениантропом. Правда, в тот же хронологический интервал попадает ещё, возможно, вид австралопитек барелгазали (Australopithecus bahrelghazali), выделенный по челюсти, обнаруженной недавно в центре восточной Сахары (Чад) и датируемой временем от 3 до 3,5 млн лет назад, но при первом описании этой находки справедливо отмечалась её близость к австралопитеку афарскому (Australopithecus afarensis). Нельзя исключить, что в данном случае правильнее говорить не о новом виде, а лишь об особом подвиде австралопитека афарского: Австралопитек афарский барелгазали (Australopithecus afarensis bahrelghazali). Конечно, вполне возможно и даже вероятно, что в Африке в период от 3 до 4 млн лет назад жили и ещё какие-то виды австралопитеков, но их кости пока либо не найдены, либо не опознаны.
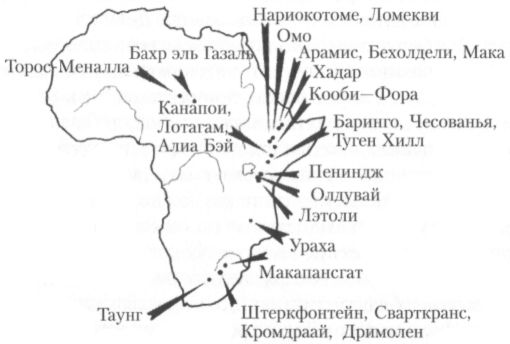
Рис. 14. Карта основных местонахождений, где были обнаружены костные останки австралопитеков и ещё более ранних гоминид
Австралопитека афарского рассматривают как правило в качестве предкового вида для всех остальных австралопитецин (кроме, разумеется, более древнего австралопитека анаменсис), или, по крайней мере, в качестве наиболее приемлемого аналога для воссоздания облика представителей такого вида. Конечно, и то и другое — лишь рабочие гипотезы, которым ещё предстоит выдержать (или не выдержать) проверку временем и новыми находками. Вообще допустимых вариантов реконструкции филогенеза ранних гоминид чрезвычайно много, и в ближайшем будущем их количество будет, скорее всего, увеличиваться, а не сокращаться. Сейчас всё чаще поднимается вопрос о том, в какой степени филогенетические схемы, построенные на анализе ископаемых костей, т. е. главным образом зубов, челюстей и иных фрагментов черепа, отражают действительные генеалогические взаимоотношения вымерших видов. Первые итоги обсуждения этой темы малоутешительны. В частности, было показано, что если бы те признаки и методы, которые служат в качестве базовых при построении родословного древа ранних гоминид, использовались для установления степени родства современных видов обезьян (как человекообразных, так и низших), то результаты оказались бы очень далёкими от реальности.[16] Антропологи, однако, вынуждены работать с тем материалом, который есть, возмещая его низкие разрешающие возможности совершенствованием методики анализа.
Особенно сложен и далёк от разрешения вопрос о том, какая из групп ранних гоминид послужила субстратом дальнейшей эволюции человека, т. е. дала начало роду Homo, и наличествуют ли вообще останки представителей такой группы в добытом к настоящему времени палеонтологическом материале. Ещё недавно наиболее перспективным, хотя и не безупречным, кандидатом на эту роль считался уже упоминавшийся выше австралопитек африканский (Australopithecus africanus), живший в период от 3 до 2,4 млн лет назад и известный главным образом по находкам в Южной Африке (местонахождения Таунг, Штеркфонтейн, Макапансгат), но в последнее время его позиции сильно пошатнулись. В результате детального сопоставления многочисленных костей верхних и нижних конечностей австралопитека африканского, открытых в ходе продолжающихся работ в Штеркфонтейне, выяснилось, что по соотношению их размеров этот вид находится намного ближе к человекообразным обезьянам, чем к гоминидам. Попросту говоря, руки у него были, скорее всего, значительно длиннее ног. Даже жившие на миллион лет раньше австралопитек анаменсис (Australopithecus anamensis) и австралопитек афарский (Australopithecus afarensis), по пропорциям конечностей выглядят более «продвинутыми». Это неожиданное открытие окончательно запутывает и без того запутанную ситуацию, поскольку по строению зубов и черепа австралопитек африканский, наоборот, намного ближе к Homo, чем австралопитек афарский, не говоря уже об австралопитеке анаменсис. Если предположить, что австралопитек африканский приобрёл это сходство с поздними гоминидами параллельно, т. е. не будучи звеном линии, ведущей к человеку, то тогда на первый план в качестве нашего возможного прямого предка выдвигается австралопитек афарский. В то же время нельзя исключить и противоположную возможность, а именно, что это как раз раннее приобретение конечностями австралопитека афарского человеческих пропорций совершилось параллельно и, следовательно, не является признаком, указывающим на близкую связь этой формы с Homo.
Недавно найденный в Штеркфонтейне череп австралопитека африканского, жившего примерно 2,6–2,8 млн. лет назад, имеет объём мозговой полости (антропологи называют её эндокраном) около 515 см3, что является рекордом для австралопитецин и сопоставимо с минимальными значениями, зафиксированными для представителей рода Homo. Правда, до недавнего времени считалось, что некоторые из поздних парантропов имели ещё более крупный мозг. Так, для двух черепов парантропа бойсов (Paranthropus boisei), датируемых началом плейстоцена (1,7–1,5 млн лет назад), ещё в 70-е годы был установлен размер мозговой полости порядка 525–545 см3. Однако новые реконструкции показали, что эти цифры сильно завышены, и что на самом деле по объёму эндокрана австралопитек африканский, как минимум, не уступает парантропам, а по структурным особенностям мозга стоит к Homo ближе, чем другие, известные сейчас, виды австралопитецин.
В ноябре 1997 г. на одном из палеонтологических местонахождений в районе Среднего Аваша (Эфиопия) был найден череп гоминида возрастом около 2,5 млн лет, послуживший основанием для выделения вида, названного австралопитек гархи (Australopithecus garhi) (слово garhi на афарском языке означает «сюрприз»). Сохранились лобная и теменные кости, а также верхняя челюсть с зубами. Несмотря на относительно большие размеры последних, австралопитек гархи лишён специфических черт массивных австралопитеков. Не похож он и на австралопитека африканского, от которого, как и от ранних представителей рода Homo, его отличает более примитивное строение лобной кости и лицевого отдела черепа, а также малый объём мозга (всего около 450 см3). Несмотря на последнее обстоятельство, по мнению исследователей, выделивших новый вид, анатомия австралопитека гархи достаточно нейтральна для того, чтобы рассматривать его в качестве одного из вероятных предков ранних Homo. Впрочем, следует ожидать, что уже в самом близком будущем число видов и даже, возможно, родов гоминид, претендующих на эту роль, возрастёт ещё более, если полевые исследования, проводимые палеоантропологами в Африке, сохранят свой нынешний размах.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Австралопитеки
Австралопитеки Вернемся к истории. Если длинная серия лирических отступлений не сбила читателя с толку, то он еще помнит, что мы остановились на ардипитеках, живших в Восточной Африке 4,4 млн лет назад. Вскоре после этого, примерно 4,2 млн лет назад, на африканскую сцену
Австралопитеки пошли своим путем
Австралопитеки пошли своим путем Приспосабливаясь к питанию грубой растительной пищей, часть потомков Люси 2,8 млн лет назад дала начало новому виду, более мощному, с укороченными клыками, — африканскому австралопитеку. От него 2,3 млн лет назад — еще один вид. Все эти