Глава 5. Неоламаркизм
Это течение эволюционной мысли и по времени своего возникновения, и по той роли, которую оно сыграло в истории борьбы эволюционных идей, должно быть поставлено на первое место. Само название «неоламаркизм» ввел в научный обиход американский палеонтолог А. Пакард (1888). Им охватывается совокупность весьма разнородных эволюционных концепций, но имеющих общий источник — эволюционное учение Ламарка.
Наивно было бы ожидать, чтобы по прошествии более полувека учение Ламарка могло возродиться в исконном виде. Прогресс биологии заставил навсегда отказаться от веры в самопроизвольное зарождение, от флюидов, от отрицания фактов вымирания видов, от формообразующего действия волевых усилий и многих других наивных суждений Ламарка. Все эволюционисты, кого стали называть неоламаркистами, в той или иной мере отклонились и от основных его постулатов, и многие — очень значительно. В ряде теорий были развиты или. модернизированы отдельные стороны доктрины Ламарка (или несколько его положений в том или ином сочетании с новыми идеями), и они дали начало автономным направлениям проламарковской ориентации. Однако в большинстве новых концепций можно обнаружить по крайней мере два постулата, соответствующих законам Ламарка: признание адекватной изменчивости, возникающей под непосредственным или косвенным влиянием среды и создающей приспособление, и унаследование приобретенных в индивидуальной жизни признаков. Общей отличительной чертой неоламаркизма является либо полное отрицание какой-либо эволюционной роли естественного отбора, либо рассмотрение его как вспомогательного инструмента. Неоламаркизм занял принципиально отличные от дарвинизма позиции по таким кардинальным вопросам, как объект, материал, движущие силы и направленность эволюции.
Потомство сполна «отомстило» современникам Ламарка за его непризнание. С того момента как Дарвин вызволил имя ученого из тени забвения, благодарные потомки вдруг оценили идеи Ламарка и вознесли их на вершину научного Олимпа. История повелела, чтобы имена этих двух гениев, по идее антиподов и соперников, отныне всегда были рядом.
Посмертная известность и слава Ламарка восторжествовали во всем мире. Во Франции, дольше всего сопротивлявшейся идее эволюции, ее победа в конце 70-х годов XIX в. закономерно была воспринята как торжество ламаркизма. Этому способствовали обостренное чувство национального самосознания, приоритет в создании первого эволюционного учения, удаленность большинства французских биологов от запросов сельскохозяйственного производства, большая терпимость к ламаркизму, нежели дарвинизму, позитивная методология Конта (ее приверженность к наблюдениям и умозаключениям, отвечающим требованиям «здравого смысла»).
Первыми, кто вспомнил Ламарка и его учение, были Анри Лaказ-Дютье (1866) и Эрнст Геккель (1863). В 1865 г. Лаказ-Дютье был избран профессором той же кафедры в Национальном музее естественной истории, которую в свое время возглавлял Ламарк. Занимаясь подготовкой публикации по истории этой кафедры, Лаказ-Дютье был вынужден ознакомиться с жизнью и трудами Ламарка, Следует отдать должное этому крупному ученому-зоологу. Будучи далеким от эволюционных представлений, он счел необходимым совершенно объективно изложить эволюционные взгляды Ламарка во вступительной лекции того же года, а в следующем году — опубликовать о Ламарке отдельную книгу (Lacaze — Duthiers, 1866). С его легкой руки о Ламарке скоро стали писать многие другие авторы — появились книги уже знакомого нам Катрфажа (1870), Мартэна (1873), Кадоде Кервиля (1883), Ланессана (1883), Дюваля (1889) и др. В короткое время Ламарк был поднят на пьедестал, а его учение стало предметом национальной гордости и восхищения.
Однако главными зачинателями неоламаркизма чаще всего считают английского философа и социолога Герберта Спенсера и американского палеонтолога Эдуарда Копа. Их оригинальные труды (Спенсер, 1870, 1894; Spencer, 1896; Соре, 1868, 1896) и пропаганда идей неоламаркистского содержания более всего способствовали упрочению нового течения в мировом масштабе.
В неоламаркизме традиционно выделяют три главных направления — механо-, орто- и психоламаркизм. Мы рассмотрим каждое из них отдельно.
Основателем механоламаркизма, самого представительного и широко распространенного направления, является опять-таки Спенсер. В Германии из числа наиболее известных эволюционистов его представлял Теодор Эймер, а во Франции к нему принадлежали почти все неоламаркисты во главе с Альфредом Жиаром.
Механоламаркисты игнорировали ведущую идею Ламарка о главенстве в эволюции внутреннего стремления организмов к совершенствованию, об их поведенческой активности во взаимоотношениях с окружающей средой, В отличие от Ламарка они всецело подчинили организм внешним условиям и сделали последние главными факторами эволюционного развития (эктогенез). Взаимоотношения с внешними условиями трактовались по Ламарку: соответствие этим условиям может достигаться либо путем прямого приспособления, либо в результате функциональной деятельности организма (т. е. через «упражнение и неупражнение органов») и унаследования приобретенных признаков.
Подтверждение такому механизму эволюции видели в многочисленных наблюдениях и экспериментах, в которых исследователи имели дело преимущественно с адаптивными модификациями. Последние воспринимались как первичное эволюционное явление. К такого рода экспериментам относились прежде всего работы морфологов растений, проведенные под руководством знаменитого французского ботаника Гастона Боннье. Выращивая растения в специально выбранных естественных условиях, Боннье и его сотрудники отмечали высокую степень изменчивости морфофизиологической организации растений, причем чаще всего она носила приспособительный характер.
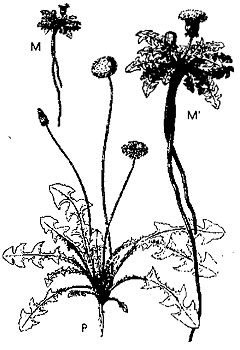
Рис. 5. Общий вид одуванчика Taraxacum Dens-Leonis: Р — выращен на равнине; М — выращен в горах; М’ — тот же одуванчик в натуральную величину (из: Bonnier, 1894].
Конкретно Боннье исследовал влияние на растения высокогорного климата. Эксперименты проводились на 203 видах растений, начиная с обычного одуванчика и кончая плодовыми культурами.
Опытные участки были заложены в Парижском ботаническом саду и в Альпах и Пиренеях на высоте 2000 м над уровнем моря. 123 вида, не выдержав суровых высокогорных условий, погибли, а у 80 выживших общий габитус и целый ряд морфологических признаков изменились до неузнаваемости. Под прямым воздействием необычных условий у растений сильно сократились или вовсе исчезли междоузлия, они стали бесстебельными, карликовыми, напоминающими подорожник розеткой прижатых к земле листьев. У них развился мощный, тоже сильно укороченный корень, сократилось число цветков, но они стали крупнее и ярче окрашенными. Различия между индивидами одного и того же вида, выросшими в Париже и в горах, были настолько резкими, что, не зная условий опыта, их трудно было признать за растения одного вида. Во многие учебники вошел рисунок Боннье (рис. 5), демонстрирующий различия между долинной и горной формами обыкновенного одуванчика. Этот пример аналогичен тому, что наблюдал Ламарк у водного лютика (Bonnier, 1894; см. также: Костантэн, 1908).
К числу наиболее показательных свидетельств справедливости механоламаркизма принадлежали также, с точки зрения его лидеров, опыты и наблюдения Г. Кутаня (1894–1895) на моллюсках, В. И. Шманкевича (1875) — на рачках родов Artemia и Brachipus, М. Штандфуса и Э. Фишера (1901–1911) — на бабочках родов Araschnia (Vanessa) и Arctia, ставших излюбленными объектами неоламаркистов, и многих других.
После исторического выступления А. Вейсмана (1883) против возможности наследования приобретенных признаков и афористического возражения ему Г. Спенсера — «или существует наследственность приобретенных признаков, или не существует эволюции» — многие механоламаркисты принялись усиленно экспериментировать. В опытах того же Кутаня (Coutagne, 1903), но теперь уже на шелковичном черве, Ф. Уссе (1907) — на курах и особенно Ш. Броун-Секара (1869–1882) — на морских свинках, выполненных еще до выступления Вейсмана, на этот «вечный» вопрос были получены положительные ответы.
Борьба вокруг проблемы наследования приобретенных признаков, с тех пор как она приняла первоначальную остроту почти 120 лет назад и превратилась в одну из центральных в эволюционной теории, вкупе с неотделимым от нее тезисом об адекватной изменчивости, стала причиной множества трагических событий, тяжелых баталий и перипетий, а в бывшем Советском Союзе стоила жизни сотням биологов. Проблема такого наследования — это птица феникс, но не единожды сожженная и воскресшая. Новейшая генетика дает ей теперь законные права на положительное решение, и такое решение скорее всего уже необратимо. Специально речь об этом пойдет в гл. 14 и 16.
Вернемся, однако, в XIX в. и рассмотрим вкратце эволюционные представления типичных представителей механоламаркизма.
Альфред Жиар — основатель и патриарх французской школы неоламаркизма, первый глава созданной в Сорбонне в 1888 г. кафедры эволюции организованных существ. Именно с этим событием можно связать победу эволюционизма над креационизмом во Франции (см.: Назаров, 1974).
Причину эволюции и ее важнейшую движущую силу Жиар (Giard, 1889) видит во внешней среде, с которой организм находится в самом тесном «динамическом равновесии». К числу ее главных факторов, именуемых первичными, он относит космические (климат, состав почвы и воды, механическое состояние среды, ветер, движение вод и т. д.) и биотические (пиша, наличие паразитов, симбиоз и пр.) условия, а также этологические реакции на эти условия, выражающиеся в приспособлении, явлениях конвергенции, защитного сходства, мимикрии и др.

Альфред Жиар (1846–1908).
Названным факторам, изучение которых составляет предмет ламаркизма, Жиар придает первостепенное значение в эволюции не только как единственным причинам изменчивости, но и как силам, достаточным для того, чтобы обеспечить прогрессивное формообразование. Он пишет, что «постоянство или периодичность действия первичного фактора, например изменения среды, есть необходимые и достаточные условия для возникновения разновидности или вида без участия какого бы то ни было вторичного фактора» (Giard, 1904. Р. 121), При этом Жиар, отмечая, что Дарвин не отрицал эволюционного значения данных факторов, упрекает его в недостаточном к ним внимании, в недооценке природы условий в возникновении изменчивости и, наоборот, в переоценке природы организма.
Признавая адекватный характер изменчивости, вытекающий из предполагаемой «точной причинной связи» между первичными факторами и вызываемыми ими изменениями, Жиар считает, однако, что одна изменчивость не в состоянии создать новую форму без аккумулирующего действия наследственности. Последняя, хотя формально и включена автором в число вторичных факторов, занимает самостоятельное место. Она трактуется чисто механистически, в соответствии с эктогенетическим пониманием эволюционного процесса. «Что же такое наследственность, — пишет Жиар, — если не повторение в определенный момент физико-химических и механических условий, идентичных с теми, которые определили у родителей морфологическое и физиологическое состояние, подобное тому, которое проявляется в данный момент у потомства?» (op. cit., р. 147). И далее отмечает: «Наследственность, собственно, не есть… ни первичный, ни вторичный фактор. Это интеграл, сумма бесконечно малых изменений, произведенных первичными факторами на каждое предшествующее поколение» (op. cit., р. 153). Получается, что природа живого организма — это простое отражение совокупности условий внешней среды.
Принцип наследования приобретенных признаков принимается Жиаром во всей полноте и представляется ему необходимым условием эволюции. Подтверждение ему Жиар находит в результатах уже упомянутых опытов Броун-Секара, экспериментах А. Лундстрема по так называемому наследственному галлообразован ию (этот исследователь якобы доказал, что деформация листьев липы клещами воспроизводится и при изоляции деревьев от этих членистоногих), по вегетативной «гибридизации» растений, а также в явлении телегонии. Более поздними исследованиями других авторов было установлено, что все перечисленные факты прямого отношения к наследственности не имеют.
Наряду с первичными факторами Жиар признает эволюционное значение и вторичных факторов, к которым относит жизненную конкуренцию, естественный и половой отбор, различные формы изоляции, гибридизацию и т. п. Из них наиболее важным Жиар считает естественный отбор (рис. 6).
Роль естественного отбора, по Жиару, очень эффективна в ускорении эволюционного процесса. В этом случае он действует путем усиления первоначального импульса внешней среды и устранения неудачных индивидов. В создании новых форм и их тонком приспособлении к условиям местообитания отбор, однако, непосредственного участия не принимает. «Отбор ничего не создает: выбирать — не значит творить, но благодаря уничтожению менее приспособленных отбор оказывается замечательным механизмом, фиксирующим то, что полезно. Он сохраняет завоевания предков» (Giard, 1907. Р. 453).

Рис. 6. Факторы эволюции (Жиар. 1904).
Как все неоламаркисты, Жиар отказывается видеть в отборе нечто большее, чем простой сортирующий механизм. Вместе с остальными вторичными факторами он сравнивает его с призмой, разделяющей световой поток на лучи разного цвета, и с гранью кристалла, пропускающей только поляризованные лучи. Жиар неоднократно протестовал против преувеличения роли того или иного вторичного фактора, к чему были склонны неодарвинисты Вейсман, Уоллес, Ромене, М. Вагнер, и считал неоправданным слишком большое внимание к их изучению. Вместо этого он призывал к углубленному исследованию влияния первичных факторов, мечтая о создании специального института по проблемам эволюции и постановке длительных (более продолжительности человеческой жизни) и дорогостоящих опытов по изучению последствий усиленного и ослабленного упражнения органов.
Впрочем, намерение исключить вторичные факторы из числа творческих причин эволюции у Жиара в ряде конкретных случаев не выполняется, и это отличает его от других механоламаркистов и собственных последователей. Иногда он допускает участие отбора в определении направленности отдельных филогенетических линий и в выработке самого приспособления. Он готов даже признать значение естественного отбора в материалистическом объяснении органической целесообразности и «изгнании дуалистической мысли о конечных целях в отношениях живых организмов между собой и к окружающей среде». В другом месте Жиар говорит: «Это будет означать и хвалу Дарвину, показавшему, как… первичное действие среды усиливается естественным отбором, смысл которого, в сущности, неотделим от понятия адаптации, где действие отбора определяется ее пределами» (Giard, 1904. Р. 159). Отмечая трудности в изучении причин эволюции, Жиар пишет, что далеко не всегда можно установить, что в образовании нового вида или разновидности связано с действием первичных и что — с действием вторичных факторов. Он указывает, в частности, на бабочку Vanessa (Araschnia) levanaprorsa, имеющую две сезонные генерации, которые отличаются морфологически. Причина ее диморфизма — исключительно температурный фактор, что доказывается опытным превращением одной формы в другую. Если бы формы levana и prorsa в природе были пространственно разграничены, их возникновение можно было бы приписать вмешательству вторичного фактора. Жиар приводит также примеры совместного действия первичных и вторичных факторов на некоторые виды креветок и мух, различающиеся только по яйцам или личинкам и обладающие совершенно сходными морфологическими признаками во взрослом состоянии. Все это идет вразрез с основной идеей ученого о приспособительном характере непосредственных реакций организмов на внешнюю среду. Таким образом, эволюционная концепция Жиара, будучи по своему существу механоламаркистской, включала в себя и известную долю дарвинизма.
Герберт Спенсер пришел в биологию от философии. Его взгляды на биологическую эволюцию, сложившиеся в середине 60-х годов XIX в., вытекали из общего представления о развитии как движении к состоянию равновесия, которое, однако, никогда не может быть достигнуто.
Согласно взглядам Спенсера, организм представляет собой «агрегат частей, называемых органами», которые находятся в состоянии подвижного равновесия между собой и в отношениях с внешней средой. Сама жизнь — это «беспрерывное приспособление внутренних отношений к отношениям внешним» (Спенсер, 1870. С. 257–258). Функции органов устроены так, что уравновешивают действие на организм внешних сил. Но последние все время изменяются и если не разрушают равновесие, достигнутое между организмом и средой, обрекая организм на смерть, то порождают в нем «неуравновешенную силу», которая способна изменить функции и строение его органов и восстановить утраченное равновесие. Отсюда можно заключить, что Спенсер видит сущность эволюции в наследственных изменениях организмов, вызванных условиями среды и направленных на уравновешивание с новыми условиями. Равновесие всегда относительно, ибо изменения не поспевают за переменами среды.

Герберт Спенсер (1820-1903).
Уравновешивание носит адаптивный характер и осуществляется двумя способами — непосредственно и косвенно. При непосредственном уравновешивании организм изменяется под прямым воздействием факторов среды (космических, геологических, климатических условий, взаимодействия с другими организмами), и эти изменения сразу наследуются и передаются следующим поколениям. Наследование приобретенных свойств Спенсер вообще рассматривает в качестве непреложного закона эволюционного развития. У животных изменения происходят через преобразование функций органов.
Мы видим, таким образом, что в этой части своей концепции Спенсер целиком стоял на позициях механоламаркизма. Но он допускал и косвенное уравновешивание, под которым понимал приспособление посредством естественного отбора. Объектами приложения его сил он считал различные пассивные образования животных, выполняющие защитные и опорные функции или служащие средством нападения (панцири, рога, копыта, когти). С помощью естественного отбора Спенсер объяснял также эволюционные изменения плодовитости, численности видов, становление коадаптаций, связанных с размножением. Таким образом, в эволюционных представлениях Спенсера, как и в концепции Жиара, исходный ламаркизм сочетался с частью дарвинизма.
Однако Спенсер сознательно стремился всячески ограничить эволюционное значение отбора. Уже в «Основаниях биологии» (Spencer, 1864; рус. пер. — J 870) он заявил о себе как о стороннике постепенного изменения самих факторов эволюции в ходе исторического развития жизни. К. М. Завадский и Э. И. Колчинский (1977), подробно осветившие этот вопрос, установили, что Спенсер связывал преобразование этих факторов с изменениями абиотических условий, повышением уровня организации живого и его активности, возрастанием видового разнообразия, усложнением биотических отношений и усилением их влияния на эволюционный процесс. Сообразно этим изменениям менялась и значимость естественного отбора. На заре развития органического мира он служил основной движущей силой эволюции. В дальнейшем, однако, по мере снижения плодовитости и численности видов, увеличения продолжительности жизни особи, повышения ее активности, развития заботы о потомстве и т. п. эволюционная роль отбора неуклонно снижалась, и в результате он сохранил известное значение только в эволюции низших организмов. И как считает Спенсер, даже по отношению к ним он оказался крайне неэффективным, о чем свидетельствуют низкие темпы их эволюции.
Напротив, по мере прогрессивной эволюции творческая роль все более переходила к фактору прямого приспособления и наследованию приобретенных признаков, как мы наблюдаем это на примере высших животных.
В 1893 г., полемизируя с Вейсманом, Спенсер выступил со статьей «Недостаточность естественного отбора» (Спенсер, 1894), где с особой силой подчеркнул первостепенную эволюционную роль ламарковских факторов. Одновременно он отметил, что в ходе эволюции естественный отбор превратился в элиминирующий механизм, поддерживающий лишь общую жизнеспособность населения вида и устраняющий нежизнеспособных особей.
Спенсеру принадлежит заслуга в изучении механизмов поддержания равновесия на уровне биотических сообществ.
Перейдем теперь к следующему направлению — ортоламаркизму (от греч. orthos — прямой). Термин ортогенез был введен немецким биологом В. Гааке (Нааке, 1893). Основателем направления многие считают немецкого зоолога Т. Эймера.
Главная отличительная черта ортоламаркизма[13] — постулирование прямолинейного характера эволюции, связанное по преимуществу с внутренними (конституционными) причинами, заложенными в живых организмах (автогенез). Этот постулат непосредственно продолжает идею Ламарка о градации, выражающей неуклонное стремление к повышению уровня организации, к усовершенствованию и прогрессу, не зависящее от влияния окружающей среды. Сторонниками автогенети чес кого понимания направленности эволюции были в основном палеонтологи. Однако к ортоламаркизму следует отнести также немногих эволюционистов, которые более склонны видеть в эволюционных преобразованиях организмов результат непосредственного воздействия внешней Среды (эктогенез), канализуемого внутриорганизменными ограничениями. При этом как автогенетики, так и эктогенетики придерживались убеждения, что направленность эволюции проистекает из направленности сдмой изменчивости, последовательно усиливающейся благодаря механизму наследования новоприобретений без всякого участия естественного отбора. Самыми известными приверженцами ортоламаркизма были, кроме Эймера, К. Негели, Э. Коп, А. Гайетт и Г. Штейнман.
Остановимся сначала на взглядах немецкого зоолога Теодора Эймера, стоящего преимущественно на позициях эктогенеза. Этот ученый признает ведущим фактором эволюции воздействие на организм внешних раздражителей — температуры, влажности, света, пищи и т. п., вызывающих соответствующие целесообразные физические и химические изменения в протоплазме, которые передаются по наследству из поколения в поколение (Eimer, 1888, 1897), Формообразующее действие внешних раздражителей осуществляется через процесс «органического роста», в основе которого лежат те же физико-химические процессы, что и в явлении роста кристаллов.
К своей концепции ортогенеза Эймер пришел индуктивным путем. Он собрал большой материал по изменчивости рисунка крыльев бабочек-листовидок рода Kallima — тех Самых, которые служат классическим примером мимикрии, — и пытался построить по этому признаку эволюционный ряд. В итоге он возвел свою концепцию ортогенеза как «определенно направленного развития» в ранг всеобщего закона эволюции, которому подчиняется филогенез как животных, так и растений (Eimer, 1897. S. 14).
Надо заметить, что даже последовательный эктогенез ввиду допущения им принципа изначальной целесообразности живого не может обойтись без элементов автогенеза в качестве его необходимого дополнения. Поэтому уместен вопрос: достаточно ли для объяснения направленного развития, или, по терминологии Эймера, филогенетического роста, одних внешних факторов? Однозначного ответа на этот вопрос в трудах Эймера нет. Ссылки на аналогию с направленным ростом кристаллов малоубедительны, и, сознавая это, Эймер допускает влияние каких-то внутренних причин, но при этом оговаривает, что совершенно не согласен с той интерпретацией, которую дает этим причинам К. Негели. Эймер отмечает также, что филогенетический рост — это проявление того же единого органического роста, от которого зависит как историческое, так и индивидуальное развитие (Eimer, 1888. S. 24).
В поисках более убедительных причин ортогенеза Эймер приходит к важной идее об ограничивающей и канализующей роли конституционных особенностей организмов как результате предшествующей эволюции. Внутренняя структура организма как бы налагает запрет на разнонаправленное развитие и ограничивает его одним или немногими направлениями.
Допущение Эймером одновременного влияния внешних и внутренних факторов эволюции послужило причиной разноречивых оценок его концепции. Нам представляется, однако, что вопреки всем противоречиям и неясностям в ней явно доминирует роль внешних абиотических агентов. Внутренние имеют подчиненное значение. Такого же понимания его учения придерживались, в частности, Ю. А. Филипченко (1923, 1977) и К. М. Завадский (1973). Как мы увидим ниже, в данной части концепция Эймера полярно отличается от учения Негели.
Действие факторов эволюции, принимаемых Эймером, не ограничивается начальными стадиями эволюции. Это общие факторы как собственно видообразования, так и надвидовой эволюции, или, если можно так сказать в данном случае, факторы «сквозного» эволюционного развития. Об этом достаточно определенно говорит сам Эймер: «Подвиды и виды — это не что иное, как группы форм, стоящие на известных ступенях развития, иными, словами, — на определенных ступенях филетического роста… Это же объяснение применимо, бесспорно, и к родам как группам видов и вообще ко всем подразделениям естественной системы, которые можно обозначить вместе с Негели общим названием “родов в широком смысле” (Sippen). Поэтому в общем было бы даже лучше говорить о происхождении подобных родственных групп, а не о происхождении видов» (Eimer, 1897. S. 19).
О «сквозном» действии эволюционных факторов свидетельствует механизм возникновения видов, который Эймер специально выделяет. Обособление видов, по его мнению, происходит в силу остановки известного числа особей на какой-либо стадии общего филогенетического роста, в то время как остальные особи продолжают дальнейшее эволюционное движение. Такой способ становления видов Эймер назвал генэпистазом, что означает в переводе остановку развития рода (op. cit., S. 21). Генэпистаз не единственный, но главный путь видообразования. Обсуждая эту проблему, Эймер категорически отвергает участие в образовании s видов естественного отбора.
Итак, эволюционная доктрина Эймера, будучи в своей основе эктогенетической, фактически совмещала тезис о воздействии факторов внешней среды с канализующим влиянием организации 3 организмов и опиралась на представление, что обе эти категории причин действуют на всех этапах эволюции — от образования подвидов до возникновения высших таксонов. Хотелось бы особо отметить, что Эймер, по сути, впервые в истории эволюционной идеи обнаружил существование фактора конституционных ограничений и включил его в число механизмов, определяющих направленность филогенетических линий. Этот номогенетический аспект концепции Эймера получил в дальнейшем преимущественное развитие в трудах Вавилова, Берга и их последователей.
В противоположность представлениям Эймера, доктрина его современника немецкого ботаника Карла Негели демонстрирует приверженность автора автогенетической трактовке эволюции. Кроме того, из всех существовавших неоламаркистских концепций (исключая концепцию палеонтолога Штейнмана) она стоит ближе всего к учению Ламарка. Мы обнаруживаем в ней тот же дуализм факторов и форм эволюции, ту же трактовку конституции организмов как сущности, слагающейся из двух категорий признаков.
Отправной точкой концепции Негели служит представление о двух формах совершенства, которые он рекомендует четко разграничивать: «совершенство организации, характеризующееся сложностью строения и дифференцированностью функций», и «совершенство приспособления организмов на каждой ступени организации» (Nageli, 1884. S. 13). «Первое, — пишет Нэгели, — я назвал ввиду отсутствия другого выражения просто совершенством, второе — приспособлением» (ibid.). Совершенство первого типа можно иными словами определить как организационное, обязанное конституционным признакам. В основе его становления и развития лежит «принцип усовершенствования», предложенный Негели еще в 1865 г. Разъясняя его действие, Нэгели пишет: «Этот принцип имеет механическую природу и являет собой закон инерции в области органического развития. Поскольку развитие однажды началось, поскольку оно не может остановиться и вынуждено далее совершаться уже избранным направлением… лучше заменить слово “совершенство” более надежным словом “прогресс” (ibid., S. 12–13). Следовательно, принцип усовершенствования, no представлениям Негели, ведет организацию исключительно по пути прогрессивного развития — по пути ее усложнения. И если бы влияние этого принципа прекратилось и сохранились одни внешние воздействия, все oрганизмы остановились бы на достигнутой к этому моменту ступени организации.

Карл Вильгельм Негели (1817–1891).
Негели однозначно связывает изменение организационных признаков с внутренними причинами, заложенными в строении наследственной субстанции. Он аргументирует это положение, в частности, методом исключения влияния внешних факторов. Последние, — по мнению Негели, способны вызывать лишь обратимые (ненаследственные) приспособительные физиологические изменения, не имеющие отношения к филогенезу как необратимому и направленному процессу.
Совершенно ясно, таким образом, что Негели, как и Ламарк, разделил эволюцию организационных и приспособительных признаков, филогенез и адаптациогенез и связал их с глубоко различными причинами. Его принцип усовершенствования оказался вполне аналогичным градационному процессу Ламарка. Любопытно при этом, что Негели по сравнению с Ламарком даже усилил приоритет имманентного фактора. Он допустил возможность включения в его компетенцию формирования не только видов, но и разновидностей, выразив сомнение, что источником их образования могут служить внешние условия.
В концепции Негели есть, однако, существенное новшество. Это разработанная им умозрительная теория наследственности, согласно которой каждая клетка организма, включая половые, делится на две части — идиоплазму и трофоплазму. Только первая является субстратом наследственной изменчивости, основу которой составляют внутренние необратимые изменения, приводящие к преобразованию организации. Внешние факторы могут затрагивать идиоплазму лишь в соматических клетках, но такие изменения (Негели первым назвал их модификациями) по наследству, как правило, не передаются. Трофоплазма, служащая для питания клеток, напротив, оказывается ареной воздействия средовых факторов и ответственна за всю совокупность приспособительных признаков (подлинные модификации). Изменения этих признаков полностью обратимы и к путям наследственной передачи отношения не имеют.
Идея Ламарка и Негели о существовании у организмов двух категорий признаков в свете данных современной генетики наконец-то перестает быть мишенью для критики и выступает как одно из замечательных пророчеств. Она справедлива без особых поправок на время! Разве что удалось установить между этими категориями более четкую границу. Мы выделяем теперь инвариантные видовые, а стало быть как раз организационные, и полиморфные, т. е. приспособительные, признаки внутривидовых подразделений и вслед за Негели считаем, что последние к эволюции отношения не имеют. Многие воспринимают в наши дни аналогичное открытие генетики как нечто совершенно новое просто потому, что не знают истории. Для тех же, кто ее знает, это хороший повод вспомнить первооткрывателей и отдать должное их гению.
Но закончим наше знакомство с наследственной субстанцией в концепции Негели. По его глубокому убеждению, идиоплазма обладает очень сложным строением и наделена внутренним стремлением ко все большему усложнению. «Филогенетическое развитие, — констатирует Негели, — состоит… в том, что идиоплазма постоянно усложняется под действием внутренних причин…» (ibid., S. 181). В итоге и весь прогресс органического мира — процесс не случайный, а упорядоченный и направленный, идущий сообразно «определенному плану». Веский аргумент в пользу такого взгляда Негели не без основания усматривает в аналогии между филогенезом и онтогенезом. Нам представляется весьма вероятным, что в конечном счете, как это следует из приведенных и многих других высказываний, Негели стремился свести принцип усовершенствования, насколько это возможно, к молекулярно-физиологическим причинам.
В заключение обзора персональных концепций остановимся на многоплановом эволюционном кредо, быть может, одной из самых интересных фигур в неоламаркизме — американского палеонтолога и теоретика этого течения Эдуарда Копа.
Скажем прежде всего, что в учении Копа дуализм факторов эволюции полностью сохраняется, только внутренние, имманентные живому причины, эквивалентные градации Ламарка, он называет «силой роста», или батмизмом. В эволюции реальных групп организмов батмизм проявляется неравномерно, и соответственно, он то ускоряет, то замедляет развитие. Эту неравномерность в проявлении основного жизненного принципа Коп предложил называть законом акцелерации и ретардации. Акцелерация связана с появлением приобретенных в ходе филогенетической истории признаков на все более ранних этапах индивидуального развития. Ретардация выражается в замедлении прибавления и в конечном счете в утрате части таксономических признаков.
По представлениям Копа (Соре, 1887), акцелерация и ретардация — важнейшие механизмы эволюции, определяющие направленность развития филогенетических линий. Они действуют совершенно независимо от естественного отбора и безотносительно к потребностям адаптации. На основе акцелерации совершалась вся прогрессивная эволюция, ретардация же ответственна за регрессивную эволюцию. Для целей нашего анализа Эдуард Коп наиболее существенно, что в эво (1840–1897) люционной концепции Копа идея филогенетической акцелерации приняла своеобразную форму независимого развития родовых и видовых признаков. Их происхождение, по убеждению Копа, совершенно различно и не имеет ничего общего с дарвиновской схемой дивергенции. При этом под родовыми признаками он, как и Негели, понимал признаки конституционные, характерные и для более высоких систематических категорий, а под видовыми — приспособительные, поверхностные.

Эдвард Коп (1840-1897).
Обе категории признаков образуют в филогенезе две непрерывные, но абсолютно независимые друг от друга генеалогические линии. Коль скоро они совмещаются в едином организме, один вид может переходить из одного рода в другой, не утрачивая при этом своих видовых признаков. Образование новых родов происходит скачкообразно: «Превращение родов, — писал Коп, — могло быть быстрым и внезапным, а промежуточные… периоды постоянства могли быть весьма длительными…» (op. cit., Р. 79). Аналогичный процесс превращения признаков лежит, по мнению Копа, и в основе перехода от родов к более высоким систематическим категориям (например, подотрядам и отрядам). В этом случае утрата родовых признаков также необязательна.
В соответствии со своей концепцией Коп допускал, что одна и та же видовая форма могла существовать в последовательно сменявших друг друга родах и в различные геологические эпохи. Более того, могла возникать ситуация, при которой один и тот же вид мог одновременно принадлежать к двум и более родам, не утрачивая при этом своих видовых признаков, В этих несколько курьезных взглядах Копа, по-видимому, своеобразно преломилось влияние закона единства типа Бэра — Дарвина.
Поскольку при прогрессивной эволюции носителями более высокого уровня организации являются признаки систематически более высокой категории, допущение Копом опережающего изменения родового типа означало признание им широкого распространения в живой природе филогенетического ускорения, т. е. формирования признаков нового крупного таксона в рамках предыдущего. К примеру, новое семейство или отряд могли возникнуть, по Копу, путем присоединения нового признака к уже существующим во время эмбрионального развития их представителей. Эта блестящая идея воплотилась в наше время в одном из главных механизмов эволюционного преобразования организации, приложимых к систематическим группам животных и растений любого ранга. Но и в середине XX в. она получила широкое развитие в трудах многих палеонтологов и эволюционных морфологов.
Надо отметить, что если принцип филогенетической акцелерации носил в теоретических построениях Копа умозрительный характер, то представления о специфике механизмов эволюции на видовом и надвидовом уровнях опирались на исследования параллелизмов в формировании структурных особенностей различных групп животных (Соре, 1896). Эти исследования привели Копа к выводу, что развитие филогенетических линий происходит строго направленно и что систематические группы, начиная от родов и выше, образуются из разных корней, т. е. имеют полифилетическое происхождение.
Коп одним из первых высказал предположение, что надвидовые таксоны скорее всего возникают сальтационно и что периоды сальтаций сменяются состояниями устойчивости. Для последователя Ламарка это совершенно удивительное прозрение, если учесть, что неоламаркисты признают исключительно непрерывный и постепенный характер эволюции и сальтационизм им чужд по природе. Зато идею Копа о скачкообразном характере надвидовой эволюции приветствовали идеологи других эволюционных течений, где качественный скачок — нормативное понятие.
В воззрениях Копа есть и еще одна «оппортунистическая» черта. Оба рассмотренных нами направления в неоламаркизме опираются на механистическую трактовку движущих сил эволюции. Коп своим толкованием этих сил явно выходит за рамки и орто- и механоламаркизма и может быть отнесен к третьему направлению — психоламаркизму. Размышляя о причинах батмизма, влекущего живое по пути прогресса, Коп пришел к заключению, что таковыми могут служить только психика, сознание, воздействием которых на косную материю обусловлено само возникновение жизни. Вся прогрессивная эволюция животного мира обязана неуклонному развитию сознания, интеллекта и волевым актам — единственной силе, способной противостоять универсальному процессу деградации и разрушения. О том, так ли это на самом деле, ученые, как известно, продолжают спорить до сих пор. И здесь занимаемая ими позиция скорее определяется приверженностью определенной идеологии, чем данными науки.
Приверженцы психоламаркизма в своих эволюционных разработках опирались на ту сторону учения Ламарка, которая апеллировала к психическим проявлениям активности животных. Основателями и лидерами этого направления, кроме Копа, были немецкие биологи А. Паули, Р. Франсэ, Э. Геринг, а также русский зоолог В. А. Вагнер. В их представлениях волевые «усилия» животных, их психика и память выступили в качестве ведущей и даже единственной причины эволюции. В универсализации фактора сознания они пошли, однако, дальше Ламарка, наделив психическими способностями также растения и каждую клетку живого организма. Многие считали сознание первичным по отношению к жизни.
Психоламаркизм был частью или одной из форм витализма, позиции которого в период кризиса дарвинизма в начале XX в. особенно усилились. В книге, посвященной плачевному положению дарвинизма в современную эпоху, Франсэ, в частности, писал: «Ламаркизм отчасти тождественен с витализмом. Именно к нему относятся воззрения Бунге, Дриша, Кассовитца и Шнейдера [психический витализм]». Франсэ тут же добавлял, что «учение об одушевленности природы [ламаркизм]… впервые выставлено Ламарком» (Франсэ, 1908. С. 103, 104). Подобно другим основателям психоламаркизма, он вообще считал, что эволюция направляется не законами причинности, а свободной волей и «силой суждения», которые только и обладают формативной активностью.
Одним из идейных вождей психоламаркизма был также немецкий физиолог Э. Геринг. Согласно его ключевой идее, память является не только функцией высокоразвитого мозга, но и основным свойством живой материи. Геринг распространил способность нервных клеток к воспроизведению образов на все категории клеток, включая половые. По его мнению, унаследование приобретенных признаков основано на запоминании идиоплазмой раздражений, которые действовали на нее в прошлых поколениях.
Близкие Герингу представления развил французский зоопсихолог Пьер-Жан (Pierre-Jean, 1925). Он наделил одноклеточных и каждую клетку многоклеточных организмов «творческим сознанием», «зачаточной формой человеческой психики». В его представлении любой орган тела создается сознательным усилием его обладателя.
Психоламаркизм получил ощутимую поддержку в лице французского философа Анри Бергсона, создателя концепции «творческой эволюции» (Bergson, 1907; рус. пер, — 1914).
Даже на образованного читателя постсоветского пространства, воспитанного в духе строгого материализма, философия жизни Бергсона способна произвести шокирующее впечатление. Для него мысль о том, что «жизнь — это сознание, пущенное в материю» (там же, с. 163), — ненаучная идея, недостойная обсуждения. Ортодоксальный материалист никогда не согласится, что его идеология — разновидность предвзятой идеи. К примеру, не столь уж уникальные факты возвращения людей к жизни после клинической смерти они воспринимают как «сказку феи для взрослых». Свободный же от предвзятости ум, отталкиваясь от подобных фактов, мог бы задаться вопросом: раз «сила жизни» способна оживить только что умершего, то почему она не могла сделать некогда косную материю живой? Пора перестать открещиваться от нематериальных сил и начать их изучать, как это делают в засекреченных лабораториях.

Анри Бергсон (1859–1941).
Сущностью, основой и одновременно носителем развития и органической эволюции, придающей ей творческий характер, является, по Бергсону, нематериальный «жизненный порыв», отражающий божественную волю. Философ мыслит его как единый и общий «начальный толчок… противоположный физическим процессам». «Вся жизнь от начального толчка, который бросил ее в мир, предстанет перед ней [интуитивной философией] как поднимающийся поток, которому противодействует нисходящее движение материи» (там же, с. 240). Хотя, подобно снопу, жизненный порыв и делится на расходящиеся линии, у него есть первичное направление, он сохраняет в силу инерции единство и цельность начального импульса. Свидетельство тому Бергсон видит в явлениях параллельного развития и конвергенции, не объяснимых с позиции теории Дарвина.
Отвергая дарвинизм, теорию де Фриза, «тезис механизма» Эймера, Бергсон поддерживает психический аспект учения Ламарка, объединяемый идеей стремления живой природы к совершенствованию. Он с удовлетворением отмечает, что «из всех современных форм эволюционной теории неоламаркизм является единственной, которая может допустить внутренний и психологический принцип развития… Остается только выяснить, не следует ли тогда слово усилие брать в более глубоком, еще более психологическом смысле, чем то делает неоламаркизм» (там же, с. 69), И далее: «Мы пытались… установить на ясном примере глаза, что если здесь есть «ортогенезис», то должна привходить причина психологическая. К причине психологического порядка и прибегают некоторые из неоламаркистов. В этом, по нашему мнению, заключается один из прочных пунктов неоламаркизма» (там же, с. 78). В философии Бергсона, таким образом, была предпринята энергичная попытка восстановить старый витализм во всей его полноте.
Чтобы покончить с первоначальными течениями в неоламаркизме, коснемся еще ламаркодарвинизма, сторонники которого, не задумываясь об эклектичности получаемого сплава наук, стремились к примирению двух конкурирующих доктрин. Это представлялось возможным благодаря допущению самим Дарвином прямого приспособления на основе унаследования приобретенных признаков до момента решительного опровержения данных представлений генетикой.
Рассматривая эволюционные взгляды Жиара и Спенсера, мы уже отмечали их благожелательное отношение к участию естественного отбора в эволюции. Упоминалась и гипотеза «совпадающего отбора» Болдуина-Моргана. В первом случае, как в этом нетрудно убедиться, отбор оказывается всего лишь полезным «довеском» к самодостаточной теории.
Истории науки известна, однако, попытка более органичного объединения ламаркизма с дарвинизмом. Она принадлежала французскому биологу, одному из признанных теоретиков неоламаркизма Феликсу Ле Дантеку.
В основу своих теоретических построений Ле Дантек помещал дарвиновский естественный отбор, Он безоговорочно считал, что этот механизм выступает главной причиной адаптации в мире одноклеточных. Пытаясь выяснить механизм адаптивной эволюции многоклеточных, Ле Дантек выдвинул предположение, что клетки последних можно рассматривать как собрание тех же одноклеточных, но находящихся в крайне ограниченной среде и остро конкурирующих друг с другом. Сконструировать эту аналогию ученому помогла идея В. Ру (1881) о «внутреннем отборе», т. е. форме естественного отбора, только перенесенного на взаимоотношения клеток, тканей и частей организма: Благодаря такому отбору, действующему непрерывно, и достигаются, по мнению Ле Дантека, целостность реакции и адаптивный характер происшедшего изменения.

Феликс Ле Дантек (1869–1917).
Разъясняя смысл своей чисто умозрительной гипотезы, Ле Дантек писал, что «координация, составляющая основу живого, является исключительно результатом естественного отбора, действующего в каждый данный момент между гистологическими элементами его тела; она выражается в функциональной ассимиляции, которая укрепляет только полезные органы» (Le Dantec, 1897. P. 463). Ле Дантеку кажется, что, использовав в качестве исходного положения дарвиновский принцип, он пришел к раскрытию внутреннего содержания ламарковского механизма адаптации и тем самым добился главной цели своей научной деятельности — примирения сторонников двух различных концепций. Более того, он приходит к выводу: так как принципы Ламарка «естественно вытекают» из дарвиновских, то «учение Ламарка представляет собой, в сущности ограниченную отрасль дарвинизма» (Le Dantec, 1908. P. 201–202). Время показало, однако, иллюзорность подобного взгляда.
На рубеже XX в. неоламаркизм проник и в Россию. Из известных ученых его идеи разделяли анатом и педагог П. Ф. Лесгафт, а также географ и анархист П. А. Кропоткин. Впоследствии число сторонников неоламаркизма продолжало увеличиваться, и в 1920-е годы между ними и дарвинистами завязались острые дискуссии. Застрельщиками в них выступали биологи-ламаркисты Е. С. Смирнов, Ю. М. Вермель, Б. С. Кузин, М. В. Волоцкой, И. И. Ежиков, врач В. А. Обух. (Отражение этих дискуссий можно найти в «Вестнике Коммунистической академии» за 1927, 1928, 1931 и последующие годы, а также в сборнике «Преформизм или эпигенезис?» за 1926 г.)
Грубой пародией на ламаркодарвинизм была псевдонаучная доктрина Т. Д. Лысенко, монополизировавшая советскую биологию в период 1948–1964 гг. и по горькой иронии судьбы именовавшая себя «творческим дарвинизмом». Представления Лысенко и его единомышленников о наследственности как неделимом свойстве всей клетки, об адекватной изменчивости как результате простой ассимиляции внешних условий и их превращении в наследственную основу организма, о скачкообразном превращении одного существующего вида в другой из «крупинок» протоплазмы первого, о тождественности «вегетативных гибридов» половым и т. п. опирались на априорные натурфилософские рассуждения об обмене веществ, на ложно поставленные опыты при отсутствии генетического контроля и находились в полном отрыве отданных и методов генетики, биохимии и биофизики того времени. Это была атавистическая реверсия к XVIII в.
Расцвет неоламаркизма пришелся на первое десятилетие XX в. С началом второго десятилетия доверие к нему стало неуклонно снижаться. Одновременно менделизм и мутационизм завоевывали все большее число сторонников. Это было связано главным образом с распространением теории зародышевой плазмы Вейсмана, в свете которой наследование приобретенных признаков (НПП) казалось невозможным уже чисто теоретически. Проверка этого постулата опытным путем, на чем настаивали генетики, не давала ему решающего подтверждения.
Надо иметь в виду, что в защите НПП неоламаркисты по сравнению с генетиками находились в менее выгодном положении: в случае, если в эксперименте НПП не подтвердится, у них не останется шанса для его защиты. Генетикам подобная ситуация не угрожает: при положительном исходе проверки они всегда найдут способ его объяснить.
Менее выигрышной была и тактика борьбы неоламаркистов. Вместо того чтобы заботиться об укреплении собственной фактический базы, они на первых порах опирались в основном на критику своих противников. С другой стороны, большинство биологов не соглашались с тем, что механизм эволюции можно открыть в лаборатории, и это делало борьбу двух лагерей достаточно затяжной.
Так или иначе, в 30-е годы XX в. неоламаркизм в целом потерпел поражение, хотя и не исчез полностью. В ряде передовых стран Запада он все же сохранил по одному или по несколько своих авторитетных сторонников. Дольше всего неоламаркизм удерживал позиции во Франции. Еще в 40-е годы он пользовался здесь поддержкой большинства эволюционистов.
Теперь защитники неоламаркизма сознательно стремились к укреплению своих позиций. Для этого они настойчиво овладевали последними достижениями генетики, биохимии, квантовой химии, иммунологии, микробиологии, данными изучения цитоплазматической наследственности и симбиоза. И решение вопроса, как подтвердить дееспособность исповедуемой доктрины, было найдено!
Чтобы понять, как это произошло, нам необходимо проследить «побочную» линию развития исследований в генетике и иммунологии. Лицо генетики определялось, конечно, решающими успехами в изучении ядерного аппарата наследственности, где главная роль принадлежала школе Т. Моргана, но от этого магистрального потока отделялась едва заметная струйка, пробивавшаяся самостоятельным путем[14].
Еще в 1913 г. немецкий протозоолог В. Йоллос начал многолетнее изучение воздействия на наследственность парамеций действия высокой температуры и мышьяка. Ему удалось установить, что у значительной части популяции ответ на эти воздействия был специфически адаптивным, например вырабатывалась устойчивость к мышьяку, причем индуцированные наследственные изменения (кстати, Йоллос назвал их направленными мутациями) передавались потомству в течение сотен поколений уже после прекращения внешних воздействий. Эта первая серия экспериментов на парамециях дала Йоллосу основание высказать предположение, что и в природе естественные факторы среды оказывают на вызываемые ими мутации направляющее воздействие, благодаря чему они складываются в ортогенетический ряд, аналогичный тем, что известны из палеонтологии.
В 1930 г., сменив объект исследований на дрозофилу, он продолжил поиски ортогенетического механизма изменчивости на генном уровне. Их результатом стало заключение, что сублетальные температурные воздействия индуцируют одни и те же фенотипические изменения — вплоть до массового развития бескрылости. Наблюдавшиеся сначала специфические цитоплазматические изменения переходили в длительные модификации (Jollos, 1934).
Исследования Йоллоса не встретили понимания среди генетиков. В 1934 г. он, как и многие другие ученые, эмигрировал из фашистской Германии, и его публикации прекратились.
Со второй половины 1940-х годов облик американской генетики ощутимо изменился. В дополнение к дрозофиле в практику исследований уверенно входил новый модельный объект — микроорганизмы. В их наследственности выявился ряд сторон, не согласовывавшихся с классической концепцией гена. Складывалась особая бактериальная генетика. Многие биологи обратились к изучению загадочной цитоплазматической наследственности.
Пришло время вспомнить работы Йоллоса. Их возобновил и продолжил американский цитогенетик Трейси Соннеборн. Вскоре последовали первые открытия: возможность путем подбора условий осуществления конъюгации между разными линиями парамеций и обнаружение в цитоплазме некоторых особей генетической частицы каппа, делавшей ее носителей способными убивать других особей, ее не имевших. Перед Соннеборном открылась перспектива установления новой системы детерминации и наследования индуцированных внешними агентами признаков на уровне цитоплазмы. Важным рубежом на этом пути стало введение Г. Винклером понятия плазмагена (Winkler, 1924), названного так по аналогии с ядерными генами. В это сборное понятие Соннеборн включил также пластиды, митохондрии, кинетосомы, каппа-частицы и прочие субмикроскопические единицы, локализованные в цитоплазме. Особенно существенным оказался тот факт, что эти автономные от ядерного аппарата образования обладали геноподобными свойствами — способностями к репликации и мутированию.
Эти данные подтвердил другой американский цитогенетик К. Линдгрен (Lindegren, 1946) с женой. Изучая плазмагены (по Линдгрену, цитогены), он установил, что эти цитоплазматические частицы ответственны за ферментативную адаптацию микроорганизмов, а у дрожжей — за неменделевское расщепление. Первоначально цитогены синтезируются в хромосомах, а затем воспроизводятся в цитоплазме в присутствии субстрата, вызывающего адаптацию, уже независимо от ядерного гена. Данные экспериментов указывали на участие цитогенов в регуляции активности хромосомных генов, а возможно, и в дифференциации соматических клеток в онтогенезе.
К аналогичным выводам пришел также французский цитогенетик А. Буавэн (Boivin, 1947), изучавший направленные мутации у бактерий. Это позволило ему высказать гипотезу, по которой цитоплазматические рибонуклеиновые кислоты контролируют у многоклеточных состав цитоплазмы, а у микроорганизмов выступают носителями приобретенных признаков.
Все эти соображения и толкования, касающиеся участия цитоплазматических носителей наследственности в направленных мутациях и фиксации приобретенных признаков, были не более чем гипотезами, и механизм этих феноменов еще предстояло открыть. На помощь пришла иммунология (см.: Аронова, 2000а).
В начале 1940-х годов австралийский иммунолог Фрэнк Макферлейн Бернет (Burnet, 1941) выдвинул предположение, что антитела формируются не путем копирования антигена, как до того времени думами, но синтезируются ферментами, у которых антигены вызывают специфические адаптивные изменения. Эти адаптивные ферменты самореплицируются в антителе и передают ему специфическую информацию при его размножении. Выросшая популяция специфично адаптированных антител обеспечивает более успешную защиту организма при вторичном заражении. Правда, позднее Бернет совместно с Ф. Феннером (Burnet, Fenner, 1949) в новом варианте теории стал утверждать, что антиген вносит свою специфическую информацию в геном, а антитело уже формируется на этой вторичной матрице.
За всеми описанными событиями в цитогенетике и иммунологии внимательно следил французский эмбриолог Поль Вентребер, ревностный поклонник Ламарка. В течение ряда лет в его сознании зрела идея о том, что направленный характер мутаций можно объяснить данными этих наук и рассматривать антитела как аналог генов.

Поль Вентребер (1867–1966).
В 1949 г. Вентребер, будучи уже на 82-м году жизни и давно на пенсии, оставляет на время свой дом на берегу Средиземного моря и выступает в Парижской академии наук с докладом о «химическом ламаркизме», а в возрасте 95 лет выпускает о своей теории большую книгу под характерным названием «Живое — творец своей эволюции» (Wintrebert, 1962). Случай, прямо сказать, неординарный! Он свидетельствовал о силе убежденности этого жизнелюбивого и многоопытного старца в истинности сделанного открытия. Одновременно он показывал, сколь продуктивным может быть свежий нетривиальный подход к обоснованию интуитивной идеи. Рассмотрим оба упомянутых труда Вентребера вместе.
Опыт и широкая осведомленность в большом массиве экспериментально-биологических работ убедили Вентребера в существовании трех категорий мутаций, вызываемых совершенно различными причинами.
Первую категорию составляют мутации, индуцируемые сильными искусственными агентами типа рентгеновских лучей, высокой температуры, токсических веществ и проч. По словам Ветребера, они вызывают «прямое калечение наиболее уязвимых генов и приводят к случайным вариациям». Они носят в целом регрессивный характер и лишены всякого эволюционного значения.
Вторая категория представлена направленными мутациями, которые вызываются естественными воздействиями (например, химическими веществами) и включением чужеродных генов. Вентребер ссылается на эксперименты А. Буавэна, индуцировавшего мутации у бактерий поливом их колоний раствором, содержавшим ДНК других бактерий. В этих экспериментах подопытные бактерии интегрировали эту чужеродную ДНК в свой геном и, соответственно, изменяли свою наследственность. Явление того же порядка происходит при «вегетативной гибридизации». В обоих этих случаях, пишет Вентребер, имеет место не поломка гена, а своего рода «прививка геном», т. е. встраивание чужого генетического материала. Такие явления случаются в кризисных ситуациях, когда приобретение нового гена позволяет организму выжить и адаптироваться к изменившимся условиям. Эта категория мутаций эволюционно гораздо более перспективна.
К третьей категории Вентребер отнес мутации, вызываемые изменением климата. Это та категория наследственной изменчивости, на которую опирается традиционный ламаркизм, т. е. истинно эволюционные мутации.
Вторую и третью категории Вентребер назвал в своей книге, биологическими мутациями. На них строится эволюционный процесс.
Вентребер полностью разделял убеждение Ламарка, что причины изменения и движущие силы эволюции заключены в самих живых организмах, а именно в их жизненно важных функциях. Более того, в таком подходе к проблеме эволюции он видел основную заслугу Ламарка. В этом состоит, собственно, самая основа концепции органицизма, на которую опирается неоламаркизм в целом. Органииизм выдержал все исторические испытания и сейчас располагает прочной базой в физиологии, в науке о поведении и в новой генетике. Он неотделим не только от понимания эволюции, но прежде всего и от понятия самой жизни. Говорить о роли в эволюции активности живого и его целесообразного поведения (а это, если мы вспомним, ведущая идея позднего творчества Майра), отрицая при этом органицизм, — это все равно что учиться правильно писать, но отвергать пользу грамматики.
Признавая всю полноту гносеологической роли генетики, Вентребер в полной мере осознал, что проблема адаптивной изменчивости и передачи по наследству приобретенных признаков перешла теперь «из морфологии в область химии» и требует для своего решения отыскания соответствующих биохимических механизмов (Wintrebert, 1949). Отсюда понятно, почему Вентребер назвал свою теорию «химическим ламаркизмом».
Залогом успеха явился оригинальный подход Вентребера. Ученый уподобил эволюционный механизм процессу выработки иммунитета, т. е. представил его как ответную реакцию организма на вторжение чужеродного вещества. До сих пор многие иммунологи, включая Бернета, опирались в своих исследованиях на принцип эволюции, но Вентребер первым применил обратную процедуру — использовал теорию иммуногенеза для раскрытия эволюционного механизма. При этом логика его рассуждений была ясна: если направленная изменчивость обусловлена генетически, то следует поискать, какая система в организме в первую очередь ответственна за образование новых генов. Сходство процесса адаптации с защитной реакцией иммунной системы определенно указывала именно на эту систему.
Представление об иммунитете издавна связано с существованием антител (ныне их чаще именуют иммуноглобулинами), образующихся в организме позвоночных животных в ответ на проникновение в него чужеродных тел — антигенов. Последние могут быть представлены различными токсинами, болезнетворными микробами, вирусами и даже собственными клетками с измененной поверхностной мембраной. Функция антител состоит в распознании и обезвреживании антигенов. Чтобы антитела могли ее выполнять, организм должен обладать способностью создавать строго специфические антитела, точно соответствующие структуре антигена.
Вентребер предположил, что отправным моментом эволюции служит наступление кризисных условий среды. Такие условия нарушают нормальную работу внутренних органов, и они начинают вырабатывать какие-то вредные вещества, которые воспринимаются организмом как антигены. На появление подобных «антигенов функциональной недостаточности» (теперь их называют стрессорами, а породившую их ситуацию — стрессом) иммунная система. отвечает созданием в лимфоидной системе новых генов — производителей соответствующих антител. Новые гены, встраиваясь в. хромосомы половых клеток, изменяют наследственность вида и прежде всего как-то перестраивают ген, управляющий работой; внутренних органов (фактически тот же механизм лежит в основе позже открытой вирусной трансдукции). Происходит биологическая мутация, направленная на исправление функциональной недостаточности. Таким образом, в отличие от самопроизвольных, ненаправленных мутаций, принимаемых в СТЭ, биологическая мутация у Вентребера — это изначально адаптивное наследственное изменение. Получается, что организм сам активно (но без участия воли) производит свои мутации. И на такого рода мутациях, по Вентреберу, основана адаптивная эволюция.
Вентребер воспринял представление Соннеборна о двойственной природе гена — его способности существовать как в составе ядерного генетического аппарата, так и внутри цитоплазмы — в форме РНК-содержащих плазмагенов. Как эмбриолог, убежденный в организующих свойствах цитоплазмы, Вентребер допускал возможность синтеза в ней новых генов и их «доработки» до соответствия строению антигена. Плазмаген как-то узнает это строение и передает о нем информацию в клеточное ядро, т. е. фиксирует ее наследственно. Ген для Вентребера — всего лишь «продукт, сотворенный живой материей, ее посланник в хромосомах… сохраняемый и используемый, где и когда понадобится» (Wintrebert, 1962. Р. 318).
Представления Вентребера опередили свое время. Они абсолютно не вписывались в центральную догму генетики, не допускавшую самой возможности воздействия цитоплазматических изменений на генотип. Поэтому нет ничего удивительного в том, что идеи Вентребера не встретили понимания среди биологов. Они могли быть восприняты только после устранения абсолютного характера этой догмы.
Всего несколько лет отделяют смерть Вентребера (1966) от открытия РНК-содержащих вирусов, способных осуществлять обратную транскрипцию и встраиваться в геном клетки-хозяина, и 16 лет — от нематричного синтеза, раскрывшего способ образования нового гена в цитоплазме, который так интересовал Вентребера.
В 1971 г. Темин (Temin, 1971) высказал гипотезу, что в экстремальных условиях ретровирусы могут заражать половые клетки, и, внеся в них чужеродную генетическую информацию, превращаться в эндогенную форму, наследуемую совместно с геномом половых клеток. В дальнейшем они становятся неотличимыми от исконных клеточных генов.
В 1982 г. в области иммуногенеза был открыт нематричный синтез ДНК: цитоплазматические ферменты «разрезают» старые гены на части и при «сшивании» нового (кодирующего иммуноглобулин антитела) встраивают между этими частями небольшой фрагмент (он состоит из нескольких нуклеотидных пар), синтезируемый ферментативно, без участия генетической матрицы. В середине 1980-х годов был раскрыт и современный механизм выработки иммунитета, складывающийся из трехэтапного клонирования генов иммуноглобулинов и их высокочастотного направленного мутирования. Вся эта ступенчатая «доработка» гена, как предугадал Вентребер, совершается в цитоплазме.
Наконец, в 1988 г. американский генетик Дж. Кэйрнс экспериментально доказал существование направленного мутагенеза. Указанные открытия способствовали быстрому возрождению ламаркизма (подробно об этом см. в гл. 14).
Итак, концепция Вентребера находилась в русле или предвосхитила несколько будущих фундаментальных открытий: обратную транскрипцию, нематричный синтез ДНК, горизонтальный перенос информации, генетически обоснованное НПП. Идея о существовании направленного мутагенеза обрела прочное экспериментальное обоснование.
Как уже говорилось, Вентребер назвал свою концепцию «химическим ламаркизмом», поскольку в ней были предложены современные биохимические механизмы ламарковской адаптации. По сути, как мы только что видели, Вентребер исходил в ней из физиологических и иммунологических представлений, и, вероятно, его концепцию точнее можно было бы назвать «физиологическим» или «иммунологическим ламаркизмом».
Вентребер был первым, кто привлек иммунологию к решению эволюционных проблем. Он первым уловил логическую возможность сближения учения об эволюции с иммунологией под главенством ламарковской идеи активной адаптации. Это стало возможным благодаря определенным успехам иммунологии и нетрадиционной генетики. Об объективности наметившейся тенденции свидетельствовало появление новых последователей старой доктрины с их новыми гипотезами.
Одним из них стал молодой австралийский иммунолог Эдвард Стил, который сразу заявил о себе публикацией книги о соматическом способе НПГТ (Steele, 1979). Е. А. Аронова (1977), которой мы обязаны знакомству с ее содержанием, отмечает, что Стилу не были известны труды Вентребера и что в поисках ответа, может ли новая генетическая информация возникать в самом организме, он шел самостоятельным путем. При этом, в отличие от Вентребера, он положил в основу своей гипотезы так называемую клонально-селекционную теорию образования антител. Согласно данной теории антитела, формирующиеся в организме в ответ на вторжение чужеродных агентов, выступают в качестве рецепторов, с которыми эти агенты избирательно взаимодействуют. Как только взаимодействие установлено, запускается механизм клональной пролиферации (размножения) клетки антитела, дающий множество клеток, достаточно специфичных к данному антигену. Небольшая часть клеток из возникшего клона в иммунологической реакции не участвует. Она образует самоподдерживающуюся популяцию, ответственную за иммунологическую память, или иммунитет. За пролиферацией следуют этап интенсивных тонковых мутаций в клональных клетках и селекция возникших мутантов. Мутанты с более полной комплементарностью к антигену обретают возможность преимущественного размножения.
Стил предположил, что клональный отбор — универсальный механизм, лежащий в основе работы не только иммунной, но и любой другой системы организма. Так, если в соматической клетке случается мутация, которая может быть полезна в изменившихся условиях среды, то потомки именно этой клетки, согласно гипотезе Стила, будут по преимуществу слагать и поддерживать соответствующую ткань. Стил вообще полагает, что эволюционно значимые мутации происходят в первую очередь в соматических клетках, а клональная селекция обеспечивает их массовое размножение. Это резко повышает вероятность того, что мутантный ген будет захвачен ретровирусами, перенесен в половые клетки и вставлен в их генетический аппарат.
Поначалу реакция на гипотезу Стила была резко критической, но очень скоро за ее публикацией последовал ряд только что отмеченных фундаментальных открытий и экспериментов, которые автоматически ввели ее в «русло времени».
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК