3.3. Структура хромосом
3.3. Структура хромосом
Каждая хроматида содержит одну молекулу ДНК, связанную с белками-гистонами и негистоновыми белками. В настоящее время принята нуклеосомная модель организации хроматина эукариот (Kornberg R., 1974; Olins А., Olins D., 1974).
Согласно этой модели, белки-гистоны (они практически одинаковы у всех эукариот) формируют особые глобулы из 8 молекул в каждой глобуле (по две молекулы гистонов Н2а, Н2б, Н3, Н4). Нить ДНК делает по два витка вокруг каждой глобулы. Структура, состоящая из гистонового октамера, обвитого участком ДНК (размером 140–160 п. н.), называется нуклеосомой. Такая укладка ДНК сокращает ее длину в 7 раз. Нуклеосомная модель получила название «бусинки на нитке». Положительно заряженные гистоны и отрицательно заряженная ДНК образуют относительно прочный ДНК-гистоновый комплекс.
Участок ДНК между нуклеосомами содержит гистон Н1. Он играет важную роль в спирализации нуклеосомной нити и образовании второго уровня организации хромосом – винтообразной структуры соленоида. Последующая многоступенчатая укладка ДНК-гистоновой нити во многом остается областью, благодатной для различных гипотез. Один из вариантов изображен на рис. 3.2. Компактная упаковка генетического материала в хромосоме получила название процесса компактизации хроматина. Всего выделяют 4–5 уровней упаковки, начиная с нуклеосомного.
Степень компактизации хроматина различается в разных участках хромосом и зависит от периода клеточного цикла. Важную роль в этом процессе играют разнообразные негистоновые белки. Благодаря процессу компактизации, гигантские молекулы ДНК упакованы в клетке в небольшом объеме. Например, ДНК хромосом человека общей длиной около 1,8 м упакована в ядре диаметром менее 1 микрометра.
Необходимо отметить, что хроматин (вещество хромосом) у эукариот упакован неодинаково. Различают два типа хроматина: эухроматин (упакован менее плотно) и гетерохроматин (упакован более плотно). В свою очередь, гетерохроматин разделяют на два класса: структурный (или конститутивный) гетерохроматин (постоянно выявляемые участки) и факультативный гетерохроматин (участки обратимой компактизации эухроматиновых районов). Структурный гетерохроматин локализован в прицентромерных областях и некоторых других районах хромосом, он хорошо выявляется С-окраской. В интерфазе участки структурного гетерохроматина часто агрегируют друг с другом и образуют хромоцентры.
Считается, что гетерохроматин генетически неактивен в связи с высокой степенью конденсации, а эухроматин – активен. Но, с другой стороны, нахождение в эухроматине является недостаточным условием для экспрессии генов. Еще больше вопросов возникает при изучении функционирования гетерохроматина. Несмотря на многолетнюю историю интенсивного изучения структурно-функциональных особенностей разных видов хроматина, в этой проблеме остается много неясного.
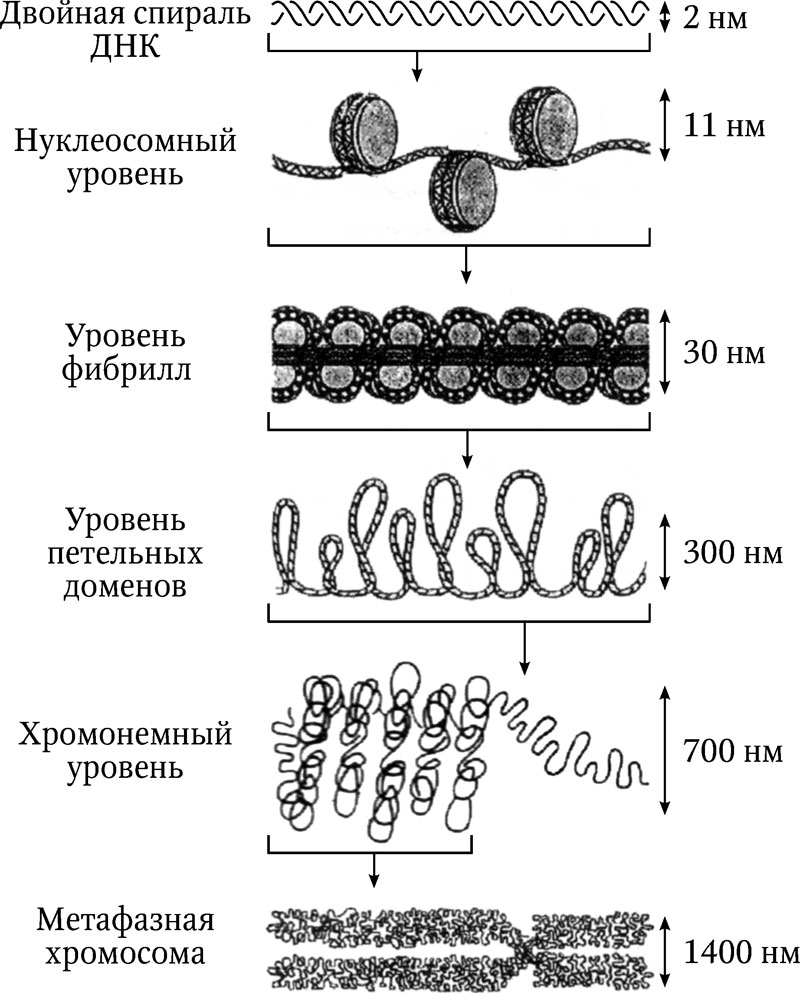
Рис. 3.2. Уровни организации хроматина эукариот
У некоторых организмов, наряду с постоянными хромосомами, в ядрах обнаружены дополнительные хромосомы – так называемые В-хромосомы. Часто они целиком состоят из гетерохроматина. Функции их до конца не понятны.
В природе наблюдаются случаи нетипичной структуры хромосом. Поскольку такие нетипичные хромосомы имеют крупные размеры, они служат удобной моделью для изучения генома.
Хромосомы типа «ламповых щеток» представляют собой растянутый и раскрученный вариант обычных хромосом ооцитов во время длительного мейоза. Лучше всего они изучены у амфибий, в связи с их особо крупными размерами. Длина таких хромосом в 30 раз превышает их длину в обычном состоянии. Хромосомы типа «ламповых щеток» получили свое название из-за наличия петель. Петли – это участки хромосомной нити, выступающие из более компактного материала и являющиеся местом активной транскрипции. В конце мейоза хромосомы типа «ламповых щеток» возвращаются к обычному состоянию.
Политенные хромосомы образуются в некоторых клетках в результате максимальной деспирализации и многократной репликации без последующего расхождения хромосом. Это явление называется эндомитозом. Перед эндомитозом гомологичные хромосомы соединяются попарно – конъюгируют. Такая конъюгация не характерна для других соматических клеток. Все политенные хромосомы кариотипа объединяются центромерами в общий хромоцентр. Лучше всего политенные хромосомы изучены у двукрылых насекомых (в том числе у классического объекта – дрозофилы), хотя встречаются и у некоторых других организмов.
Поскольку политенные хромосомы содержат более 1000 нитей, они в 1000 раз толще обычных хромосом и у них хорошо видны участки более плотной спирализации – диски. В геноме дрозофилы выявлено около 5000 дисков – все они пронумерованы и формируют цитологические картыхромосом. Каждый диск представляет собой самостоятельную функциональную единицу, содержащую от одного до нескольких генов. Во время экспрессии активные диски «вздуваются» и образуют пуфы, которые появляются и исчезают в определенной последовательности, в зависимости от активности генов на разной стадии онтогенеза.
Цитологический анализ хромосом этих двух типов заложил основы представлений о хромомерном принципе организации хромосом. Хромомеры – это участки временно конденсированной неактивной ДНК. Расположение хромомеров для каждой хромосомы относительно постоянно. Хромомеры могут деконденсироваться и переходить в активное состояние, формируя петли, на которых происходит синтез РНК.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Структура антител
Структура антител Теперь кратко рассмотрим структуру молекулы антитела. Все антитела имеют общий план строения — это белки, состоящие из нескольких субъединиц. На рис. 3.2 показано строение наиболее обычного антитела, называемого IgG. Эта структура была впервые описана в
2. СТРУКТУРА МУРАВЕЙНИКА
2. СТРУКТУРА МУРАВЕЙНИКА Облик муравьев различен. Среди нескольких тысяч видов муравьев есть и карлики размером до 2 мм, и великаны, у которых рабочие достигают 2 см. Белесые, почти прозрачные и глянцево-черные, одноцветные, двухцветные и т. д. Разнообразны форма головы и
Повреждения хромосом
Повреждения хромосом Как вы могли заметить из содержания предыдущих глав, различные нарушения пола у человека чаще всего вызываются добавочными половыми хромосомами. Для специалиста посчитать число хромосом в клетках любого многоклеточного организма относительно
1. Структура
1. Структура ВИЧ относится к семейству ретровирусов.Вирион имеет сферическую форму, диаметром 100–150 нм. Кубический тип симметрии. Наружная (суперкапсидная) оболочка вируса состоит из бимолекулярного слоя липидов, который имеет происхождение из клеточной мембраны клетки
Сексуальные игры хромосом
Сексуальные игры хромосом Иногда природа шалит. Она может наградить человека лишней хромосомой или, напротив, недодать одну. Порой она перемешивает хромосомные наборы матери и плода, или двух близнецов. Подобные эксперименты природы приводят к появлению на свет людей с
В чьей клетке больше хромосом – человека или утки?
В чьей клетке больше хромосом – человека или утки? Для каждого организма характерно строго определенное число хромосом, содержащихся в каждой из составляющих его клеток. У плодовой мушки (дрозофилы) 8 хромосом, у сорго – 10, у садового гороха – 14, у кукурузы – 20, у жабы – 22,
Может ли набор хромосом преступника служить оправданием совершенного им преступления?
Может ли набор хромосом преступника служить оправданием совершенного им преступления? Одним из нарушений со стороны половых хромосом является лишняя Y-хромосома в кариотипе (совокупности признаков хромосом, характерной для клеток тела организма того или иного вида)
Структурно-функциональная организация хромосом
Структурно-функциональная организация хромосом Морфология хромосом эукариот характеризуется единством структурной организации. Каждая хромосома кариотипа представляет собой сложную структуру, в которой нить ДНК взаимодействует с различными белками. Благодаря
7.2. Структура биоценоза
7.2. Структура биоценоза Структура любой системы – это закономерности в соотношении и связях ее частей. Структура биоценоза многопланова, и при изучении ее выделяют различные
Структура сна
Структура сна Ночной сон состоит из нескольких циклов, каждый из которых длится примерно полтора часа и включает в себя две качественно различные стадии, которые называются «медленный сон» и «быстрый сон». Стадии быстрого и медленного сна есть не только у человека, но и
Соотношение половых хромосом и аутосом как фактор определения пола
Соотношение половых хромосом и аутосом как фактор определения пола Ученые уже в первой половине XX века активно обсуждали вопрос о том, что же лежит в основе определения пола. Широкую известность получила балансовая теория К. Бриджеса. По мнению американского генетика, не
Эволюция половых хромосом
Эволюция половых хромосом Исследователи давно интересуются эволюцией половых хромосом. В 1991г. Б. Чарльсворс опубликовал на эту тему специальную статью в Nature. М. Маджерус в недавно вышедшей книге (2003) доказывает, что половые хромосомы эволюционировали многократно и