Глава 11. Номогенез
Этот термин (от греч. «nomos» — закон) впервые вошел в биологию благодаря изданию Л. C. Бергом одноименной книги с полным названием «Номогенез, или эволюция на основе закономерностей» (1922). С тех пор им стали обозначать прогрессионистские эволюционные гипотезы, авторы которых рассматривают эволюцию как запрограмированный процесс реализации внутренних, имманентных живому организму закономерностей. В своей исходной форме номогенез противопоставлялся теории Дарвина как основанной исключительно на случайной изменчивости и потому названной тихогенезом.
Руководствуясь априорными суждениями в духе кантианского агностицизма, Берг провозгласил основным законом эволюции «автономический ортогенез» — имманентное свойство живой природы производить независимо от внешней среды все более совершенные формы. Постулируя наличие у организмов такого целенаправленного внутреннего процесса, или силы, Берг указывает, что нечто аналогичное уже было высказано в России К. Бэром, Н. Я. Данилевским и Н. Н. Страховым, а на Западе — К. Негели и Э. Копом.
Концентрируя свое внимание главным образом на прогрессивной эволюции и случаях «истинного новообразования», Берг честно признает, что о причинах прогресса ему ничего не известно. Зато о способах его осуществления он пишет с полной определенностью. Согласно его номогенетической концепции, автономические факторы изменяют «существенные признаки, определяющие самый план строения данной группы» (там же, с. 182), и ведут ее по пути прогресса. В итоге возникают новые органы и образуются систематические группы от уровня вида до класса, причем Берг специально подчеркивает, что соответствующие признаки часто «образуются в определенном направлении <…> независимо от пользы… а иногда — даже во вред организму» (там же, с. 179).
По мнению Берга, запрограммированность эволюционного развития филогенетических линий органически включает в себя явление преадаптации. Наряду с автономическим ортогенезом ученый обосновывает его ссылками на преобладание в эволюции параллелизмов и конвергенций и иллюстрирует их искусно подобранными примерами.
Источник прогрессивных преобразований Берг усматривает в стереохимических свойствах белков, т. е. в изменениях их пространственной структуры, побуждающей формы изменяться только в определенном и всегда целесообразном направлении.
Как же представлял себе Берг механизм возникновения новых признаков, а вместе с ними — новых видов и более высоких таксонов?
Во всех случаях это процесс, одновременно охватывающий всех особей данного вида и сразу на громадной территории. При его описании Берг пользуется такими эпитетами, как эпидемический, массовый, стихийный, и многократно возвращается к этому вопросу.
Тезис о массовом характере преобразования населения вида Берг настойчиво противопоставляет положению Дарвина о действии естественного отбора на индивидуальную изменчивость и преимущественное сохранение первоначально небольшого числа изменившихся индивидов. Изменчивость, лежащая в основе образования новых признаков, никогда не бывает случайной. Она всегда возникает закономерно, т. е. в нужное время, и направлена в сторону, полезную для ее обладателей. В этом, по Бергу, «и заключается вся соль вопроса об эволюции: получается ли полезное случайно или закономерно?» (там же, с. 180). К этому следует добавить, что в полном отрицании случайности и, напротив, в утверждении строгой закономерности преобразования как раз и состоит идейное ядро теории номогенеза. Но коль скоро изменчивость закономерна и направленна, нужда в естественном отборе как факторе эволюции полностью отпадает.
Массовые преобразования форм, связанные с возникновением новых признаков, могут осуществляться не только под действием автономических процессов, но и под влиянием географического ландшафта, т. е. причин хорономических. В этом случае новые формы возникают в результате географической изоляции части популяции вида, испытывающей «принудительное» превращение.
Влияние географического ландшафта по сравнению с действием автономических причин намного скромнее по результатам, так как оно способно приводить лишь к образованию подвидов, в лучшем случае — викариирующих видов. Соответственно, Берг уделяет этому фактору гораздо меньше внимания.
Отдавая приоритет причинам автономическим, Берг разъясняет, что их действие проявляется в так называемых мутациях Ваагена[27] — морфологических изменениях, происходящих во времени» которое разделяет соседние геологические горизонты. При этом новые виды образуются путем замещения (субституции) старых, материнских на основе «массового преобразования» громадного количества особей, а не путем дивергенции, как это мыслится Дарвином. Более того, массовое тиражирование нового признака только и есть гарантия его прохождения сквозь бдительные сети отбора и его наследственного закрепления. Сами же мутации совершаются исключительно скачками, почему между истинными таксонами никогда не бывает переходных форм. Берг отмечает также, что «массовое преобразование есть явление геологического порядка: оно связано с изменением фауны данного горизонта и происходит в известные промежутки времени, чтобы затем опять на долгое время прекратиться.
Это и есть путь прогрессивной эволюции» (курсив мой. — В. Н.) (там же, с. 317).

Лев Семенович Берг (1876-1950).
Что касается масштаба возникающих новообразований, то он бывает самым различным, но главное, что это часто «резкий и заметный шаг… в морфологическом отношении: это может быть образование плаценты, конечности типа пятипалой, конечности типа летающей, появление гетеростилии, семени, двуполого цветка и т. п.» (там же, с. 329), Таким путем возникают, по Бергу, новые роды, отряды и даже классы. Способ образования высших таксономических единиц такой же, как и низших. При этом прогрессивные изменения совершаются впервые в молодом возрасте или в эмбриональном состоянии.
В противоположность мутациям Ваагена, т. е. филетическим преобразованиям, мутации де Фриза, по мнению Берга, никакого видообразовательного и вообще эволюционного значения не имеют, ибо они появляются у единичных индивидов, а их носители — мутанты — обычно образуются путем утраты генов. На подобных мутациях основывать прогрессивную эволюцию невозможно.
Преобразования одних форм в другие происходят периодически, скачками. Есть эпохи, пишет Берг, когда творческая сила природы дает калейдоскоп органических форм, а есть времена, когда эта сила как бы дремлет. Внешнее выражение такого хода эволюции мы видим в самом делении геологической истории на эры, периоды и эпохи. Если проследить последовательность родственных родов и видов, замещавших друг друга исторически, то, как бы ни были полны палеонтологические данные, история всегда оказывается прерванной. Берг склонен считать скачкообразность законом прогрессивной эволюции и меняет смысл известного афоризма Лейбница на противоположный: «Природа делает скачки».
Представления Берга о судьбе внутривидовых подразделений прямо противоположны дарвиновским. Предваряя идеи многих современных макрогенетиков, он полагал, что эти низшие внутривидовые единицы никогда не в состоянии «дорасти» до вида в результате дивергенции, а, наоборот, виды, возникнув сразу, скачком, разделяются на подвиды и более мелкие единицы. В современную нам эпоху, отмечал Берг, вполне можно наблюдать разложение сборного (линнеевского) вида на его составные элементы, но никто еще не видел обратного процесса — превращения расы в вид путем подбора, и нельзя допустить, чтобы такой процесс мог происходить. Следовательно, по Бергу, сначала образуется вид, а уже затем происходит его расщепление на соответствующие внутренние составные части. В обратную сторону процесс не идет.
В соответствии с представлением о первичном разнообразии форм жизни и их параллельном эволюционном развитии Берг отстаивает идею крайней полифилии и противопоставляет ее дарвиновскому принципу дивергенции. Соответственно, филогенетические отношения между систематическими группами представлялись ему не в форме ветвистого дерева, а в виде ржаного поля. Интересно, что, подобно голландскому генетику Я. Лотси — представителю совершенно иного эволюционного направления, — Берг допускал возможность политопного и повторного образования видов и высших таксонов и верил в частичную обратимость эволюции.
Но вернемся к стержневой идее «Номогенеза…». Берг сам ясно определил ее выбором одного из эпиграфов к главе об определенном направлении, или — закономерности, эволюции. «В области органической природы, — говорится в нем, — точно так же, как и в области неорганической природы, случайность отсутствует, и полезность многих деталей тела возникает по законам, а не в результате случайностей или случайных событий» (Osborn, 1909. Р. 225).
Благодаря единым законам развития «эволюция идет в определенном направлении», по конкретному руслу, подобно электрическому току, распространяющемуся вдоль проволоки (сравнение Берга). Она складывается из направленного (а отнюдь не хаотического) изменения признаков организмов. Варьировать же в определенном направлении организмы побуждают главным образом внутренние, автономические причины.
Направленность эволюции всего нагляднее проявляется в явлениях конвергенции (параллелизма), при которых у двух или более рядов форм развиваются сходные признаки, поскольку эти явления вызываются «наследственной склонностью варьировать в одинаковом направлении».
Берг особо отмечает, что конвергенция — не исключительное явление, как думал Дарвин, а «основной закон эволюции органического мира» (там же, с. 228). В силу этого закона сходство между организмами может быть не только результатом кровного родства, но следствием более общего принципа — развития живого по одинаковым законам. Поэтому Берг не делает принципиального различия между гомологией и конвергенцией.
Книга Берга насыщена примерами конвергенции как родственных, так и далеких друг от друга групп организмов, относящихся и к животному, и к растительному миру. Фактически вся сравнительная анатомия могла бы, по его мнению, служить в этом отношении иллюстрацией. Среди позвоночных параллельное развитие демонстрируют эволюция зубов у рептилий и млекопитающих, постепенное окостенение позвоночника у высших рыб, уменьшение числа костей в черепе, превращение сердца из двухкамерного в трех- и четырехкамерное (последнее развилось совершенно независимо у крокодилов, птиц и млекопитающих). По целому ряду внешних и внутренних признаков обнаруживается сходство ихтиозавров с дельфинами, хищных птиц с совами. Совершенно удивительно одинаковое устройство органа зрения у кольчатых червей, членистоногих, головоногих моллюсков и позвоночных. Такой орган, как плацента, кроме соответствующего инфракласса млекопитающих имеется у ряда мшанок, у некоторых насекомых и скорпионов, у оболочников, у акулы Mustelus lаеvis, а также у некоторых сумчатых. Природа трижды сделала попытку создать формы с автостилическим черепом среди позвоночных, а именно у химер (рыбы), у двоякодышащих и, наконец, у четвероногих.
Большое число параллельных рядов форм позволяет построить также палеонтологический материал. Они широко известны среди гониатитов, аммонитов, паплюдин, динозавров, теридонтов, лошадиных, между птерозаврами и птицами, крокодилами и птицами и т. д.
О закономерной направленности эволюции свидетельствуют также явления филогенетического ускорения или предварения признаков, а также закон гомологических рядов в наследственной изменчивости, установленный Н. И. Вавиловым (1920; Вавилов, 1968; Vavilov, 1922). Говоря о последнем, Берг замечает, что «Вавилов проводит идею номогенеза более успешно, чем это делаю я в настоящей работе» (там же, с. 224), а также многократно ссылается на его данные.

Николай Иванович Вавилов (1887-1943).
Сущность закона гомологических рядов совершенно ясна в той формулировке, которую ему дал сам автор:
«1. Виды и роды, генетически близкие, — писал Вавилов, — характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и линнеоны, тем полнее сходство в рядах их изменчивости.
2. Целые семейства растений, в общем, характеризуются определенным циклом изменчивости, проходящей через все роды и виды» (Вавилов, 1968. С. 32).
Данный закон основан на анализе громадного материала по изменчивости культурных растений и их диких сородичей и никогда никем не опровергался. Вавилов привел удивительно правильные параллельные ряды форм, как в пределах одного рода (у пшеницы, ячменя, овса, пырея), так и у разных родов (ржи и пшеницы, разных представителей бобовых и тыквенных).
Особенно нагляден пример поразительного сходства разных родов — культурной чечевицы (Ervum lens) и часто засоряющей ее посевы плоскосемянной вики (Vicia saliva). Оба растения одновременно цветут и созревают, а их семена по размеру и форме настолько похожи друг на друга, что сортировочные машины не в состоянии их разделить. Хотя роль искусственного отбора в данном случае несомненна, сами формы с одинаковыми семенами были обнаружены в природе, где они образовались в полном соответствии с законом изменчивости до и помимо всякого отбора.
В свете закона Вавилова особенно поблекла значимость явлений мимикрии как комплекса адаптивных внешних признаков, имитирующих внешность хорошо защищенных видов, и как приспособления, якобы возникшего под действием естественного отбора. Вавилов и Берг высказываются по этому поводу в унисон: первый видит в случаях мимикрии «повторение циклов изменчивости в различных семействах и родах», а второй утверждает, что «явления мимикрии… целиком могут быть подведены под понятие гомологических рядов» (Берг, 1977. С. 313). В цитируемой книге Берг собрал много примеров (главным образом среди бабочек) бесполезности мимикрии, когда имитаторы подражают сразу многим видам других семейств, когда имитатор и модель никогда не видели друг друга, так как обитают на разных континентах. Опираясь на закон Вавилова, полное развенчание дарвиновского толкования миметизма осуществил австрийский энтомолог Ф. Гейкертингер (Heikertinger, 1954).
Дарвинисты, всегда яростно критиковавшие Берга, в отношении закона Вавилова часто использовали тактику замалчивания и уж во всяком случае редко вспоминали, что, наряду со сходной изменчивостью видов, родов и семейств, Вавилов признавал существование у них специфических, до поры неизменных признаков — радикалов. К числу таких радикалов относятся, в частности, величина и кратность хромосомных наборов. У пшеницы, например, она обычно кратна семи (у однозернянок — 14, у твердых — 28, у мягких — 42). Понятно, что подобный радикал, как и вообще любой количественный признак, не может возникнуть постепенно, как — обычно принято считать — развиваются качественные признаки под действием отбора. Он мог образоваться только сразу, одномоментно, например в результате гибридизации или геномной мутации.
Закон Вавилова приобрел не только универсальное общебиологическое значение. Он оказался одним из самых практичных теоретических обобщений генетики. Подобно периодическому закону Менделеева, позволившему целенаправленно искать в природе еще не открытые химические элементы, он создал ориентиры для поисков доселе неизвестных видов и разновидностей, — могущих оказаться полезными в селекции тех или иных культур. В частности, широко известен пример обнаружения Вавиловым наперед им предсказанных форм ржи без лигулы у основания листовой пластинки и с опушенными колосьями, совершенно аналогичных соответствующим формам пшеницы. Им же было найдено так называемое голое просо. Многие другие формы растений и целые гомологические ряды были обнаружены сотрудниками, учениками и последователями Вавилова, способствуя тем самым популяризации его закона. Последний стал известен и западному научному сообществу благодаря публикации в 1922 г. статьи Вавилова в «Journal of Genetics» (Vavilov, 1922). Его с одобрением восприняли многие крупные генетики. Он послужил также толчком для разработки А. А. Заварзиным (1923; см. также: 1986) концепции параллелизма в филогенетическом развитии тканей.
Вавилов (как, впрочем, и многие интерпретаторы его закона) писал, что закон гомологических рядов «не противоречит дарвинизму, наоборот, развивает его» (Вавилов, 1939. С, 519). Одним из доводов в пользу такого толкования служило предположение о проявлении действия гомологичных генов, унаследованных от общего предка, аналогичного тому, которым для объяснения параллельной изменчивости пользовался Дарвин. Кроме того, Вавилов допускал возникновение фенотипических сходств в результате действия разных аллелей одного гена и даже разных генов, что было подтверждено уже через полвека данными молекулярной биологии при изучении так называемых неполных и ложных гомологий на уровне генотипа (см., например: Медников, 1980, 1981, 1983).
Сделаем маленькое отступление. Как Вавилов установил свой закон для фенотипических признаков, так и Дарвин, естественно, описывал случаи аналогичной изменчивости по видовым признакам, т. е. оба этих явления относятся к морфологическому уровню организации. Современная молекулярная генетика с еще большей убедительностью, чем классическая генетика времен Вавилова, показала самостоятельность, или глубокую автономию, процессов формообразования от генетических факторов. Во всяком случае, такое впечатление будет сохраняться до тех пор, пока фенетика не воссоздаст всю цепочку событий от гена (или генов) до интересующего нас морфологического признака. А поэтому не стоит полагать, что доказательство плейотропной или полигенной детерминации фенотипических сходств может снять противоречие между направленной изменчивостью у Вавилова и ненаправленной — у Дарвина. Это противоречие остается реальным фактом. Поэтому абсурдно одновременно принимать закон Вавилова и осуждать закон конвергенции Берга.
Впрочем, в условиях уже сложившегося в СССР в 1930-е гг. тоталитарного строя и жесткого идеологического контроля даже крупные биологи могли писать одно, а думать другое. Подгонять же под материалистическую диалектику и дарвинизм новые открытия считалось обязательным требованием лояльности. Тем более значимым для нас становится непредвзятое суждение А. А. Любишева, считавшего, что с открытием закона гомологических рядов Вавилов сделал «очень крупный шаг по пути проникновения в закономерности систематики и эволюции», который «не гармонировал <…> с общей системой дарвиновских взглядов на эволюцию…» (Любищев, 1982. С. 248, 252–253).
Эволюционной концепции Берга близки взгляды палеонтолога Д. Н. Соболева. Подобно Бергу, он признавал существование закона автономического ортогенеза, считая его выражением автономного и имманентного живому свойства изменяться во времени. Но в отличие от Берга, который его только постулировал, Соболев стремился найти ему прочное обоснование на ископаемом материале.
Развивая свою теорию биогенеза, Соболев (1924), как и Берг, считал, что филогенетические превращения, ведущие к возникновению высших таксонов, осуществляются преимущественно сальтационным путем. Порождаемую сальтациями прерывистость эволюции он даже провозгласил законом биогенеза. При этом, следуя заде Фризом и Бэтсоном, Соболев полагал, будто сальтации сводятся, как правило, к утрате наследственного фактора и поэтому направляют эволюцию в сторону регресса.

Дмитрий Николаевич Соболев (1872–1949).
Сближает Соболева с Бергом и представление, что случающиеся филогенетические превращения организмы испытывают уже при своем рождении. К этому Соболев добавляет, что «подобно индивидуумам, и высшие органические единицы, очевидно, также родятся, они возникают благодаря более или менее глубокому превращению или филогенетическому метаморфозу» (там же, с, 171). Соболев допускал и частичную обратимость эволюции, утверждая, что почву для сальтационистских превращений, «по-видимому, всегда готовило обратное развитие или омоложение» (там же, с. 164).
Однако нас больше интересует другая, более ранняя работа Соболева, посвященная исследованию закономерностей филогенетического развития девонских головоногих моллюсков — гониатитов (Соболев, 1913). Опубликованная девятью годами ранее «Номогенеза» Берга, эта работа в ряде моментов, относящихся к особенностям параллельной изменчивости, опережала Берга концептуально и предвосхищала разработки современных его последователей.
Коснемся прежде некоторых понятий, которыми оперирует Соболев. Согласно его представлениям, существуют три рода эволюционных изменений: изменения комбинационные, градационные и мутации де Фриза. Последние характеризуются внезапностью, но их формообразовательная роль ограниченна, и Соболев их не рассматривает.
В центре внимания ученого, безусловно, градационные изменения[28], которые представлены уже упоминавшимися нами мутациями Ваагена — морфологическими изменениями в разрезе геологического времени (современные исследователи чаще называют их хроноклинами). Несколько последовательных мутаций образуют ряд, или линию.
Вообще, ряд как таковой можно построить только по одному признаку. Но в палеонтологии, имеющей дело с биологическими объектами и, следовательно, с коррелятивной изменчивостью, возможны ряды по двум или нескольким признакам (как, например, в случае ряда копытных). Фактически все параллельные ряды форм, приводимые Бергом, построены по одному признаку.
Соболев сознательно анализирует сочетания или комбинации признаков. Он, очевидно, полагал, что если существует направленная и параллельная изменчивость, то в одной линии одновременно может быть несколько параллельных рядов — для каждого признака свой. Так, он пишет, что, взяв две пары признаков, мы получим четыре их возможных сочетания попарно и столько же больших групп гониатитов. Теперь можно проследить их градационные изменения. Сам Соболев проследил преобразования немногих признаков морфологии раковины и обнаружил, что изменения в различных линиях и группах линий происходят в одинаковом порядке — через те же градации и стадии, что и в других. При таком параллельном развитии целый ряд «видов» какого-либо «рода» может претерпеть подобные и одинаково направленные изменения и, таким образом, приобрести признаки другого «рода» (там же, с. 10).
Параллельно развивающиеся линии не обособлены друг от друга. Время от времени они скрещиваются, и тогда комбинации признаков соседних рядов (линий) соединяются промежуточными комбинациями, состоящими из признаков обеих линий. При многократном соединении (анастомозах) перекрещивающиеся линии образуют сложную сеть родственных связей, которую Соболев называет сетью скрещивания. Таким образом, пишет Соболев, мы приходим к представлению о «сетчатом строении органического мира», поскольку последний «состоит из комбинаций» (там же, с. 85).
Соболев обнаружил также, что один и тот же признак у форм, живущих одновременно, может быть представлен в нескольких вариациях. Носителей таких вариаций Соболев назвал изомерами. Примером может служить такой важный систематический признак, как характер сутурной (лопастной) линии. Ее видоизменения при одинаковом числе лопастей связаны с их разной локализацией. В ходе эволюции получаются изомерные стадии сутуры. Соболев установил четыре независимых параллельных ряда по этому признаку. Наконец, Соболев пришел и к идее отображения эволюционного многообразия параллельных рядов в виде таблицы, как это предлагают современные номогенетики. Вот как он говорит об этом сам: «Если комбинации с одинаковыми формулами [признаков] расположить в вертикальные ряды по стадиям сутуры и притом таким образом, чтобы одинаковые стадии расположились горизонтальными рядами, тогда место пересечения вертикального ряда с горизонтальным точно определяет положение комбинации (или группы комбинаций)». Он добавляет, что так мы получаем «более точную и естественную систему». В приводимых таблицах гониатитов и климений Соболев располагает комбинации каждой линии по градациям и стадиям сутуры.
Исследования параллельной изменчивости после открытия закона Вавилова, столь многочисленные в 1920-е годы, к началу 1940-х годов полностью прекратились. Об этом законе и номогенезе Берга забыли на целых два десятилетия. К проблеме вернулись только в начале 1960-х годов, причем в западных публикациях имена Вавилова и Берга чаше всего уже не упоминались[29].
Особняком стоит французский зоолог-биоспелеолог Альбер Вандель, уже знакомый нам как типичный представитель финализма. Можно сказать, что по всем ключевым характеристикам развиваемой им эволюционной концепции (Vandel, 1948, 1949, 1963, 1968) он солидарен с Бергом. Это касается и идеи авторегуляции, родственной автономическому ортогенезу Берга, и филогенетического преформизма, и представлений о единстве законов онто- и филогенеза, о массовом и приспособительном характере изменчивости, делающей излишним вмешательство отбора, об исключительно сальтационном формообразовании и о том, что эволюция начинается с преобразования типов организации (подробнее см.: Назаров, 1984).
Интерес к закономерностям эволюции определялся у разных авторов разными причинами, но наиболее общими, по-видимому, были затруднения в истолковании параллелизмов, направленности и телеономичности эволюции с позиции теории отбора (СТЭ), выявление эволюционной значимости активности организмов, способности живого к самоорганизации, склонность искать объяснения сложных вопросов упорядоченности эволюции в рамках определенных философских и научных традиций (например, приверженность многих французских эволюционистов гипотезе антислучайности, разработанной в 30-е годы Л. Кено).
Заслуживают упоминания (а некоторые и более подробного рассмотрения) четыре концепции. Причины канализованности и телеономичности эволюции в них связывают с увеличением количества ДНК (С. Оно), со структурой белков (Ж. Моно), с процессами трасформаиии, заложенными в организации вещества и энергии (А. Лима-де-Фария), с законами системной общности всех объектов природы (С. В. Мейен, Ю. А. Урманцев).
Американский молекулярный биолог Сусуму Оно попытался связать грандиозные макроэволюционные события с многократной дупликацией генов и увеличением на этой основе количества. ядерной ДНК (Ohno, 1969; Оно, 1973). Путем дупликации избыточных (нефункционирующих копйй) генов возникают новые — как структурные, так и регуляторные — гены, ответственные за крупные морфологические преобразования. Это так называемые тандемные дупликации, которые влекут за собой также пропорциональное увеличение размеров клеток тела и делают невозможной дальнейшую функциональную дивергенцию вновь возникающих копий. Поэтому для обеспечения нормального хода макроэволюции тандемные дупликации должны были чередоваться с полиплоидией — дупликацией всего генома.
По мнению Оно, по достижении предками амниот уровня организации рептилий с высокоспециализированным механизмом определения пола «великий эксперимент природы с дупликациями генов» должен был прекратиться, о чем свидетельствует относительная стабилизация размеров генома (у змей и ящериц он колеблется в пределах от 60 до 67 %, а у птиц — от 44 до 59 % размера генома плацентарных млекопитающих). Адаптивная радиация млекопитающих происходила уже без заметного числа дупликаций — только за счет ранее накопленных копий генов. Избыточные копии, не используемые в настоящее время, могут оказаться полезными в будущем при новых обстоятельствах.
С момента публикации работ Оно был установлен ряд фактов, нарушающих постулированную закономерность. Помимо того что у некоторых рыб и земноводных обнаружилось количество ДНК, в 25 раз превышающее ее содержание у любого из видов млекопитающих (Уотсон, 1978. С. 507), фактически в пределах любого крупного таксона этот показатель обнаруживает большой разброс. Даже в пределах одного семейства, а также рода количество ДНК разнится в несколько раз: у разных видов дрозофил — в 2,5 раза, у близких видов злаков — в 3 раза, а у лютиков — в 5 раз (Голубовский, 2000. С. 81). Кроме того, выяснилось, что в состав ДНК входят фракции из многократно повторяющихся последовательностей, не кодирующих никаких полипептидных цепей, на долю которых приходится до 80–90 % генома. Поэтому находится немало специалистов, которые вообще отрицают какую бы то ни было корреляцию между прогрессивной эволюцией и величиной генома (Корочкин, 1985).
Французский биохимик Ж. Моно (Monod, 1970) наделил телеономическими свойствами белки и свел к их принципам организации все характеристики сложного многоклеточного организма, почти буквально повторив Берга. Подобные свойства белков зависят, по Моно, от их способности вступать в стереоспецифические взаимодействия нековалентного характера с другими соединениями. Отсюда он сделал вывод, что структурная самосборка организма в онтогенезе и изменения в ней, переходящие в филогенез, представляют собой суммарный итог самопроизвольной организации белков, информация о которой заключена в их структуре. К идее направленности и неслучайности эволюции приходят и многие другие зарубежные биологи разных специальностей (Whyte, 1965; Riedl, 1978; Taylor, 1983).
Среди современных номогенетических толкований эволюции выделяется своим радикализмом концепция автоэволюции шведского цитогенетика испанского происхождения Антонио Лима-де-Фариа (1991). Она в равной мере отражает идеи западного структурализма и глобального эволюционизма. По последовательности проведения принципов последнего она стоит в одном ряду с построениями Берга, Тейяра де Шардена и Янча.
В понимании А. Лима-де-Фариа биологическая эволюция — всего лишь продолжение эволюции физико-химической, которая началась с рождением Вселенной. В этой предбиологической эволюции он выделяет три последовательных автономных уровня — эволюции элементарных частиц, химических элементов и минералов. Присущие последним законы и правила задают все особенности биологической эволюции и прежде всего налагают на нее все новые ограничения, пока не останется всего один или несколько возможных путей ее реализации. Эта стержневая идея Лима-де-Фариа проходит лейтмотивом через всю его книгу.
Согласно общепринятой точке зрения канализация эволюционного развития есть следствие ограничений, накопленных организацией биологических объектов в ходе предшествовавшей эволюции. Новизна концепции Лима-де-Фариа заключается в том, что ограничения, по его мнению, возникают на трех низших уровнях организации и биологическая эволюция их только отражает. Это положение он пытается продемонстрировать на примерах бесчисленных сходств.
Нельзя не согласиться с тем, что высшие формы движения материи, включая в себя низшие, не затушевывают и не отменяют действующих в них закономерностей. Так, если белки при нагревании свыше 45–50°C испытывают денатурацию, то и организмы, тела которых построены из белков, погибают при тех же температурах. Ток, вырабатываемый электрической батарейкой, представляет собой поток ионов, возникающий в электролите в результате окислительно-восстановительной реакции. Электрические потенциалы, образующиеся на биологических мембранах, в клетках и специальных электрических органах у ряда рыб, имеют ионную природу.

Рис. 14. Листовидные структуры. А. Минерал: чистый висмут в самородной форме (Medenbach, Sussieck-Fornefeld, 1983). Б. Растение: лист сумаха (Feirvnger, 1956). В. Беспозвоночное: бабочка-листовидка (Kallima) со сложенными крыльями, так что видна нижняя поверхность обоих крыльев (Cott, 1951). Г. Беспозвоночное: листовидка Chitonisctis feedjeanus; видоизменения передних крыльев, в том числе средней и боковых жилок, делающие ее похожей на лист (Cott, 1951). По Лима-де-Фариа.
Однако Лима-де-Фариа заявляет о себе как о крайнем редукционисте, Он серьезно полагает, что «биологическая эволюция полностью обусловлена упорядоченностью трех предшествовавших эволюций» и что все биологические явления можно и нужно свести к законам физики и химии (там же, с. 22, 365). Закономерности и канализация органической эволюции задаются уже на уровне элементарных частиц, а последующие уровни их только еще больше упорядочивают.
Следствием такого жестко канализованного развития оказывается, по Лима-де-Фариа, гомологичность изоморфизмов, наблюдаемых в живой и неживой природе. На страницах его книги мы видим многочисленные и зачастую поразительные примеры морфологического сходства кристаллических структур минералов со структурами животных и растений (рис. 14–18). Но при этом мы не обнаруживаем у него ни малейшего стремления к отысканию какого-либо критерия для отделения сходств, имеющих общую причину, от чисто внешних, ничего не говорящих аналогий. В результате в одном ряду с примерами, заслуживающими изучения, помещены и подобные следующему: сеть жилок листа Arum, капиллярная сеть лапки лягушки и фрагмент растрескавшегося от засухи песчаника (!). Но самое слабое место развиваемой концепции состоит в том, что ее автор подбирает только желательные примеры, но не может доказать единства лежащего в их основе механизма, ибо, как он сам признает, физика элементарных частиц и особенно процесса кристаллизации еще остается недостаточно изученной.
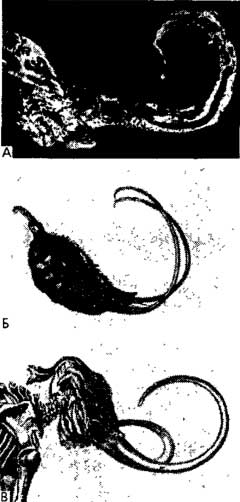
Рис. 15. Изогнутые выросты. А. Минерал, самородки серебра обычно имеют изогнутую форму (Ehrhardt, 1939). Б. Растение: плод Martynia lutea (сем. Martyniaceae) (Heywood, 1978). В. Позвоночное: скелет мамонта, вымершего хоботного, обитавшего не севере Европы и е Азии (Pierantoni, 1944). По Лима-де-Фариа.
Лима-де-Фариа убежден, что причину биологической эволюции надо искать в том, откуда берут свое начало форма и функция. А они, безусловно, имеют своих предшественников в мире минералов, химических элементов и элементарных частиц. В этом мире еще не было и в помине ДНК и генов, а закрепление во времени характерных структур (паттернов) уже имело место. Так проявлялось свойство атомов, молекул и минералов. «Спиральная форма, характерная для раковин моллюсков, — пишет Лима-де-Фариа, — уже существовала в галактиках, а гексагональные структуры глаза насекомого — в снежинках» (там же, с. 330).
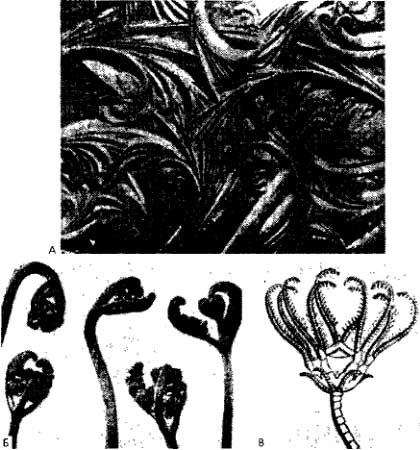
Рис. 16. Растущие кристаллы и органы. А. Молекулы: кристаллы льда, образовавшиеся из конденсирующихся паров воды (Cabrera, 1937). Б. Растение: молодые побеги папоротника Pteridium aquilinum (Cabrera, 1936). В. Беспозвоночные: личиночная стадия стеблевой морской лилии (Pierantoni, 1944) По Лима-де-Фариа.
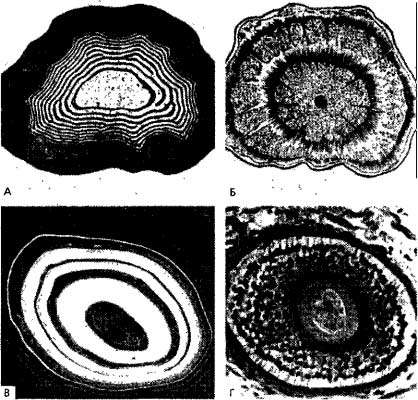
Рис. 17. Слоистые кольцевые структуры. А. Молекулярный процесс: кольца Лизеганга в смеси азотнокислого серебра и двухромовокислого калия в желатине (Rinne, 1928). Б. Растение: поперечный разрез стебля Mucuna altissima (сем. Papilionaceae) (Strasburger, 1943). 8. Минерал: агат; растворенные минералы осаждаются слоями в тонкозернистом кварце (Desautets, 1968). Г. Позвоночное: поперечный разрез основания волоса в кожечеловека (Nachtigall, Kage, 1980). По Лима-де-Фариа.
Становится понятно, почему Лима-де-Фариа назвал свою концепцию автоэволюцией. Действительно, развертывание биологических программ, согласно этой концепции уже предусмотрено процессом трансформации, заложенным в первичной материи и энергии. Одним из наглядных проявлений высокой степени независимости развития от внешних причин выступают явления самосборки и самоорганизации, проявляющиеся на всех уровнях — от первозданной материи до человеческих сообществ. К спонтанной упорядоченной ассоциации способны как элементарные частицы, атомы и молекулы, так и клетки, органы, организмы и сообщества.
Очевидно, что с концепцией автоэволюции совершенно несовместимы случайность и отбор, и Лима-де-Фариа возражает против них особенно резко, призывая на помощь все свое красноречие. Он называет отбор «абстрактной концепцией», которая должна быть изгнана из биологии. Что касается случайности, то это тоже вымышленная категория, которую постоянно эксплуатируют «неодарвинисты» для прикрытия собственного невежества. В биологических явлениях нет места случайности, и даже мутационный процесс благодаря молекулярным ограничениям носит направленный характер.

Рис. 18. Структуры, напоминающие цветок. А. Минерал: арагонит — карбонат кальция с большей плотностью, нем кальцит (Ehrhardt, 1939). Б. Беспозвоночное: актиния с ее «жалящими» щупальцами (Wheeler, 1940). В. Растение: продольный разрез цветка Cycadeoidea ingens (Strasburger, 1943). Г. Беспозвоночное: голотурия Cucumaria plane (Pierantoni, 1944). По Лима-де-Фариа.
В представлении Лима-де-Фариа законы и механизмы эволюции еще предстоит открыть, а гены и хромосомы играют в ней лишь второстепенную роль.
Между автоэволюцией и преформизмом можно вполне поставить знак равенства. Лима-де-Фариа считает, что даже самые крупные макроэволюционные события, ознаменовавшие становление планов организации, не означают появления чего-то истинно нового. Уровень эволюции, воспринимаемый как новый, возникает в результате перекомбинирования уже существовавших компонентов. И так во всем, будь то какая-то особая клетка, форма, структура или функция.
Одним из первых, кто воспринял в Советском Союзе концепцию Берга после, казалось, ее полного забвения, был известный палеонтолог Б. Л. Личков (1965), горячий сторонник Ж. Кювье и автор одной из сопряженных с геологическими циклами гипотезы эволюции. По его убеждению, все прогрессивное развитие органического мира совершалось не на основе случайностей, а в силу строгих закономерностей. Однако уточнять эти закономерности, а тем более раскрывать их природу Личков не стал.
Эту миссию взяли на себя талантливый палеоботаник и теоретик-эволюционист, рано ушедший из жизни, С. В. Мейен и тесно сотрудничавший с ним Ю. В. Чайковский. Можно сказать, что они стали главными продолжателями Берга и некоторым образом Любищева в силу своей естественной склонности искать закон и порядок там, где его особенно трудно обнаружить. Мейен и Чайковский вообще стремились к «номотетизации» биологии, но, в отличие от господствовавшей тенденции к осуществлению этого намерения с помощью редукционистской методологии, они решали эту задачу путем познания биологического разнообразия.
Как известно, порядок в разнообразии, изучаемом типологией (у Чайковского — диатропикой), создает систематика (таксономия), которая опирается на данные о строении организмов, или, точнее, об их признаках. Эту науку о признаках Мейен (1977) предложил называть мерономией. В отличие от Таксономии, распределяющий формы организмов по группам, в случае мерономии мы делим организм на части — по морфологическим, физиологическим или экологическим признакам, а классифицируя эти последние, получаем мероны («классы частей»). Примерами меронов могут служить любые части целого (органы, ткани, определенный тип клетки, физиологическая функция и т. п.), общие для данного таксона. В сумме они составляют его архетип, или план строения. Мерономия обеспечивает таксономию «признаковым пространством и данными о соотношении признаков у разных объектов» (Мейен, 1978. С. 496).
Теперь, сравнивая ряды параллельных таксонов, Мейен констатировал, что у них наблюдается сходный, а иногда и идентичный набор меронов. Чем ближе друг другу таксоны, тем больше число одинаковых меронов. У видов одного рода они почти все совладают. Эту повторяющуюся последовательность меронов в паралельных таксонах Мейен назвал рефреном (там же, с. 501).
По аналогии с периодическим законом в химии он считал полезным графическое отображение рефренов в виде таблицы, где по горизонтали представлены изменения мерона в рядах сравниваемых таксонов. Вертикальные столбцы означают тогда одинаковые состояния выбранного мерона в этих таксонах (аналог одинаковой валентности в таблице Менделеева). Фактически это те же гомологические ряды Вавилова, только сведенные в таблицу. Мейен считал, что подобная форма записи ценна не столько для систематики, сколько для понимания процесса эволюции.
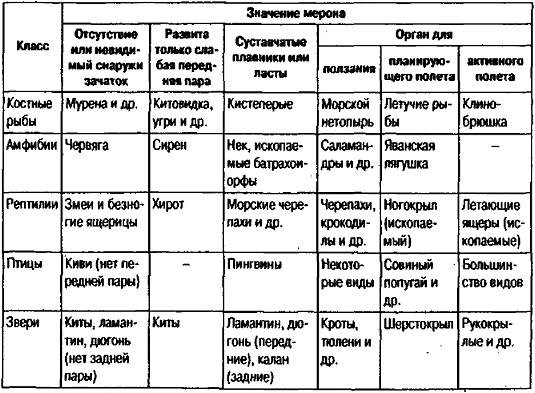
Рис. 19. Рефрен мерона «парные конечности» (из: Чайковский, 1990).
Так, при подобной записи мерона «парные конечности» (рис. 19) для всех классов позвоночных выявляется общий рефрен: от полного отсутствия обеих пар до образования органа полета — крыла. Таблица наглядно демонстрирует возникающее в силу закона параллелизмов признаковое пространство, позволяющее предсказывать, что может и чего не может быть в эволюции. В природе, правда, вовсе не обязательно должно существовать такое число разных форм, какое способно занять все клетки таблицы. Так, возвращаясь к мерону «парные конечности», надо отметить, что в истории никогда не было крылатых амфибий и птиц без задних конечностей. Мейен справедливо замечает, что без выявления всего мыслимого разнообразия нельзя установить и существования многих запретов.

Рис. 20. Типы расчленений листовой пластинки (из: Мейен, 1973).
Один из самых наглядных примеров рефрена на ботаническом материале — типы расчленения листовой пластинки (рис. 20). Хотя в типологии обычен поиск закономерности в повторяемости признаков внутри таксонов, самая суть рефренов, по мнению Мейена, в выявлении закономерностей в изменчивости признаков между таксонами. Данные о них фрагментарны. Но без знания рефренов заполнить по отдельности все разновидности переходов между меронами — столь же невыполнимая задача, как заполнить все склонения каждого существительного, если не знать правил склонения.
Нам представляется весьма важной также еще одна закономерность формообразования в эволюции, выделенная Мейеном и названная им транзитивным полиморфизмом (Мейен, 1978). Она состоит в том, что новый таксон рождается с тем же набором составляющих его таксономических единиц или форм, существовавших у предкового таксона, которые повторяют и соответствующие признаки (рефрен). Причем даже небольшое число уцелевших особей способны восстановить все внутренее разнообразие истребленного таксона. Иными словами, разнообразие порождает разнообразие, на каком бы уровне мы его ни рассматривали, и только что сказанное — его следствие. Существование подобной закономерности в сочетании с происходящей при смене поколений свободной комбинаторикой признаков чрезвычайно усложняет обнаружение таксонов-предков и восстановление филогении, а то и вовсе делает это занятие бессмысленным. Эволюция идет и сразу «заметает» за собой следы.
Надо полагать, что, исследуя эту закономерность, Мейен знал, что идея об эволюционном переходе внутривидовой структуры от вида к виду уже существовала (Алтухов, Рычков, 1972). Мейен только расширил ее рамки.
Ученый обратил внимание на то, что в формулировке закона Вавилова речь идет только о повторности признаков от таксона к таксону, но в ней ничего не говорится о повторности в правилах их преобразования. С точки же зрения выявления типологических закономерностей это имело бы особое значение, Гораздо важнее самих морфологических параллелизмов тот факт, что, например, в разных семействах цветковых наблюдаются одинаковые тенденции в преобразовании морфологических особенностей. Именно общность тенденции, наличие одного правила преобразования позволяют предсказывать и целенаправленно искать недостающие члены параллельных рядов.

Сергей Викторович Мейен (1935–1987).
Мейен не ограничился только упорядочением закономерностей в рядах изменчивости, установленных своими предшественниками; он идет дальше, расширяя рамки самого закона Вавилова. Он убедился, что полнота проявлений параллелизма не всегда связана с генетической общностью. Сплошь и рядом она, напротив, обратно пропорциональна систематической близости таксонов. Так, параллелизм между головоногими моллюсками и фораминиферами по спиралям раковины гораздо полнее, чем между головоногими и брюхоногими по тому же признаку; параллелизм жизненных форм кактусов и молочаев полнее, чем между кактусами и более близкими к ним Caryophyllaceae. Немало примеров параллелизма в строении цветков и соцветий, листьев и филлодиев, в биохимических и генетических характеристиках далеких форм. Еще более удивительны параллелизмы между живыми и неживыми объектами, где уже не приходится говорить о конвергенции ввиду сходных условий существования. Примеры таких параллелизмов собрал еще д’Арси Томпсон (1942), а ближе к нашему времени — Ю. А. Урманцев (1970, 1988) и А. Лима-де-Фариа (1991). Урманцев, в частности, подтвердил сходства гомологических рядов в развитии животных и растений с таковыми спиртов и углеводородов, установленные, соответственно, Э. Копом и Н. И. Вавиловым; обнаружил сходство между 9 изомерами венчика барбариса и 9 изомерами инозита, сходство генома с языком (речью), эволюционной генетики со сравнительным языкознанием и многие другие. Объяснение существованию подобного рода параллелизмов Мейен, вслед за Урманцевым, усматривает в системной природе объектов (Мейен, 1975).
Названные случаи изоморфического сходства, не сводимые ни к генетической общности, ни к подобию условий существования, привели Мейена к выводу о существовании чисто морфологических (типологических) и нестатистических законов, являющихся наиболее общими и пока еще очень слабо изученными. В свете такого взгляда закон гомологических рядов Вавилова, очевидно, становится его частным проявлением.
Следует отметить еще один новый момент в трактовке все того же закона. Обычно не учитывается, замечает Мейен, что признаки радикала тоже иногда испытывают изменчивость, которая в силу своей редкости считается тератологической. Эта изменчивость тоже следует закону Вавилова, причем часто признак, расцениваемый как уродство в одном таксоне, становится нормой в другом. Этот частный случай, описанный Н. П. Кренке (1933-1935), Мейен предложил называть правилом Кренке.
Среди ученых — экспериментаторов и теоретиков — безусловно существуют в относительно чистом виде две категории: «примирителе», склонные к компромиссу и объединению кажущихся противоположностей в высшем синтезе, и «непримиримые», производящие выбор среди противоположностей и стремящиеся подавить бракуемую. Любищев принадлежал ко второй, а Мейен — к первой. Но на путях к синтезу Мейена постигла явная неудача.
Развивая стержневую идею номогенеза, он, в отличие от Берга, не создал целостной концепции. Берга интересовали и закономерности эволюции, и ее движущие силы. Собственно, с постулата, что автономический ортогенез — главная побудительная сила всякого прогрессивного развития, и начинается изложение его концепции о направленности эволюции. Во времена оные за это Берга наградили нелестными эпитетами — считали идеалистом, автогенетиком, преформистом, виталистом… Можно ли было в конце XX в. как-то развить, усовершенствовать это ядро номогенеза? Это нелегко. Для этого нужно предложить свое видение проблемы, так как в первую очередь всех интересуют именно факторы эволюции.
Мейен принял в качестве факторов эволюции мутации и отбор в том виде, как они еще существуют в СТЭ. Более того, он считал своей главной стратегической задачей поиски путей снятия противоречий между тихогенезом (селекционизмом) и номогенезом (Мейен, 1974, 1978, 1984а) и полагал, будто она достигнута с созданием общей теории систем, в рамках которой дивергенция и параллелизм стали дополнительными понятиями.
А между тем фундаментальное и в наши дни исключительно актуальное обобщение Мейена о тропиках как «колыбели» и «музее» растительного багатства планеты (Мейен, 19846, 1986) находится в разительном противоречии с теорией селектогенеза. На огромном ископаемом материале ему удалось показать, что почти все наиболее крупные таксоны, включая семейства, возникли в фитохориях, лежащих в экваториальном поясе. И это стало возможно, констатирует Мейен, только благодаря тому, что естественный отбор здесь сильно «заторможен» и потому допускает всевозможные эксперименты природы. .
Можно только гадать, как такой высокоэрудированный специалист с острым и проницательным умом мог закрывать глаза на очевидную несовместимость СТЭ и номогенеза.
Другой убежденный последователь номогенеза — Ю. В. Чайковский, принимая и высоко оценивая вклад Мейена, избежал его непоследовательности. Обладая строго системным мышлением комби нативного типа, он не мог не воспринять экосистем ной теории эволюции, новой генетики и новых принципов индивидуального развития. Именно его труды лучше всего показывают, что старая эволюционная парадигма заслуживает не улучшения, а замены.
К познанию законов и механизмов эволюции Ю. В. Чайковский подходит как методолог и системолог (самому ему больше импонирует считать себя натурфилософом!), стремящийся вовлечь в познавательный процесс весь исторический багаж биологических знаний и достижения всех остальных естественных и многих гуманитарных наук (вплоть до лингвистики и мифологии), способных послужить нуждам эволюционной теории. Отсюда глубокое убеждение Чайковского, что данная теория может развиваться и дальше только как междисциплинарная отрасль знания (Чайковский, 1994). Значение этого методологического принципа он убедительно продемонстрировал в своей монографии об эволюционной диатропике (Чайковский, 1990).
Одной из самых продуктивных для эволюционной теории дисциплин Чайковский считает современную термодинамику неравновесных процессов с ее ключевой идеей самоорганизации и самосборки. Как известно, она была разработана в трудах И. Пригожина с соавторами (Николис, Пригожин, 1979; Пригожин, Стенгерс, 1986) и Е. Янча (Jantsch, 1980). По его мнению, именно данный раздел термодинамики демонстрирует неизбежность саморазвития таких сложных и далеких от равновесия систем, какими являются живые организмы. Термодинамика побудила к отказу от взгляда на эволюцию как последовательную цепь реакций биологических объектов на внешние воздействия и представила ее как совокупность актов самоорганизации. Междисциплинарный характер строящейся теории самоорганизации делает, по мнению Чайковского, излишним создание обособленной теории происхождения видов.
Чайковский совершенно справедливо считает логически неуязвимым современный взгляд на эволюцию как процесс преобразования систем и, следовательно, целостностей. Вслед за В. И. Вернадским, Дж. Берналом и Г. А. Заварзиным (1979, 1984) он полагает, что первичная жизнь на Земле возникла в форме экосистем, а не в виде отдельных и немногочисленных протоорганизмов, и потому единственно адекватным отображением ее первых эволюционных шагов может быть только системное описание. «Эта жизнь умеет существовать только целиком, и мы не в силах всерьез представить себе, как мог бы существовать один-единственный вид, а не то что единственная особь» (Чайковский, 1990. С. 183).
Фактор целостности выступает как направляющая сила, он един и для индивидуального развития, и для филогенеза. Некоторые философы (Мещерякова, 2001) склонны допустить идеальную природу целостности и тем воскрешают в памяти небезызвестную концепцию холизма первой половины XX в.
Наконец, как номогенетик Чайковский видит в учении о биологической эволюции часть или раздел современного глобального эволюционизма и мыслит его дальнейший прогресс в рамках их тесного взаимодействия.
Выше уже упоминалось о загадочных топологических законах развития параллелизмов, не сводимых к известным причинам. В 1970-е годы именно этот аспект существования и преобразования всего живого привлек пристальное внимание философа-системолога Ю. А. Урманцева. Изучение поли- и изоморфизмов, описанных Бергом, Копом, Медниковым и другими, а также подмеченных им лично на большом числе объектов живой природы, послужило для Урманцева отправным моментом к разработке оригинального универсального варианта общей теории систем (ОТС) (Урманцев, 1971, 1974). Эта теория должна была прежде всего дать объяснение тем случаям изоморфического сходства, которые не могут быть сведены к традиционным общепринятым причинам — родству, одинаковым условиям существования, отправлению одинаковых функций. В конечном счете ОТС была призвана дать в руки исследователей четкое указание: «что должно быть», «что может быть» и «чего быть не может» в существовании и развитии любых объектов-систем, как материальных, так и идеальных (Урманцев, 1974. С. 51). Она также должна была обладать предельно широкими возможностями в части обобщений, предсказаний, постановки новых вопросов, связей с научными теориями и принципами.
В свете ОТС интересующее нас тонкое изоморфическое сходство есть следствие системной природы самих объектов-систем. Названное Урманцевым (1988. С. 64) системной общностью, оно являет собой, по его мнению, третий основный тип подобия, не сводимого ни к одному из типов сходства, известных в естествознании.
Не вдаваясь в технические тонкости, отметим, что, согласно Урманцеву (19886, 1999), существует математический закон системных преобразований, по которому любой объект-система (или их совокупность) может переходить в другой только посредством 7 неэволюционных преобразований (количества, качества, отношений и их комбинаций) и 7 эволюционных преобразований — всего 255 способами, а в пределе — бесконечным числом способов (при неоднократном использовании их). С точки зрения этого закона все созданные до этого момента доктрины и теории (диалектика, СТЭ, номогенез, морфогенез, эволюция «эволюции» и пр.) обладают существенной «неполнотой», и их необходимо достроить «на 6/8 и 7/8» (Урманцев, 1999. С. 50). ОТС побудила Урманцева к разработке в качестве ее раздела «общей теории развития систем природы, общества, мышления» — эволюционики (19886). Наряду с уже приведенными законами системных преобразований в нем рассмотрены также изменения в самой биологической эволюции.
Позволяя выявлять сотни и тысячи новых классов изоморфизма, ОТС и эволюционика хранят, однако, полное молчание по поводу запретов, неуклонно нарастающих при переходе от нижележащих уровней организации биологических объектов к вышележащим. Совершенно очевидно, что эти запреты «работают» против полиморфизации и тем самым ограничивают полноту параллелизмов.
Если ОТС предполагает возможность прогноза и способна предсказать, «что должно… и может быть», то биологи хотели бы, скажем, знать, почему до сих пор на Земле не было крылатых амфибий и безногих птиц.
Нам думается, что, создавая ОТС и увлекшись возможностями, которые она открывает для упорядочения многообразия материальных и идеальных объектов, Урманцев в неменьшей степени, чем Лима-де-Фариа, проигнорировал специфику живого, его способность порождать единичное, уникальное и неповторимое. Поэтому мы склонны полагать, что в деле постижения номогенетических аспектов органической эволюции объем задач, стоящих перед «чистыми» биологами, вовсе не стал меньше.
* * *
Поскольку стержневой идеей номогенеза является закономерный и направленный характер изменчивости и эволюционного процесса, это течение имеет много общего с финализмом. С последним его сближают также убежденность в единстве законов индивидуального и исторического развития, допущение преформированности филогенеза и, как правило, решительное неприятие концепции естественного отбора.
Стержневая идея номогенеза роднит его и с неоламаркизмом, а представления о скачкообразности формообразования и преобладании конвергентного пути эволюции создают для него точки соприкосновения с симгенезом и сальтационизмом.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК