Пермь
298,9 – 251,902 миллиона лет назад: Зима палеозоя, взлет и падение звероящеров
МЕЖДУНАРОДНАЯ И РОССИЙСКАЯ ШКАЛА
298,9 млн л. н.: приуральская эпоха: ассельский век (293,52) – сакмарский век (290,1) – артинский век (283,5) – кунгурский век или уфимский век (272,95) – гваделупская (биармийская) эпоха: роудский (казанский) век (268,8) – вордский (уржумский) век (265,1) – (татарская эпоха) кептенский (северодвинский) век (259,1) – лопинская эпоха: вучапинский (вятский) век (254,14) – чансинский век (251,902)

Пермский период – самый холодный период палеозоя и один из самых холодных за всю историю планеты. Ужасный климат не мог не подтолкнуть эволюцию – неспроста именно в перми появились все предпосылки для возникновения млекопитающих. Еще немного – и звери бы окончательно сформировались и заняли планету, а разум мог воссиять на сотни миллионов лет раньше. Но пермь – и самый трагический период: в его конце почти все достижения палеозоя оказались уничтожены, причем, как часто бывает с самыми ужасными событиями, этот катаклизм к тому же до сих пор малоизвестен большинству людей.
* * *
История открытия пермского периода весьма поучительна. Описал его английский геолог Р.И. Мурчисон, который до этого успел выделить силур и девон. Изыскания Р.И. Мурчисона в немалой степени касались залежей угля – главного промышленного источника энергии XIX века. Именно слава специалиста по углю позволила Р.И. Мурчисону оказаться в России, где важность его миссии была подчеркнута двумя встречами с императором Николаем I. Разгоняющаяся уральская промышленность требовала энергетической подпитки, а для эффективного использования несметных богатств требовался научный подход. За пять месяцев путешествий по России Р.И. Мурчисон исследовал огромное количество отложений. В некоторый момент его путь лежал в Вологду, но распутица не пустила именитого геолога в этот город, заставив повернуть в Пермь, о чем он вовсе не пожалел. А ведь в Вологодской области отложения пермского периода не менее выразительны, чем в Пермском крае. Если бы не дорожная грязь, один из важнейших периодов истории планеты назывался бы вологодским, или вологодием.
* * *
В перми все континенты в последний раз плотно прижались друг к другу в единую Пангею, как будто пытаясь согреться. На самом деле, причина и следствие тут прямо обратны: огромный суперконтинент перегородил экваториальные течения, что привело к похолоданию в мировом масштабе. Огромные размеры суши привели к континентальности климата: облака просто не долетали до глубинных областей, в которых раскинулись обширные пустоши и даже настоящие пустыни. Конечно, в экваториальной области холодов не было, а побережья и склоны гор исправно орошались дождями, но в среднем по планете было холодно и сухо.
Еще в конце гжельского века карбона начался раннепермский гляциопериод, продолжавшийся до начала артинского века. За это время случилось как минимум два ледниковых эпизода, следы которых обнаружены в Австралии, Бразилии, Африке, Антарктиде, Аравии, Индии, Бирме и на Малакке. Ледники расходились от Антарктиды во все стороны – вплоть до 30° южной широты, то есть до широты современных южных частей Африки, Австралии и Бразилии. Температуры были в среднем на 4° ниже современных – казалось бы, совсем ничего, но пермские оледенения уступают, и то ненамного, только самым страшным холодам криогения и плейстоцена.
Огромные массы пресной воды уходили в ледники, отчего соленость воды в океане резко повысилась – наибольшим образом за весь фанерозой. Соль концентрировалась, выпаривалась на побережьях и откладывалась огромными толщами. За весь пермский период ушло в осадок до 10 % соли от количества в современном мировом океане! Это значит, что до перми соленость океана могла быть на 10 % выше. Очевидно, это не могло не сказаться на всей океанической жизни.
Маленькая тонкость
Пермское соленакопление повлияло на мировую историю. Благодаря пермским оледенениям в Пермской области образовались огромные отложения солей, которые издавна разрабатывались местными жителями. Соль была одной из экономических основ про тогосударства Перми Великой, добывают ее и поныне. Неспроста возникло и словосочетание «пермяк – соленые уши», в прошлом сугубо ругательное, а сейчас ставшее «модным слоганом».
С другой стороны, из-за ухода воды в ледники уровень океана понизился, открыв огромные просторы, прежде залитые мелководными морями. На освободившихся просторах началось выветривание карбоновых угленосных отложений с сульфидами, пошло сульфатонакопление и, соответственно, захоронение кислорода. Однако продуктивность экосистем снизилась, в отличие от карбонового изобилия, органического вещества захоранивалось мало, ранее образовавшиеся карбонаты постепенно разрушались, отчего росла концентрация фосфатов и углекислого газа в воде, что, в свою очередь, стимулировало фотосинтез цианобактерий и водорослей и увеличивало количество кислорода. Такова экологическая диалектика!
Все же в целом жить становилось сложнее. Всю пермь разнообразие морских животных только потихоньку падало, превратившись под конец в ужасный обрыв в пропасть. Но до катаклизма были долгих 47 миллионов лет.
* * *
Показательны изменения кораллов: весь период их разнообразие и размеры только сокращаются. Табуляты стали мелкими и часто паразитическими, отличным примером чего могут служить Pseudofavosites. В конце периода коралловые постройки становятся редкими и локализованными в самых благоприятных местах.
Из кунгурского века Южной Африки известен первый достоверный ланцетник Palaeobranchiostoma hamatotergum довольно странного вида: в отличие от современного потомка, палеозойский имел почти треугольную форму за счет острого носа, крупного спинного и особенно длинного нижнего плавника. Самая удивительная особенность палеобранхиостомы – ряд колючек вдоль спины от «головы» до кончика спинного плавника. Как же сурова была пермь, если даже ланцетники становились ежиками!
Из рыб в перми особенно удались акулы. Едва ли не самые загадочные хрящевые всех времен и народов – Helicoprionidae. Первыми были умеренные представители – раннепермские Parahelicoprion с Урала и из Боливии, а также позднепермский Sarcoprion edax из Гренландии. Зубы на передней части их нижней челюсти – симфизе – выступали выпуклой вверх дугой. Как часто бывает с хрящевыми рыбами, общий вид и даже размеры тела устанавливаются сугубо предположительно. Некоторые парагеликоприоны, вероятно, достигали аж 12 м длины, саркоприон был вдвое скромнее – «всего» 6 м. Таким образом, эти рыбы имеют рекордные для палеозойских животных размеры.
Экзотическим развитием таких монстров стали ранне-среднепермские представители Helicoprion, из которых первым был описан и больше всего известен H. bessonovi. У этой рыбы зубы заворачивались довольно плотной спиралью на два-три полных оборота, причем в центре спирали зубы были маленькими, а к наружному концу увеличивались; в одной спирали могло быть до 190 зубов. Судя по всяческим тонкостям, рост начинался именно от центра спирали, правда тогда трудно понять, как вся конструкция крепилась – неужели челюсть заворачивалась как рулон или вовсе была некая хрящевая ось? Стертость зубов минимальна, так что вообще малопонятно – зачем тогда такое количество зубов, если они почти не использовались? Согласно наиболее современным и проработанным реконструкциям, зубная спираль геликоприона лежала на дне ротовой полости или даже в глотке и использовалась для обрезания щупалец головоногих моллюсков. Учитывая строение родственных акул, законно предположить, что во рту геликоприона могли быть и зубы других типов, использовавшиеся для раздавливания панцирей и раковин. Самые огромные спирали – до 56 см – могли принадлежать рыбам 7,5 м, а по другим оценкам – 9 – 12 м длиной.
Работа над ошибками
Чего только не предполагали палеоихтиологи относительно местоположения и способа использования зубных спиралей: что они загибались вверх на носу, вниз, вперед или вверх на симфизе нижней челюсти, в глотке, в специальной капсуле на дне ротовой полости, на спинном или даже хвостовом плавнике, что у одной рыбы была одна спираль или пара, что вся конструкция могла двигаться-распрямляться, что старые зубы были самыми большими или самыми маленькими. Пока кроме самих спиралей не найдено никаких других остатков геликоприонов, все это – более или менее вероятные предположения. Впрочем, сравнивая геликоприонов с другими родственными акулами, можно смело исключить половину вариантов. Рано или поздно найдется красивый отпечаток головы, и одни палеоихтиологи будут торжествовать: «Я же говорил!», а другие скромненько помалкивать.
Другие необычные пермские акулы – Xenacanthida (или Xenacanthiformes), например, Orthacanthus и Xenacanthus. Это были немаленькие рыбки – от метра до трех и даже четырех, особенно учитывая, что жили они в пресных водоемах. У этих вытянутых акул на загривке торчал здоровенный шип, который у предков располагался в основании спинного плавника, но со временем сместился на голову; спинной же плавник превратился в длинную низкую лопасть. Верхняя лопасть хвостового плавника стала длинным заостряющимся концом тела, а нижняя лопасть сместилась далеко вперед, став фактически вторым анальным плавником. Зубы имели странную V-образную форму, видимо, предназначенную для разгрызания панцирей раков и палеонисков. На самом деле, представители обоих упомянутых родов появились еще в конце девона, а вымерли аж в конце триаса – удивительная стабильность, учитывая массу случившихся за это время событий.

Xenacanthus
В начале перми (но, что характерно, не на границе периодов) вымирают остатние акантоды и кистеперые рипидистии. В пермских отложениях найдена ископаемая нора двоякодышащего чешуйчатника Gnathorhiza: конкреция в форме не то булавы с желобком, не то очень толстой ложки, образовавшаяся, когда рыба закопалась в глубокий ил, а потом свернулась там пополам для сна во время засухи, да только сон оказался вечным.
В конце перми от палеонисков отделяются костные ганоиды Holostei, уже очень похожие на полноценных костистых рыб и иногда объединяемые с ними в одну группу Neopterygii. Древнейшая амия Acenthrophorus varians из поздней перми Англии выглядела довольно скучно, как и полагается великому предку огромной группы, возникшей из нее.
Работа над ошибками
В научной терминологии иногда допускаются большие вольности, а иногда несколько букв полностью меняют смысл. Одна из самых распространенных путаниц – с понятиями «костные» и «костистые» рыбы. Костные рыбы Osteichthyes – это все, у которых хоть где-то в скелете есть костная ткань; при этом большая его часть запросто может быть хрящевой, как у хрящевых ганоидов и лопастеперых. Костистые же рыбы Teleostei – это только совсем-совсем костные, у которых окостеневает весь скелет. Ясно, что есть и масса других важнейших признаков. Костные ганоиды попадают между первыми и вторыми: их скелет окостеневает, а потому иногда они объединяются с тру-костистыми в одну группу Neopterygii, но их чешуя ганоидная, а в строении есть масса архаики. Современные костные ганоиды включают только панцирных щук Lepisosteus и амию Amia calva, но в мезозое это была очень богатая группа.
Панцирные щуки неспроста иначе называются каймановыми рыбами – их зубастая морда, правда, напоминает крокодилью. Любимое дело американских рыбаков – поймать парочку панцирников, положить на берег, отойти метра на два и сфотографироваться в пафосной позе с большой глубиной резкости так, чтобы казалось, что отважный рыболов отловил каких-то ужасающих монстров. Впрочем, некоторые панцирники и безо всякого монтажа вымахивают до четырех метров длины. Маленьких же щучек иногда держат в аквариумах, где они радуют глаз своим палеозойским обликом.
* * *
Сухопутные флоры и фауны перми были удивительно однообразны от Полярного Урала до Южной Африки, ведь единство Пангеи гарантировало легкость миграций.
Маленькая тонкость
Между прочим, именно распространение идентичных следов карбоновых и пермских оледенений на удаленных ныне материках, а также сходство фаун Гондваны в перми и триасе натолкнули А. Вегенера на мысль о дрейфе континентов.
Впрочем, в разных областях экосистемы все же выглядели не идентично. В последних заповедниках влажных тропиков Еврамерийской области древовидные хвощи и плауны мельчали. Поздние Calamites и Sigillaria, а также менее известные Paichoia и Signacularia гораздо скромнее своих величественных предков. Болота, из которых торчали столбы плаунов, покрытые сплавинами – плавающими матами – из водяных мхов, печеночников, водорослей и цианобактерий, постепенно сохли, а их места занимали папоротниковые и хвойные леса. Крайне распространенными тут растениями были многочисленные виды древовидного папоротника Asterotheca.
Холодные северные области Приуралья и Сибири были покрыты бесконечной голосеменной кордаитовой тайгой. Сибирь – она и есть Сибирь, даже приближенность к экватору ей не помогала, тайга и холод – 300 миллионов лет стабильности! Пермские кордаиты имели совсем мелкую листву, примером чего могут служить многочисленные виды Crassinervia.
Холодные южные области, а таковыми по-прежнему были слившиеся Южная Америка, Африка, Индия, Антарктида и Австралия, заросли лесами из примитивных голосеменных Glossopteridales (все тот же Glossopteris) и Ginkgoopsida.
Первые достоверные гинкго – французские Trichopitys heteromorpha и Sphenobaiera – появились в начале перми. У них были узенькие дихотомически ветвящиеся листья, больше похожие на иголки, плотно усыпающие побеги. Другой вероятный родственник гинкго – аргентинский Polyspermophyllum sergii, чьи листья были похожи на змеиные языки; наверняка так бы и назвали его местные жители, доживи он до современности.
В конце перми как на севере, так и на юге появились новые леса, составленные елкоподобными Voltzia, с несколькими родственниками образующими группу Voltziales. Это были засухоустойчивые деревья, листья которых превратились в чешуеподобные иглы, плотно покрывавшие ветви; впрочем, кончики веток были украшены длинными иглами типа сосновых.
* * *
Смена флоры приводила к смене членистоногой фауны.
Впрочем, пермские ракообразные щитни Triops и Lepidurus на родовом уровне уже точно не отличались от современных; в принципе, их можно упоминать в каждом последующем периоде, причем одними и теми же словами, для них День Сурка настал давно и безнадежно.
Архаичные древнекрылые насекомые никуда не делись. Из карбона в пермь плавно перешли палеодиктиоптеры Goldenbergia и Dunbaria, питавшиеся пыльцой кордаитов. Отлично чувствовали себя поденки, например Misthodotes, причем известны и их личинки – Kukalova americana.
Как бы ни славились своими размерами карбоновые протострекозы, самым большим насекомым за всю историю планеты был раннепермский Meganeuropsis permiana из Канзаса и Оклахомы: размах его крыльев достигал 71 см! Другие немаленькие пермские меганевры – Tupus и Arctotypus.
В перми возник отряд веснянок Plecoptera – родственников тараканов, чьи личинки опять перешли к водному образу жизни. Древнейшая веснянка Perlopsis filicornis из кунгурского века Приуралья известна и во взрослом виде, и в личиночном; позже появилось великое множество ее родственников. Обилием растительной трухи питались также первые сеноеды Psocoptera (или Copeognatha; например, Permopsocus congener) – далекие и пока вегетарианские предки вшей.
В начале перми возникли предки цикад и клопов – например, Archescytina, Maueria и Permopsylla. Трудно назвать самую-самую первую цикаду, они появляются вдруг и сразу большой компанией. Само по себе существование сосущих растительные соки насекомых крайне показательно: стало быть, растения стали достаточно сочными, чтобы из них можно было высосать какую-то пользу.
Как вы там, потомки?
Два близких отряда насекомых – цикады Homoptera и клопы Hemiptera некоторыми энтомологами объединяются в один отряд хоботных Rhynchota, или Cimicida. Их объединяет сосущий ротовой аппарат, снабженный острыми стилетами для протыкания твердых предметов. Отличает же их то, что все цикады (включая, кстати, тлей) пьют сок растений, а их передние и задние крылья устроены примерно одинаково, тогда как многие клопы – хищники, а их передние крылья наполовину жесткие. Из различий крыльев проистекают альтернативные названия: цикады – равнокрылые, а клопы – полужесткокрылые.
Среди позднепермских цикад можно видеть предков современных листоблошек Psylloidea – Protopsyllidium, белокрылок Aleurodoidea – Permateurodes rotundatus, настоящих цикад Cicadina (или Auchenorrhyncha) – Prosbolopsis ovalis, а также других родственников. В средней перми Франции жила Lutevanaphis permiana – предок тлей Aphidomorpha (или Aphidoidea).
Сетчатокрылые Neuroptera (они же Planipennia, или Myrmeleontida) известны уже из ранней перми Чехии и США; много их найдено в местонахождении Чекарда в Пермском крае. В поздней перми они становятся очень многочисленны, хотя и однообразны, отличным примером может служить, скажем, Sialidopsis kargalensis. Родственный, полностью вымерший отряд Jurinida (или Glosselytrodea) был представлен, например, видами Archoglossopterum shoricum и Glossopterum martynovae.
Среди многочисленных отрядов насекомых с полным превращением самым важным, конечно, были жуки, или жесткокрылые Coleoptera (или Scarabaeida), достоверно появившиеся в начале перми, например, чешский Moravocoleus permianus. Показательно, что крайне трудно назвать самого древнего жука: в самых ранних фаунах они представлены уже значительным набором видов. Очевидно, особо удачная форма тела сразу дала столько преимуществ, что эволюция рванула с места в карьер. К концу перми жуков было уже великое множество – Permocupes, Stegocupes и многие-многие другие. Здорово, что даже самые древние представители выглядели совершенно современно. Надкрылья – уплотнившиеся передние крылья, защищающие тонкие задние крылышки и брюшко, более-менее компактная форма, грызущие челюсти, неспециализированные ножки – все эти черты не поменялись с пермского периода до наших дней. Такая конструкция тела оказалась универсальной, с минимальными изменениями она может быть приспособлена для любого образа жизни – на деревьях и в траве, в пещерах и в пустыне, под землей и под водой, где угодно. Неспроста жуки – отряд с самым большим числом видов среди всех живых организмов на планете! Если кто не знает, мы живем на планете жуков, а послепермская эволюция всех других животных лишь незначимая пенка по краям их величия.
В начале перми появились также скорпионницы Mecoptera (или Panorpida, например, Platychorista venosa), из которых быстро возникли ручейники Trichoptera (или Phryganeida, например, Microptysma sibiricum) – предки будущих бабочек.
Как вы там, потомки?
Глядя на современных потомков пермских насекомых, мы можем ощутить себя в позднем палеозое.
Муравьиные львы Myrmeleo – самые известные сетчатокрылые – радикально отличаются в детстве и взрослом виде. Личинка муравьиного льва – хтоническое чудовище в полном смысле этого слова, злобное существо, роющее коварные ямы-ловушки в песке, сидящее на их дне и сшибающее попавших туда муравьев песчинками, а потом цепляющее несчастную жертву огромными зазубренными челюстями и жадно выпивающее кровушку. Взрослый же муравьиный лев – невинный эльф, пьющий нектар или иногда вообще не способный питаться, порхающий на лужайках над цветочками. Многие видели муравьиных львов, но не все об этом догадываются, путая их со стрекозами. На самом деле, отличия очевидны: тогда как стрекоза – целеустремленный хищник с огромными глазами, виртуозно маневрирующий и стремительно настигающий жертву, муравьиный лев выглядит вроде бы тоже как стрекоза, но основательно перебравшая браги – с маленькими круглыми бессмысленными глазками, расхлябанно повисшими длинными усиками, бесцельно болтающийся над цветочками, бестолково, вразнобой и как попало размахивающий своими крылышками.
Глядя на взрослую златоглазку Chrysopa – изящное эфемерное, почти прозрачное создание с золотыми глазками, трудно представить, что ее личинка – ужасное колючее существо с огромными челюстями-клешнями, которое не только заживо высасывает тлей, но еще и делает из их шкурок накидку от солнца; да что уж «накидка» – часто это огромная безобразная куча разодранных кож на спине. Маньяки-каннибалы из фильмов ужасов – просто малые дети на фоне личинок златоглазок. Впрочем, изуверское детство сказывается в будущем: как бы ни были прекрасны взрослые златоглазки, стоит их тронуть – воняют они отвратительно.
Примерно то же можно сказать про похожих на бабочек франтов аскалафов Libelloides (или Ascalaphus), чьи личинки-гопники бросаются на ни в чем не повинных проходящих мимо насекомых из темной засады под камнем. У красивой взрослой Nemoptera, тоже подобной бабочке, задние крылышки превратились в изящные веслообразные хвостики, тогда как уродливая личинка похожа на помесь жирного паука с жирафом: у нее толстое тело, вытянутые тонкие ноги, ужасно длинная переднегрудь и огромные челюсти-клещи.
Отбросили всякую видимость приличия мантиспы Mantispa. Их личинки живут в коконах пауков, пожирая их яйца и вылупляющихся паучат в то время, как ничего не подозревающая паучиха старательно охраняет кладку. Некоторые мантиспы творят такой же беспредел в гнездах пчел и ос. Адски выглядит и взрослая мантиспа – с длинной переднегрудью и огромными клешнями, как у бог омола.
Здорово, что среди современных сетчатокрылых можно проследить все стадии приспособления от полностью почвенной личинки к полностью водной.
Скорпионницы Panorna замечательны тем, что у самцов брюшко имеет форму совершенно как у скорпиона. Более того, эти насекомые очень наглые и ничего не боятся. Если на вас села скорпионница, и вы решите потыркать ее пальцем, она не убежит, а начнет угрожающе замахиваться «хвостом» с «жалом». На самом деле, ничего вредного букашка сделать не может, но ее блеф всех весьма впечатляет, трогать ее правда совсем не хочется. Удивительно, но такое защитное поведение действует даже в районах, где самих скорпионов нет, так что боязнь ужаливания вроде бы совсем не актуальна. Вероятно, еще в палеозое, когда скорпионы были первыми и самыми опасными хищниками суши, у всех нововылазящих из воды быстро на генном уровне прописывался ужас от их образа. Строго говоря, никто не знает, как вообще можно в генах записать неприязнь к определенной форме тела, но, судя по успехам скорпионниц, как-то оно таки работает. Скорпионницы падальщики, причем едят трупы не только других насекомых, но и позвоночных животных.
Другие необычные родственники скорпионниц – комаровки Bittacus: они висят на растениях и ловят добычу своими очень длинными задними ногами, кончики которых работают как маленькие вывернутые задом наперед клешни богомолов.
Личинки ручейников знамениты тем, что строят домики-трубочки из всякого мусора, найденного на дне водоемов. На этом основан особый вид современного искусства: если осторожно вытащить личинку из ее домика и подкинуть в аквариум, скажем, бисера или золотого песка с брильянтами (у кого на что хватит), то личинка добросовестно выстроит ювелирный шедевр.
Как видно, в перми жило множество самых разных летающих насекомых, чьи личинки жили в воде – поденок, веснянок, ручейников и прочих. Как бы ни были субтильны эти создания, они умудрились устроить сначала революцию на суше, а после – грандиознейшую экологическую катастрофу по всей планете. Дело в том, что эти насекомые были единственными, кто переносил вещество из воды на сушу. До сих пор такого на планете не было. В лучшем случае какие-то бывшие участки морского дна поднимались над водами или из-за тектонических движений, или из-за опускания уровня океана. Но тут появилась новая сила, которая без остановки и в промышленных масштабах стала перекачивать азот и фосфор из водоемов на водоразделы: личинки накапливали в себе вещества, а взрослые насекомые разлетались в разные стороны. Казалось бы, одна букашка практически ничего не весит, но, как часто бывает, бесконечные числа отлично срабатывают – миллиарды насекомых каждый год на протяжении миллионов лет способны перетащить в своих хилых тельцах миллионы тонн ценнейших веществ. Например, на озере Мичиган сброшенные от последней линьки шкурки поденок могут покрывать берега едва ли не метровым слоем. В дочеловеческие же времена масштабы бедствия могли быть на порядки больше.
Ясно, что такое обильное удобрение в первом приближении создавало идеальные условия для роста наземных растений и помогало им в нелегком завоевании холодеющей суши. С другой же стороны, беспрецедентный вынос азота и фосфора из воды не мог не сказаться на водных экосистемах – и сказался. Но до катастрофы на суше успело произойти еще много интересного…
* * *
Хотя стегоцефалы во второй половине карбона серьезно пострадали, многие группы продолжали существовать и в перми.
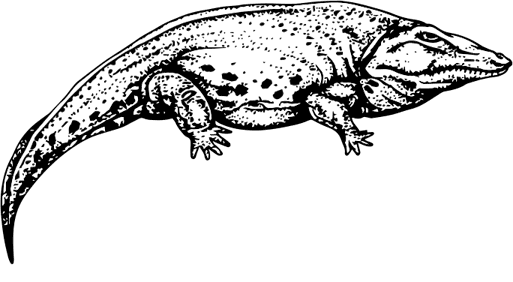
Eryops

Cacops aspidephorus
Самыми выдающимися амфибиями перми были, без сомнения, гигантские «живые капканы» Eryopoidea – двухметровый Eryops с шестидесятисантиметровой трапециевидной головой, Acheloma, Sclerocephalus, Cacops aspidephorus и многие другие их родственники. Эти огромные твари с короткими мускулистыми ногами, коротким широким телом с мощными ребрами, коротким хвостом и головой длиной с туловище лежали в донном иле и подстерегали добычу подобно тому, как это делают современные хищные пресноводные черепахи. Это были истинные крокодилы своего времени, и зубищи у них были соответственные. Сходство усиливалось рядами костных пластин на спине, иногда даже срастающихся с позвонками, что не очень-то типично для амфибий. Черепа родственных Platyoposaurus (он же Platyops), Archegosaurus и Melosaurus были еще больше похожи на крокодильи: длинное узкое рыло, острые зубы – мелкие по всей длине челюсти и большие на кончике, глаза, смотрящие вверх.
Крайне своеобразно выглядел ближайший родственник эриопоидов – Platyhystrix rugosus с самой границы карбона и перми США: на его спине возвышался высоченный гребень из остистых отростков позвонков, изогнутых и расширенных на концах, как клюшки, да еще к тому же с пупырышками. Вероятно, как и во многих случаях после – у пеликозавров и некоторых текодонтов и динозавров – такой гребень работал как радиатор: прохладным утром своей большой площадью он поглощал тепло и позволял холоднокровной зверюшке быстрее активироваться, а в жаркий полдень, стоило развернуться вдоль солнечных лучей, отдавал тепло вовне, чтобы не перегреться. С другой стороны, появление такой терморегулирующей системы однозначно говорит о сильных суточных перепадах температуры.
Продолжали радовать лепоспондилы нектридеи – полностью водная группа амфибий. Из них самые красивые, конечно, Diplocaulus и Diploceraspis, множество видов которых известно с границы карбона и перми США. Задние углы их черепов с возрастом несказанно разрастались вширь огромными рогами, размах которых у самого известного вида Diplocaulus magnicornis достигал трети метра. Как обычно, это был результат гонки средств нападения и защиты: проглотить такую жихарку-растопырку не могли даже самые широкомордые эриопоиды. Заодно на кончики рогов могли крепиться края кожных складок, тянущихся вдоль почти круглого плоского тела, так что амфибия представляла собой надувной матрас. Коротенькие ножки явно не были приспособлены для быстрого плавания и уж тем более бега. Видимо, такие существа медленно болтались вдоль дна, выедая там какую-то мелочь.
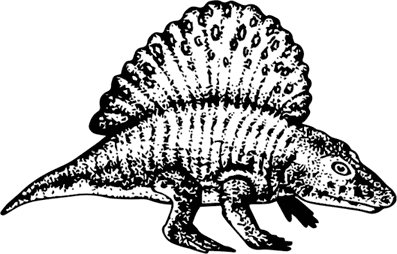
Platyhystrix rugosus
Уже в начале перми среди темноспондилов появляются амфибии, переходные от примитивных к современным. Идеальным великим предком нынешних хвостатых и бесхвостых амфибий является Gerobatrachus hottoni из кунгурского века Техаса. Внешне он был похож на саламандру с полукруглой лягушачьей головой. Другая раннепермская амфибия – Doleserpeton annectens из Оклахомы – тоже близка к основанию современных амфибий, но, что приятно, может быть основателем другой ветви амфибий – безногих червяг, хотя у самого долесерпетона с ногами все в порядке.
Несмотря на то что рептилии давным-давно появились, многие группы амфибий упорно продолжали рептилизоваться. Особенно знамениты в этом отношении Seymouriamorpha, а среди них – Seymouria baylorensis, вошедшая в учебники как переходное звено между амфибиями и рептилиями. Действительно, признаки сеймурий вполне промежуточны, проблема только одна – датировка. Даже раннепермские сеймурии опоздали на целый период. Как часто бывает, этот их минус был выявлен давным-давно, но учебники, вопреки стереотипу, переписываются крайне редко, так что про промежуточных сеймурий прочитать можно и сейчас. У сеймурии, кстати, много родственников, например, более поздняя, но и более примитивная Kotlassia prima, или синхронный, но более прогрессивный Discosauriscus. Сеймуриаморфы, с одной стороны, имели личинок с жабрами, что видно по отпечаткам раннепермского Ariekanerpeton sigalovi, боковую линию, местами даже в костном канале, как у рыб, что мы знаем по черепам Raphaniscus tverdochlebovae, один крестцовый позвонок – у Seymouria baylorensis. С другой же стороны, спина у этих существ была покрыта костными пластинками, шейных позвонков у той же сеймурии было целых пять, а во взрослом виде она вела полностью наземный образ жизни. Внешне сеймуриаморфы, скорее всего, были похожи на помесь саламандры с вараном.
Близки к сеймуриаморфам или даже включаются в них Chroniosuchidae (или Chroniosuchia), например, Chroniosaurus. У этих странных амфибий, как и у большинства рептилий, в черепе появилось дополнительное разгрузочное окно, но не за глазами, в перед ними, вдоль челюстного края; не зря один из видов назван Chroniosuchus paradoxus. Спина их была покрыта панцирем из широких последовательных пластин, отчего хрониозухиды похожи на микрокрокодильчиков. Вообще, обилие бронированных пермских амфибий (а их известно гораздо больше, чем тут перечислено) говорит само за себя.
Некоторых животных очень трудно определить как амфибий или рептилий, систематика их крайне спорна. Иногда их объединяют в группу Diadectomorpha (или Diadecta), иногда распределяют по другим группам. Образцовый пример – раннепермский Diadectes – тяжеловесное, коротконогое, приземистое трехметровое существо. Его передние зубы преобразовались в колышковидные резцы, растопыренные редким веером, а задние расширены подобно молярам млекопитающих, да к тому же трехбугорковые – крайне необычный для амфибий вариант. Судя по очень большой высоте и укороченности нижней челюсти, эти животные были растительноядными. Потребление растений в пищу входило в моду.
Маленькая тонкость
На макушке черепа Diadectes красуется огромное круглое отверстие. На самом деле, такое отверстие крайне характерно и для многих лопастеперых рыб, и для большинства архаичных амфибий, и для многих и многих рептилий, в том числе современных. Это глазница для третьего глаза, сделанного на основе верхней части промежуточного мозга. Известны два основных варианта третьего глаза – из эпифиза и из парапинеального тела. У многих современных амфибий и рептилий такой глаз тоже присутствует. Например, у лягушки-быка Lithobates catesbeianus, ящериц анолисов Anolis или игуан Cyclura третий глаз прекрасно виден в виде пятнышка между обычными глазами, а у гаттерии Sphenodon punctatus он снабжен хрусталиком и всеми прочими атрибутами полноценного глаза. У современных трехглазых животных этот орган вполне распознает свет и тьму, а у ископаемых, судя по размерам глазниц, мог быть еще полноценнее.
Вероятно, также растительноядным был раннепермский Bolosaurus striatus из Техаса, чей череп был треугольным при виде как сверху, так и сбоку, передние зубы торчали вперед, а задние несли как острый бугорок, так и расширенную площадку. Еще оригинальнее зубы у уральского Belebey: передние зубы в виде странно изогнутых резцов, за ними следовали мелкие «премоляры», а задние поперечно растянутые «моляры» на верхней челюсти имели острую вершинку спереди и плоскую поверхность сзади, а на нижней наоборот – площадку спереди и острый крючок сзади, так что вместе они образовывали отличную перетирающую молотилку, обогнавшую млекопитающих на целый период.
Вероятный родственник диадектеса – раннепермский двухметровый Limnoscelis paludis был, напротив, хищником, о чем говорит его треугольная голова с огромными клыками в передней части верхней челюсти.
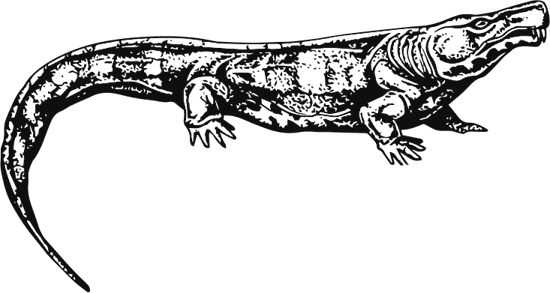
Limnoscelis paludis
Позднепермские Lanthanosuchus watsoni, Lanthaniscus efremovi и Chalcosaurus из Татарстана и Оренбургской области – необычные не то амфибии-антракозавры, не то рептилии-парарептилии. Их полуовальные черепа совершенно плоские и покрыты тончайшей ажурной и очень красивой сеточкой. Что ж, Поволжье славится разными кружевами – наличниками, пуховыми платками, морозными узорами, так что черепа лантанозухов тут очень в тему.

Lanthanosuchus watsoni
Как бы ни старались пермские амфибии и полурептилии, настоящие пресмыкающиеся уже развернулись в полную силу.
В пермских отложениях Техаса найдено древнейшее яйцо, причем его скорлупа, судя по структуре, была не известковая, а кожистая. Именно этого не хватало амфибиям, именно это вывело рептилий на первые места в эволюционной гонке.
В перми усовершенствовались карбоновые пресмыкающиеся и возникли многие новые, например, Captorhinida, Mesosauria и Millerosauria.
Капториниды, например Captorhinus и Labidosaurus, внешне самые скучные, типа «ящерок» с большими головами и короткими хвостами. Но и среди них есть странные создания. У Gecatogomphius kavejevi нижняя челюсть крайне расширялась спереди назад, причем в передней части была снабжена колышковидными клыками, а в задней каждая половина усыпана пятью рядами круглых зубов-пупырышков, за что зверушка и получила свое имя. Такими челюстями можно было здорово хватать и давить жестких насекомых или наземных моллюсков.
Мезозавры с границы карбона и перми, представленные несколькими видами рода Mesosaurus, – водные рептилии, на реконструкции выглядящие как брутальные аналоги крокодилов. Есть, правда, тонкость: мезозавры совсем маленькие – примерно метр с хвостом, причем хвост был длиной со все остальное тело. Судя по утяжеленным ребрам, мезозавры хорошо ныряли, а судя по крупным задним лапам и высоким отросткам на хвостовых позвонках, они довольно быстро гребли в воде, причем довольно длинная шея позволяла вертеть головой. Очень узкая вытянутая морда была усыпана щеткой тончайших зубов; учитывая, что вся голова длиной около 10 см, каждый зуб был едва ли не с волос толщиной. Большой вопрос – кого можно ловить такими зубами? Если рыбу, то не переломает ли она эти зубы-волоски, когда будет дрыгаться? Да и как ее потом снимать с зубов, когда она на них наколется? Возможно, мезозавры ели не позвоночных, а мелких рачков, не кусая, а фильтруя воду через зубное сито; тогда их логичнее сравнивать не с крокодилами, а с усатыми китами. Замечательно, что почти одинаковые мезозавры известны из Южной Африки и Бразилии.
Систематическое положение мезозавров непонятно. Иногда их сближают с капторинидами и пеликозаврами, котилозаврами или парарептилиями, иногда ставят особняком и даже выделяют в специальную группу Proganosauria.
Миллерозавры – небольшая, непонятная, но важная группа, поскольку ее члены претендуют на звание великих предков для черепах. Примитивные члены, типа Milleretta rubidgei, похожи на ящериц, но с очень широкими ребрами. Самый же интересный миллерозавр – Eunotosaurus africanus из поздней перми Южной Африки. У него было приплюснутое овальное тело, ширина которого была почти равна длине, с очень широкими ребрами, образовывавшими почти сплошной панцирь, маленькая бронированная голова, мощные растопыренные лапы и очень длинный хвост. Судя по мелким колышковидным зубам, эунотозавр был растительноядным или всеядным. Предполагается, что такие существа рыли норы в рыхлом грунте (нормальных трав и, соответственно, прочного дерна еще и в помине не было), своды которых регулярно обваливались им на спины. Для защиты от этой неприятности позвоночник и спина усилились, ребра раздались и срослись с ороговевшей кожей. Это было тем проще, что многочисленные прецеденты, как уже говорилось, известны даже среди амфибий. Правда, эунотозавр – не единственный претендент на звание предка черепах, причем более вероятные претенденты жили гораздо позже, в триасе. У многих из них ребра были заметно у?же, чем у эунотозавра, так что его специализация, вероятно, слишком обогнала свое время.
Маленькая тонкость
Сейчас понятно, что височные окна могли возникать независимо в разных группах. Появлялись они и среди анапсид, хотя, казалось бы, отсутствие окон – черта, давшая им название. Например, височные окна по-разному развиты у разных видов даже одного рода Mesosaurus; среди миллерозавров у Milleretops их нет, а у Milleretta и Millerosaurus – есть, среди парейазавров у большинства нет, а у Bradysaurus baini есть. То же можно сказать и о многих других группах анапсид.
Другие потенциальные предки черепах – парарептилии Cotylosauria, в частности – Procolophonia (или Procolophonomorpha). Большинство из них выглядит как простые ящерицы, например, Nyctiphruretus acudens, но была и у них фишка – отсутствие ребер в поясничном отделе, прямо как у млекопитающих; другая продвинутая черта – три крестцовых позвонка. Проколофоны появились в конце перми и существовали весь триас. Растительноядный башкирский Rhipaeosaurus tricuspidens с его бугристыми щеками и характерными листовидными зазубренными зубами мог быть предком парейазавров, хотя имел удивительно длинную для парарептилий шею.
Одни из самых характерных растительноядных котилозавров поздней перми – Pareiasauria. Это были тяжеловесные коренастые животные длиной от метра до трех, с короткими мощными ногами и короткой широкой головой. Особый колорит парейазаврам придают бугристые развернутые «щеки» и множество пупырышков по всей поверхности головы. Вся спина тоже была покрыта окостеневшими бляшками. Характернейшая черта парейазавров – здоровенные отростки снизу нижней челюсти – рычаги для жевательных мышц, помогавшие пережевывать массу растений. Этому же способствовали зубы с бугристым краем и длинными корнями. Есть несколько предположений, как жили парейазавры. Некоторые палеонтологи считали их сугубо наземными, другие предполагали, что они не вылезали из воды, болтаясь там как огромные надувные матрасы. Судя по найденным следам ног, всё же парейазавры могли прогуливаться по грязным берегам, но изящно скакать они были точно не способны.
Самые известные парейазавры – Scutosaurus с нашего российского севера, но есть и масса иных, например Pareiasaurus и Deltavjatia vjatkensis. На весь мир знаменито Котельничское местонахождение в Кировской области, в котором сохранились целые скелеты в стоячем положении: когда болота подсыхали, парейазавры вязли в липкой трясине и погибали в безуспешных попытках выбраться. Вытянувшиеся и изогнутые позы скелетов красноречиво свидетельствуют об этих трагедиях. Часто верхние части скелетов разрушены падальщиками, а иногда и сами падальщики тоже тонули в грязи рядом с парейазаврами.
Самый странный парейазавр – карликовая Elginia mirabilis из Шотландии, выраставшая всего до 0,6 – 1 м длины. Вся голова эльгинии покрыта острыми шипами, а в стороны и назад растопырены настоящие рога. Забавно, что голова эльгинии очень напоминает внешность шотландской породы коров хайленд – низкорослых, с бахромой на морде и с точно такими же рогами.
Наконец, в перми начался расцвет нормальных пресмыкающихся – диапсид. Araeoscelis были стройными длинношеими «ящерками» с длинными тонкими ногами, бодро лазящими по деревьям в погоне за насекомыми. Такой образ жизни логично завершился появлением Weigeltisauridae (или Coelurosauravidae) – рептилий, крайне похожих на современных летучих драконов Draco из Юго-Восточной Азии. По бокам тела у вейгельтизаврид торчали очень длинные специальные косточки, похожие на ребра, образующие изящные «крылья», на которых можно было парить, прыгая с дерева на дерево. Длинными тонкими пальцами с острыми коготками вейгельтизавриды цеплялись за кору при приземлении, вернее, придревнении. Самые красивые вейгельтизавриды найдены на Мадагаскаре – Weigeltisaurus jaekeli и Coelurosauravus elivensis. Задняя сторона их изящного треугольного черепа была расширена в виде гребешка с красивым фестончатым краем.
В конце перми появляются Lepidosauria – группа, включающая современных клювоголовых Rhynchocephalia и чешуйчатых Squamata. Первые лепидозавры – Eosuchia – предки гаттерий и ящериц, представлены южноафриканской Youngina gracilis и мадагаскарским Thadeosaurus colcanapi, внешне совсем невыразительными «ящерками», выделяющимися разве что очень большими стопами. Последняя тенденция еще больше усилилась у мадагаскарских Claudiosaurus germaini и Hovasaurus boulei. Эти существа стали активно плавать, гребя огромными задними ногами-ластами и длиннющим плоским, сплющенным с боков, высоким хвостом. У клаудиозавра маленькая голова вертелась на не слишком длинной, но уже удлиненной шее; в будущем это дало плоды у потомков – завроптеригий, то есть плиозавров и плезиозавров.
В сакмарском веке Оклахомы жил Orovenator mayorum – уже очень-очень близкая к ящерицам рептилия. Наконец, в конце пермского периода появились и настоящие чешуйчатые Squamata – ящерицы Lacertilia. Самая древняя достоверная ящерица найдена в Архангельской области – Lanthanolania ivakhnenkoi, с границы поздней перми и раннего триаса Южной Африки известна Paliguana whitei, долгое время державшая звание древнейшей ящерицы.
Но это все были цветочки. Самый ужас зародился в самом конце перми – возникла группа Archosauromorpha. Примитивнейшие представители – татарский Eorasaurus olsoni, южноафриканский Aenigmastropheus parringtoni – были небольшими ящерицеподобными животными, но эволюция шла неслыханными темпами. Тут же образовались Archosauria, а внутри групы – Thecodontia, первым из которых был Archosaurus rossicus из России и Польши. У этого двухметрового хищника конец морды был коварно загнут вниз для удержания добычи. В какой-то степени это, конечно, была компенсация слабости челюстей и зубов, но компенсация очень успешная, которая вывела в следующем периоде архозавров на пик могущества. Но до их торжества на планете успела порулить другая группа – наши родственники и предки, зверообразные рептилии.
А где же люди? Первые звероящеры
Зверообразные рептилии, как и полагается знатной группе, имеют несколько альтернативных названий, из которых самые ходовые Synapsida и Theromorpha. В некоторых вариантах систематики между этими понятиями вводятся различия, в некоторых это синонимы. Классификация их сложна и запутана, даже в работах одного автора-специалиста, вышедших в разные годы, бывают существенные расхождения на эту тему. Приведенная далее система – одна из возможных, заведомо упрощенная.
В первом приближении зверообразные делятся на две группы: более примитивных Pelycosauria и намного более совершенных Therapsida. Особенно стоит подчеркнуть, что зверообразные рептилии хотя и существовали на всем протяжении перми, дали значимое разнообразие только в конце периода.
Зверообразные рептилии эволюционировали во все ухудшающемся пермском климате. В холоде выживали только рептилии с повышенным обменом веществ, способные регулировать температуру своего тела. Для того чтобы выделять много энергии, нужно ее много получать. Для этого сперва менялась зубная система: среди одинаковых колышкообразных зубов сначала выделялись более крупные клыки, а потом и прочие зубы разделялись на классы.
Маленькая тонкость
Зубные системы бывают разные. У подавляющего большинства рыб, амфибий и рептилий все зубы в челюстях одинаковые. Если же тип питания совершенствуется и усложняется, могут появиться разные классы зубов. Тогда зубная система называется гетеродонтной – «разнозубой». Обычно передние зубы становятся шпателевидными хватательными или щипательными, а задние – острыми разгрыза тельными или уплощенными жевательными. Иногда между передними и задними еще добавляются клыки – немногочисленные, зато очень большие и острые, приспособленные либо для убивания добычи, либо, если животное растительноядное, для красоты. У современных рептилий гетеродонтность – редкостное исключение, но у ископаемых встречается весьма регулярно. Гетеродонтные зубные системы возникали не раз, но только у зверообразных рептилий и млекопитающих стали базовым вариантом по умолчанию.
Увеличение мощности челюстей сделало череп высоким и узким. Челюсти зверообразных рептилий стали способны совершать жевательные движения сбоку-набок – пережевывание пищи в разы увеличивает усваивание полезных веществ. Из-за этого изменилось все основание черепа, строение нижней челюсти и нижнечелюстного сустава. Для боковых движений необходимы вынесенные вбок мышцы – появились скуловые дуги. Для пущей прочности при жевании число костей в нижней челюсти стало уменьшаться. В какой-то момент появились и щеки, чтобы пища не вываливалась изо рта.
Маленькая тонкость
Одна из ярких особенностей продвинутых зверообразных рептилий – широко расставленные скуловые дуги. У «стандартных» рептилий, как уже говорилось, в черепе обычно есть некоторое количество окон, между которыми расположены костяные перемычки, которые, на самом деле, тоже могут рассматриваться как скуловые дуги, только вот они обычно не выгнуты вбок, а потому называть их «дугами» несколько странно. Так или иначе, к ним тоже крепятся мышцы, только не «жевательные», а скорее «кусательные». За счет специфического расположения «дуг» у диапсидных рептилий (в том числе ящериц и динозавров) повышается точность укуса, но несколько снижается его мощность. Такой вариант возник еще в карбоне для охоты за насекомыми – легкими, но верткими. Зверообразные же заняли в большей степени экологическую нишу охотников на крупную тяжелую добычу, для которой точность укуса была не так важна, а сила – весьма актуальна. В последующем это приспособление удачно пригодилось для развития сначала всеядности, а потом и растительноядности, породив по пути экзотические варианты типа пермских аномодонтов Tiarajudens eccentricus и Anomocephalus africanus – со всеядными «резцами» спереди, неимоверными прямыми саблезубыми клыками в задней части верхней челюсти и плоскими «молярами» на нёбе.
Побочным эффектом стало изменение слуха. Раньше барабанная перепонка была натянута между черепом и нижней челюстью, а звук передавался на нее с почвы, так как у малоэнергичных рептилий голова обычно лежала на земле. Теперь же и голова стала подниматься выше на полувыпрямленных передних лапах, и барабанную перепонку нельзя было прикреплять по-прежнему, так как челюсть стала двигаться не только вверх-вниз (кстати, размах открывания пасти тоже увеличился), но и вбок. Из-за этого барабанная перепонка заглубилась в слуховой проход, образованный височной костью и бывшими элементами нижней челюсти; раньше они образовывали сустав, а теперь стали барабанной пластинкой и двумя дополнительными слуховыми косточками – молоточком и наковальней. Вся система стала громоздкой и тяжеловесной, отчего ее эффективность снизилась. В компенсацию добавилась слуховая раковина – рупор, чтобы окончательно не оглохнуть.
Раз слух стал хуже, стали усиливаться другие органы чувств, отвечающие за обоняние и осязание. Обонятельный эпителий, располагавшийся в верхней части ротовой полости, для лучшей эффективности стал отделяться от этой полости дополнительной пластинкой – твердым нёбом, постоянно нараставшим от передней стороны челюстей назад. Твердое нёбо разделило носовую и ротовую полости. Хоаны – задние носовые отверстия, соответственно, стали тоже уезжать назад, заодно позволяя одновременно жевать и дышать. В головном мозге обонятельные луковицы заметно увеличились.
Осязание усилилось за счет странных выростов на коже – вибрисс. О их наличии у зверообразных рептилий мы знаем по усилению кровоснабжения и иннервации костей черепа в тех местах, где у современных млекопитающих располагаются «усы» – вибриссы. Появление же вибрисс стало возможным благодаря активной работе кожных желез, вырабатывающих большое количество белка-кератина. Помогло то, что зверообразные рептилии отделились от прочих в тот самый момент, когда рыбья чешуя наконец-то исчезла, а рептилийная еще не появилась. В этом смысле гладкая кожа тероморфов более амфибийная, чем у многих пермских амфибий. Активно эволюционировали и другие кожные железы, став в последующем сальными, потовыми и молочными.
Обработка многократно увеличившегося потока информации не могла не вызвать увеличения размера мозга и усложнения его строения.
Мало усилить обмен веществ и добыть много энергии, хорошо бы эту энергию не терять зазря. Раз уж появилась система выращивания длинных отростков на коже, возникновение шерсти было просто следующим логичным шагом. Волосы – одновременно и осязательные элементы, и отлично сохраняют тепло, а по совместительству выполняют массу других функций – пугают, привлекают, скрывают и помогают держаться детенышам.
Маленькая тонкость
Выросты на коже могут образовываться самыми разными способами. Кожные зубы акул и чешуя костных рыб, «волосы» волосатой лягушки и чешуя рептилий, вибриссы и волосы млекопитающих – это всё разные по устройству образования. Если чешуя рептилий и перья птиц – производные поверхностных слоев кожи, иногда заглубляющиеся внутрь, волосы зверей – образования глубоких слоев, выступающие наружу.
Чтобы добыть больше пищи, надо быстрее и активнее двигаться. Ползая на брюхе, много не наловишь, благостные времена ленивых «живых капканов» уходили в прошлое. Особенно активно поменялся плечевой пояс, передние лапы стали полувыпрямленными, хотя до совершенства млекопитающих, конечно, еще было далековато. Правда тут тероморфы попались в стандартную эволюционную ловушку: в тот момент, когда способ бега стал круче, чем у конкурентов, отбор автоматически прекратился. Лапы зверообразных, по нашим нынешним меркам, остались весьма несовершенными, но в перми это был самый писк моды, вершина эволюции. Принцип «и так сойдет» – ведь и от таких кривоногих зверообразных никто убежать уже не мог – сработал, как обычно. Эта недоработка аукнулась тероморфам уже в конце триаса, когда самые продвинутые архозавры-текодонты изобрели бег на задних ногах. Двуногим динозаврам зверообразные уже ничего не могли противопоставить; но это было намного позже.
Особенно чудесно, что все эти изменения происходили постепенно, а благодаря изобилию материала мы можем проследить за деталями многих изменений. У ранних пеликозавров из вышеперечисленного почти ничего не было, а у поздних терапсид имелся полный комплекс, так что их уже довольно трудно отличить от примитивных млекопитающих.
* * *
Как уже было сказано, зверообразные рептилии в первом приближении разделяются на две большие группы – примитивных пеликозавров и продвинутых терапсид.
Маленький Tetraceratops insignis из кунгурского века Техаса имел короткий и высокий череп с двумя парами коротких бугристых недорожек – на носу и над глазами, а дополнительные пупыры торчали в задней части нижней челюсти. Зубы тетрацератопса выглядели просто, но были очень разной длины – большие спереди и перед глазами и мелкие в промежутках. Тетрацератопс, хотя и довольно своеобразен, и хронологически, и морфологически удачно совмещает признаки пеликозавров и терапсид. Очень похоже выглядел и Eothyris parkei – тоже с короткой мордой и парой огромных клыков на каждой стороне верхней челюсти, но уже более определенно относящийся к пеликозаврам.
Большинство пеликозавров Pelycosauria не слишком выразительны, но есть и выдающиеся формы.
Самый базовый вариант пеликозавров – Ophiacodontia, например Ophiacodon – коротконогие, большеголовые, приземистые твари с очень короткой шеей. Не исключено, что офиакодоны жили рядом с водой и питались рыбой. Очень похожи на них Varanopsidae (или Varanopidae) – Varanops brevirostris, Varanodon agilis и Aerosaurus – незамысловатые ящерицеподобные хищники.
Специализация многих пеликозавров пошла по пути изменения спины. Отлично это видно в ряду пеликозавров Sphenacodontia: Haptodus – Sphenacodon – Ctenospondylus – Dimetrodon. У первого из них позвоночник вполне обычный, у второго остистые отростки позвонков несколько удлинены, у третьего они уже очень длинные, а у последнего образуют высоченный парус на спине. Между отростками позвонков, конечно, была натянута кожа, так что вся система работала по тому же принципу, что и у платигистрикса. Судя по всему, у пеликозавров такие гребни возникали несколько раз независимо, а это явно говорит о несовершенстве их терморегуляции.
Всех этих существ объединяет форма головы: узкая, высокая, горбоносая, с короткой мордой, изогнутым зубным краем верхней и нижней челюстей, смещенными назад глазницами и очень маленьким мозговым отделом. От всего этого возникает впечатление, будто голова постоянно наполовину повернута на нас, должен же где-то быть мозг! Судя по острым зубам, среди коих некоторые выделялись в условные клыки, диметродон и его многочисленные родственники были хищниками. Сейчас нам может показаться странным, как существа с такой громоздкой конструкцией на спине вообще способны кого-то поймать, но всегда стоит помнить, что добыча была достойна хищников – кривоногие тормознутые плюшки, даже если могли вовремя заметить надвигающегося супостата (трудно не заметить полутораметровый парус!), всяко не успевали ретироваться. Показательно, что с появлением, с одной стороны, более бронированной и шустрой добычи, а с другой – более развитых хищников, гребнистый вариант почти полностью пропал; впрочем, некоторые подобные пеликозавры могли дожить и до начала триаса, а среди динозавров намного позже возникли полные аналоги.
Среди родственников диметродона были свои оригиналы: Secodontosaurus, хотя тоже имел гребень на спине, обладал крокодилоподобной – длинной и узкой – мордой.
В своем роде параллельной линией развития стали пеликозавры Edaphosauria. Примитивные Glaucosaurus megalops и Mycterosaurus longiceps были стройняшками безо всяких спинных украшений и с банальными колышковидными зубками без особых вывертов. Их потомки Edaphosaurus заметно увеличились – до 3 м, потяжелели, перешли на питание растениями и отрастили на спине гребень, но не треугольной, как у диметродонов, а полукруглой формы. При этом на остистых отростках эдафозавров по бокам торчали короткие поперечные перекладинки неочевидного назначения. Родственник эдафозавра Lupeosaurus kayi тоже имел парус на спине, но без поперечных перекладинок. Учитывая, что родственники-иантазавры с гребнем жили уже в конце карбона, даже среди эдафозавров такая специализация могла возникнуть трижды.
Работа над ошибками
Высокие спинные гребни диметродона, эдафозавра и их родственников столь колоритны, что их профиль очень часто вставляют в цепочки образов наших предков – «от бактерии до человека». Существует масса рисунков и мультиков, где сменяющие друг друга существа превращаются один в другого и в конце – в человека. И обязательно где-то в этом ряду мелькает пеликозавр с гребнем, который чудесным образом преобразуется в млекопитающего с обычной спиной. Нет! Такая специализация бесследно не проходит. Гребнистые формы не были нашими прямыми предками, они полностью вымерли, а у наших пращуров спина была самой банальной.
Едва ли не более удивительная особенность пермских эдафозавров – их зубная система. При взгляде на череп сбоку ничего необычного не видать – просто ряд мелких тупых однообразных зубов. Однако значительная часть нёба и внутренней поверхности нижней челюсти была усеяна многочисленными мелкими зубами, образовывавшими прямо-таки бугристые пластины для пережевывания растений. Неспроста и скуловые дуги эдафозавров не в пример шире, нежели, скажем, у диметродонов.
Совсем несуразно выглядели родственники эдафозавров Caseidae – Casea, Cotylorhynchus и Alierasaurus ronchii: непропорционально мелкая, перекошенная, абсолютно плоская сверху голова, тяжелое туловище, короткие растопыренные ноги и длиннющий хвост. Особый колорит этим животным придавали выступающий нос и убегающая, скошенная нижняя челюсть намного короче верхней, а также завернутые назад зубы – выражение лица, как у деревенского дурачка. Казеиды были растительноядными – одни из первых крупных позвоночных вегетарианцев.
* * *
Вторая большая группа зверообразных – Therapsida – намного богаче и разнообразнее. Самый их простой вариант – Eotitanosuchia (они же Phthinosuchia, или Biarmosuchia). Образцовый исходный представитель – Eotitanosuchus olsoni (он же Ivantosaurus ensifer) из вордского века. Этот здоровенный – 2,5 м длиной – хищник имел мощный череп с огромными саблезубыми клыками. Подобный образ оказался почти универсальным и в последующем не раз воспроизводился в позднепермских зверозубых рептилиях. Близкий родственник эотитанозуха, только чуть меньшего размера – Biarmosuchus. У мелкого Lemurosaurus pricei были огромнейшие глазницы – едва ли не в полчерепа. Некоторые подобные животные отрастили на голове разнообразные шишки, чаще над глазами и по средней линии головы, но также на скуловых дугах – Proburnetia viatkensis, Burnetia mirabilis, Lobalopex mordax, Paraburnetia sneeubergensis и прочие. Черепные выросты явно не имели прямого функционального назначения, а стало быть – были результатом полового отбора. На их поверхности есть следы увеличенных кровеносных сосудов, что связано либо с терморегуляцией, либо с изменением окраски, либо с ростом вибрисс и волос. Мохнатые бугры должны были выглядеть весьма оригинально. Есть даже предположение, что жесткая шерсть образовывала нечто наподобие носорожьих рогов.
Здорово и то, что очень похожие и явно близкородственные животные обнаружены на Урале и в Южной Африке, причем в областях, где климат был довольно прохладным. Возможно, именно по этой причине хвост у эутитанозухий, равно как и у прочих терапсид, был заметно укорочен – это снижало теплопотери.
Не только огромными размерами, но и названием замечателен Dinosaurus murchisoni, описанный еще в 1840 г., когда слово «динозавр» еще не «приросло» к тем архозаврам, которых сейчас все знают под этим именем. Правда, в последующем были попытки уничтожить такую красоту, заменив ее на Rhopalodon или синонимизировав динозавра с описанным еще в 1838 г. Brithopus priscus, но первая замена вообще незаконна, а бритопус описан по плечевой кости, так что произвольно соединять его с черепом динозавра как-то странно.
Странноголовые Dinocephalia (или Deinocephalia) в первом же приближении разделяются на Titanosuchia и Tapinocephalia. Некоторые титанозухии были хищниками, например Syodon и Titanophoneus potens, внешне мало отличающиеся от эотитанозуха, а другие – растительноядными. Среди последних особо эффектны два вида уральских Estemmenosuchus – E. mirabilis и E. uralensis. Головы этих трехметровых тварей были украшены короткими ветвистыми рогами, огромными бугристыми выростами на скулах и шишкой на лбу, а клыки, несмотря на растительноядность, были огромными, прямо-таки саблезубыми. Что характерно, самки подобных украшений не имели, отчего первоначально даже получили собственные названия – Anoplosuchus tenuirostris и Zopherosuchus luceus. Очевидно, рога и клыки были созданы половым отбором, а это говорит о сравнительно сложном брачном поведении. Надо думать, впечатляющий внешний вид дополнялся и соответствующим поведением – демонстрациями и боданием. Известны кожные окостенения и окаменевшие участки кожи эстемменозухов и эотитанозуха, по ним видно, что наружный покров был гладкий, без чешуи и шерсти, но с защитными уплотнениями и многочисленными слизистыми железами.
Маленькая тонкость
Образ коренастого растительноядного животного – саблезубого и со странными выростами на черепе – возникал много раз. Кроме пермских зверообразных эстемменозухов, это млекопитающие диноцераты Uintatherium, свиноподобные Archaeotherium и даже гигантские павианы Theropithecus brumpti. Остается только удивляться, как в разные времена и вроде бы даже в разных условиях могли появиться столь похожие твари.
Тапиноцефалы сменили эстемменозухов в экологической нише крупных растительноядных. Очень массивным сложением отличались трехметровые южноафриканские Moschops capensis и Tapinocephalus atherstonei, а также Ulemosaurus из Татарстана, чьи лбы были вздуты десятисантиметровыми шишками. Совсем уж странно выглядела голова Styracocephalus platyrhynchus – с длинным рылом, огромным вздутым лбом и широким костным «воротником» в задней части черепа. У Struthiocephalus whaitsi морда была не только длинная, но и плоская, чтобы можно было захватывать как можно больше еды. У него и более короткорылого Keratocephalus moloch мощные лобные бугры, возможно, были покрыты роговым чехлом, образуя настоящие рога.
Странно была устроена жевательная система тапиноцефалов. Из-за того, что полноценное твердое нёбо еще не сформировалось, и хоаны все еще располагались в передней части ротовой полости, одновременно и жевать, и дышать этим животным было трудно. А растительноядность требует постоянного жевания. Решением стало расширение передних зубов, на внутреннем крае которых возникла широкая «пятка» для перетирания еды; верхние и нижние зубы очень плотно сходились друг с другом в шахматном порядке. Задние зубы при этом оставались мелкими. Получается, что зубная система была как бы повернута задом наперед сравнительно с нашей, в которой как раз маленькие зубы расположены спереди, а большие расширенные – сзади. У Deuterosaurus между огромными резцами и мелкими «молярами» добавлялись огромные клыки, а на носу и над глазами возвышались шероховатые бугры, почти рога. Тапиноцефалы были главными крупными растительноядными вордского и кептенского веков.
Работа над ошибками
Часто понятия «растительноядный» и «травоядный» используют как синонимы, даже не задумываясь, тем более, что «траво-» говорить проще, чем «растительно-». На самом деле, разница велика. Любой «траво-» – «растительно-», но далеко не любой «растительно-» – «траво-». Ведь бывают листоядные, фруктоядные, плодоядные, семеноядные, нектароядные, смолоядные животные, те, кто ест цветы, стебли, кору, корни и корневища. Кто-то может есть все подряд, как овца, кто-то – утонченный гурман, как панда. По строению зубов или типу стертости иногда можно примерно угадать тонкости диеты, но для ископаемых животных это удается далеко не всегда, тем более что не всегда нам известна флора, окружавшая ту или иную зверюшку. На помощь приходит и общее сложение тела. Например, эстемменозухи и тапиноцефалы имели сильно поднятую переднюю часть тела, так что вряд ли они питались низкими растениями. Да и травы в современном виде в перми было не так много – лишь хвощи да плауны. Вытянутая широкая морда и сдвинутые назад и вверх глаза могли бы свидетельствовать о питании водорослями в воде, но ноздри находились на кончике морды, так что вероятнее питание вайями жестких папоротников и ветками колючих голосеменных.
Масса забавных существ объединяется в группу Anomodonta. Самые простенькие – Dromasauria, например, Galepus jouberti и несколько видов Galechirus. Эти тридцатисантиметровые существа были очень похожи на ящериц, но с большой вероятностью имели вибриссы. Согласитесь, усатая ящерица – это оригинально.
Более экзотичными были Venjukoviamorpha (или Venyukovioidea). Уральские Venyukovia prima (она же Venjukovia), Ulemica invisa, Suminia getmanovi и Otsheria netzvetajevi были мелкими животными с короткой головой и огромными глазами. Между прочим, как положено, на макушке располагался третий глаз, который у очерии был вынесен на высокую трубочку. Такое расположение говорит о двух вещах: во-первых, глаз был вполне функциональным, а во-вторых, с большой вероятностью уже имелась шерсть; по крайней мере, у современных северных и потому особо мохнатых копытных глазницы подобным же образом приподняты над поверхностью черепа. Особо странными были здоровенные передние зубы-резцы венюковиаморф, долотовидная форма которых наводит на мысли о грызунах; клыки были толстыми, но тупыми и короткими. На нижней челюсти вперед выдавался двойной волевой подбородок. На реконструкциях венюковиаморфы выглядят как пучеглазые «ящерицы-обезьяны», истинные рептилоиды. Суминия имела сравнительно длинные ноги с огромными кистями, большой палец которых противопоставлялся остальным; видимо, она умела неплохо лазать по деревьям.
Маленькая тонкость
Необычный источник сведений о жизни древних животных – копролиты. В ротовой полости молодого венюковиаморфа Niaftasuchus zekkeli найдены зубы взрослой особи с резорбированными основаниями, а также белесая масса, неотличимая от вещества копролитов. Судя по всему, молодые особи ели фекалии взрослых (в которых случайно оказались проглоченные старые сменявшиеся зубы), чтобы получить полезную микрофлору, так необходимую растительноядным животным.
Ценнейшая информация получена при изучении копролитов каких-то хищных зверообразных и архозавров из самого конца перми Вязников во Владимирской области. Судя по размерам и встречаемости остатков, больше всего тут нагадили тероцефал Moschowhaitsia vjuschkovi и Archosaurus rossicus. Сенсационна находка шерстинок в этих копролитах – прямое доказательство существования шерсти в перми, примерно 252 миллиона лет назад! В тех же копролитах обнаружены кусочки костей и чешуя рыб, причем у зверообразных рептилий кости переварены намного слабее, чем у архозавров. Дело в том, что у теплокровных животных обмен веществ активнее, пища задерживается в желудке ненадолго, а концентрация соляной кислоты там меньше, тогда как холоднокровные рептилии могут переваривать пищу буквально месяцами, да к тому же в более концентрированной кислоте. С одной стороны, именно по степени переваренности можно догадаться, кому принадлежали копролиты, с другой – это еще одно доказательно теплокровности зверообразных. Если шерстинки в желудках архозавров однозначно принадлежат жертвам, возможно, дицинодонтам, то зверообразные рептилии вполне могли вылизываться, как кошки, и проглатывать собственные волоски.
Известны и другие копролиты, например, растительноядных дицинодонтов, и совсем странные – звездчатые в сечении, кому они принадлежали, непонятно.
Линию странности продолжают следующие аномодонты – Dicynodontia (или Dicynodontoidea). Это были массивные коротконогие и короткохвостые животные с тяжелой головой. В конце перми они сменили тапиноцефалов в экологической нише медленных растительноядных. У примитивных представителей типа Cerataelurus mirabllis еще сохранялись многочисленные мелкие зубы, но уже выделялись огромные клыки. У нестандартных Endothiodon (или Emydochampsa) клыки исчезли, а мелкие зубки дополняли огромный изогнутый клюв, в который превратились челюсти; Oudenodon потеряли вообще все зубы, которые у них заменил острый, выступающий треугольником край челюсти. У большинства же дицинодонтов сохранились лишь два здоровенных клыка, за что, собственно, эти животные и получили свое название – «двуклыковые». Впрочем, известны индивиды без этих клыков, вероятно, самки, что однозначно указывает на декоративность клыков. Дицинодонты были растительноядными животными, срывавшими растения своими клювами и умевшими активно жевать, для чего по бокам черепа раздались очень широкие скуловые дуги и рельефные костные гребни. Особенно впечатляют скуловые дуги короткомордого Aulacephalodon – почти весь череп состоял из них. Поскольку зубы дицинодонтов почти или совсем исчезли, жевательные движения происходили в основном спереди-назад, так что скуловые дуги были не только раздвинуты вбок, но и смещены назад, за мозговую коробку, которая превратилась в крошечную перемычку между огромными костными арматуринами.
Размер дицинодонтов менялся в очень широких пределах. Карликовые Dicynodontoides (он же Kingoria) и Pristerodon mackayi были всего 60 см длиной, а название Elph borealis говорит само за себя. Самые стандартные дицинодонты – метровые африканские Dicynodon. Самые гигантские формы появились позже, в триасе.
Маленькая тонкость
Растительноядность – трудное дело. Растительная клетка имеет целлюлозную клеточную стенку, в растениях много жестких механических тканей, а иногда и ядов, тогда как питательных веществ может быть не так уж много. Переваривать целлюлозу трудно. Строго говоря, это научились делать лишь бактерии, грибы и некоторые простейшие, а растительноядные животные – это лишь бурдюки со встроенной сенокосилкой. Своими челюстями они размельчают растения, а в своих внутренностях создают оптимальную среду для жизни все тех же бактерий и простейших, которыми, собственно, в немалой степени и питаются. Отсюда следуют две основные особенности растительноядных, сочетающиеся в разной степени: во-первых, они должны иметь отличный жевательный аппарат, а во-вторых, большой пищеварительный тракт – желудок и кишечник. Необходимо время и на появление симбиотических микроорганизмов.
Из-за сложностей растительноядности наземные позвоночные так долго – весь девон и карбон – оставались хищными. Растения потреблялись только одноклеточными, грибами и в лучшем случае членистоногими. С другой стороны, появление крупных наземных растительноядных заметно изменило экосистемы. Оно повлияло и на эволюцию растений, и на состав почв, и на хищных позвоночных. Одно дело – гоняться за насекомыми, другое – уметь придушить саблезубого эстемменозуха или дицинодонта.
Диноцефалы пошли самым простым путем, нарастив размер и, стало быть, огромное брюхо. Венюковиаморфы и дицинодонты в этом смысле особенно важны, так как были сравнительно небольшими животными, грызунами своего времени. Это стало возможным благодаря эволюции жевательного аппарата, причем у венюковиаморф совершенствовались зубы, а у дицинодонтов – жевательная мускулатура. А множество мелких животных могут потребить намного больше растений, чем одно крупное. Тем более интересно, что как раз поздние дицинодонты ударились в гигантизм; собственно, это их и сгубило.
С большой вероятностью дицинодонты имели шерсть, возможно, это их волоски найдены в копролитах хищников из Вязников. Об эволюции теплокровности свидетельствует и насыщенность костей кровеносными сосудами. В то же время изотопный анализ костей показал, что пермские дицинодонты, вероятно, все еще были холоднокровными, тогда как продвинутые триасовые формы уже развили теплокровность. Учитывая данные по другим зверообразным, получается, что теплокровность возникала неоднократно и независимо.
Очень необычной была шотландская Geikia – мелкая, совершенно беззубая, с плоским лицом, квадратными скуловыми дугами, направленными вперед глазами, над которыми торчали тупые недорога. Почти столь же короткую и широкую голову, столь же беззубые челюсти и такие же огромные обращенные вперед глаза имел еще один карликовый южноафриканский дицинодонт, похожий на гнома, – Cistecephalus (он же Kistecephalus). Его коротенькие растопыренные лапки были пятипалы, но крайние пальцы резко уменьшились и, скорее всего, снаружи были вообще не видны, так что фактически функциональными оставались только два пальца, да и в тех часть фаланг срослась. Скорее всего, такие лапки были приспособлены для копания, хотя некоторые палеонтологи, учитывая бинокулярность, считают кистецефала древолазающим животным.
Полуметровый Diictodon feliceps знаменит не только малыми размерами, но и способностью рыть спиральные полутораметровые норы, в которых и находят их многочисленные скелеты. Норы располагались группами около мест с изобилием еды, но не соединялись между собой; дииктодоны были единоличниками. Эти «сурки» были так успешны, что в Южной Африке примерно половина находок всех пермских позвоночных – именно останки дииктодонов.
Работа над ошибками
Дицинодонты – очень колоритные животные, их мощные клыки и тяжелые черепа отлично сохраняются, поэтому исследователи часто их находят и описывают. Причем описывают чаще, чем надо бы. Например, в роде Dicynodon описано более 160 видов, из которых валидных – то есть признаваемых и адекватных – на самом деле всего парочка. Часть из представленного сонма – одно и то же, часть принадлежит к другим родам. С другой стороны, практически каждый род дицинодонтов имеет огромное количество синонимов, причем палеонтологи имеют разные представления об их самостоятельности. Разобраться в этой катавасии очень сложно.
Самая продвинутая группа зверообразных – зверозубые Theriodontia, они включают как самых впечатляющих наземных хищников поздней перми, так и самых невзрачных и мелких зверьков – наших прямых Великих Предков.
Gorgonopsia (или Gorgonopida) – очередная вариация на тему гигантского саблезубого хищника. Kamagorgon ulanovi из казанского века, переходный от эотитанозухов к горгонопсам, имел череп 45 см длиной. Nochnitsa geminidens из конца кептенского века Кировской области – наиболее примитивная истинная горгонопсия, но совсем маленькая, а потому названа в честь злобного ночного духа. Самый известный и самый крупный представитель горгонопсов – Inostrancevia. У I. alexandri череп достигал полуметра длины, а у I. latifrons и I. uralensis – даже 60 см, а это в полтора раза больше, чем у белого медведя! С хвостом зверюга была не меньше 3,5 метров, в холке – около метра, а ее клыки имели длину 15 см. Показательно, что при обилии добычи пермские зверообразные хищники не превзошли этой величины. Крокодилы и динозавры в более поздние времена без проблем отъедались до гораздо более приличных масштабов. Причиной может быть теплокровность горгонопсий: большой обмен веществ требует очень много еды, особенно в холодном климате, тогда как на жаре теплокровному труднее остыть. Известно великое множество похожих существ, например, вдвое меньшие, чем иностранцевии, Sauroctonus и Pravoslavlevia parva. Еще умереннее выглядел Lycaenops ornatus с его сравнительно тонкими, длинными и более-менее выпрямленными ногами. Это был позднепермский аналог волка.
* * *
Наконец, самая важная группа зверозубых – Eutheriodontia (они же Therosauria, или Eutherapsida). В первом приближении они делятся на Therocephalia (или Scylacosauroidea) и Cynodontia (или Galesauroidea). У них появилась масса продвинутых черт, большей частью связанных с усилением жевательных способностей. Развилось длинное вторичное нёбо, так что хоаны, наконец, отодвинулись далеко назад, и можно было долго и вдумчиво жевать. Височная мышца стала больше, что вызвало целый каскад изменений. Височное окно соединилось с глазницей, задняя часть нижней челюсти изогнулась наверх в виде восходящей ветви с отдельным венечным отростком – рычагом для височной мышцы, которая стала крепиться на сагиттальный гребень на макушке, который поднялся на том месте, где зияла теменная глазница, которая тут же у большинства эутериодонтов исчезла. Зубы окончательно разделились на резцы, клыки и щечные; исчезли нёбные зубы.
Вероятно, у самых продвинутых появились губы, а стало быть – потенциальная возможность сосать молоко (впрочем, современные однопроходные лижут молоко и без губ). Нижнечелюстной сустав стал почти как у млекопитающих, возможно, появилось наружное ухо. Скелет стал легче, ноги прямее, существа бодрее, а для этого, вероятно, возникла диафрагма, отделившая брюшную полость от грудной и давшая еще одно направление растяжения легких и, стало быть, углубившая вдох. По крайней мере, ребра в поясничном отделе исчезли. Самое главное – сильно увеличилась мозговая коробка, что явно говорит о том, что зверозубые поумнели. Понятно, что все эти изменения произошли не вдруг; здорово, что обилие материала позволяет проследить детали и этапы эволюции.
Therocephalia – очень важная группа, так как это – неудачная попытка стать млекопитающими. Эти мелкие животные выглядели как помесь саламандры, ящерицы и крысы. Голова в целом повторяла форму горгонопсий, но в уменьшенном, приплюснутом и вытянутом варианте; теменной глаз сохранялся, но был уже совсем крошечным, тесним формирующимся сагиттальным гребнем; вторичное нёбо было зачаточным, хоаны все еще располагались в передней части ротовой полости. Несмотря на это, один из древнейших тероцефалов – южноафриканский кептенский Glanosuchus macrops – уже имел носовые раковины, подобные таковым у млекопитающих, а также среднее ухо, почти строго промежуточное между рептилийным и млекопитающим вариантами. Большей частью тероцефалы были мелкими насекомоядными зверьками. Известно великое изобилие тероцефалов, но большинство из них «на одно лицо» – Ictidodraco longiceps, Rubidgina angusticeps, Ictidosuchops intermedius, Lycideops longiceps и многие-многие прочие: вытянутая совершенно плоская, как катком прокатанная, голова, острые зубки, крупные клыки, очень большие глаза.
На общем фоне выделяется Euchambersia mirabilis, за клыками которой располагались огромные полости с желобком, ведущим к основанию клыка. Скорее всего, в полости размещалась ядовитая железа. Видимо, следствием ядовитости было и исчезновение заклыковых зубов и скуловой дуги у эучамберсии – отравленная добыча не могла сопротивляться, так что на ее удержание приходилось тратить намного меньше сил.
Названием замечателен Gorynychus masyutinae: назван он в честь Змея Горыныча, но одновременно играет на созвучии с английским словом «гори» – «окровавленный» и греческим «онихус» – «коготь». Этот зверь размером с волка жил в конце перми на территории нынешней Кировской области и был самым крупным хищником котельнической фауны, тираня тамошних парейазавров.
Были и водоплавающие тероцефалы. В водах плескались Viatkosuchus sumini и рыбоядный Annatherapsidus petri размером с выдру, а также совсем мелкий Perplexisaurus foveatus, имевший зато очень большой мозг и электрорецепторные органы на морде, с помощью которых он находил моллюсков и прочую беспозвоночную мелочь в придонной мути.
Тероцефалы самого конца перми – Moschowhaitsia vjuschkovi и Megawhaitsia patrichae – заметно подросли (у последнего вида череп достигал полуметра) и вытеснили горгонопсов из ниши главных хищников, хотя сами все еще были довольно примитивны и не слишком прытки, о чем явно свидетельствуют растопыренные в стороны короткие лапы. Каналы в челюсти мегавайтсии могут быть следами ядовитых протоков. Если это так, то понятно, почему у нее не было саблезубых клыков – они просто были не нужны. Между прочим, ядовитость сама по себе говорит о теплокровности жертв – дицинодонтов, ведь холоднокровное животное с низким обменом быстро от яда не погибнет и может далеко убежать от хищника.
Cynodontia – прямые предки млекопитающих. Самые ранние и примитивные цинодонты – Charassognathus gracilis из вучапинского века Южной Африки, а также немного более поздние и продвинутые Procynosuchus из Германии, Замбии и Южной Африки еще имели расположенные спереди хоаны. Это были совсем мелкие зверюшки, длиной 25 – 50 см, скорее всего, насекомоядные. Чуть крупнее была Dvinia prima (она же Permocynodon sushkini) из самого конца перми – вятского века Архангельской области. Ее зубы были очень оригинальны: острые резцы, «предклык», клык и задние зубы, причем верхняя зубная дуга намного шире нижней, так что верхние зубы не смыкались с нижними, а только плотно ухватывали добычу, тогда как нижние давили ее о нёбо. Задние зубы овальные, плоские, с множеством крупных бугорков по периметру и мелких в центре. Носовая полость двинии уже была устроена почти как у млекопитающих, с разделением решетчатой и верхнечелюстной носовых раковин и, соответственно, верхнего обонятельного и нижнего дыхательного носовых ходов. Некоторые палеонтологи считают это приспособлением для нагревания вдыхаемого воздуха и, стало быть, признаком теплокровности.
Как вы там, потомки?
Поздние зверообразные рептилии очень похожи на млекопитающих, но они всё же не млекопитающие. В строении их черепа есть масса примитивных особенностей, из которых самые внешне заметные и очевидные – теменное отверстие для третьего глаза и неслившиеся носовые отверстия. Есть, конечно, и масса других рептилийных черт, например, в строении плечевого пояса: головка плечевой кости образовывала сустав одновременно с лопаткой, коракоидом и прокоракоидом, впереди от лопатки сохранялся клейтрум – наследство еще от рипидистий, а по средней линии между ключицами – межключица. В целом такой плечевой пояс больше походит на странно повернутый таз и выглядит неоправданно усложненным даже на фоне современных рептилий, не говоря уж о млекопитающих, у которых из всего этого богатства обычно сохраняется лишь лопатка, да у самых примитивных (людей, например) – ключица. О размножении зверообразных рептилий мы знаем немного, но нет особых сомнений, что они откладывали яйца.
И вот ведь диво: современные однопроходные, или яйцекладущие млекопитающие Prototheria, они же Monotremata, по многим и многим признакам оказываются ближе к тероморфам, нежели к нормальным зверям Theria. У однопроходных – утконоса Ornithorhynchus anatinus, ехидны Tachyglossus aculeatus и трех видов проехидн Zaglossus – в плечевом поясе по-прежнему есть прокоракоид, коракоид и межключица, разве что клейтрум наконец слился с лопаткой, хотя еще и не стал полноценной лопаточной остью. Передние и задние лапы однопроходных и поставлены, и двигаются как у рептилий. Есть у них и надгрудинник, и подвижные шейные ребра, и предлобные и заднелобные кости в черепе, и клоака, и размножаются они, о ткладывая яйца. У них нет губ, а теплокровность очень несовершенна. Весьма вероятно, что рептильный нижнечелюстной сустав был утрачен, а молоточек и наковальня приобретены однопроходными независимо от прочих млекопитающих. Некоторые зоологи прямо предлагают включать их в тероморфов. Всё же обычно их относят к млекопитающим по нескольким причинам: они покрыты шерстью, они худо-бедно, но теплокровны, они кормят детенышей молоком (хотя молочных желез с соском у них нет), в мозге развит неокортекс, а в шее семь позвонков, молочные зубы утконоса похожи на зубы многобугорчатых млекопитающих.
* * *
Гондванскую гляциоэру завершил позднепермский гляциопериод, состоящий из двух ледниковых эпизодов – от конца кунгурского до середины казанского века и в середине татарского века, то есть в конце вордского и кептенском веке. Оледенение захватило не только Гондвану, как в предыдущие разы, но и северо-восток Азии, которая сдвинулась на север.
Похолодание сопровождалось аридизацией, то есть повышением сухости. Сначала это не слишком сказывалось на экосистемах, в которых все еще было много водных животных, но под конец перми осушение пошло уже всерьез, так что огромное число амфибий и зависимых от воды рептилий исчезло бесследно. Вслед за стегоцефалами, парейазаврами и дицинодонтами в могилу сошли и гигантские зверообразные хищники. Понятно, что пострадали не только позвоночные. Суммарно вымерли 70 – 75 % видов наземных животных, а насекомых – 83 % видов.
Но это все ерунда по сравнению с тем апокалипсисом, что творился в морях. Пермское вымирание – самое грандиозное за всю историю планеты. За несколько десятков тысяч лет – по геологическим меркам ничтожный срок – исчезли 50 % семейств, 70 % родов и 96 % видов морских организмов! Окончательно и бесповоротно пропали все трилобиты, ракоскорпионы, кораллы ругозы и хиолиты. Почти полностью исчезли табуляты, но это не спасло мшанок, сохранившихся по остаточному принципу и проигравших своим вечным конкурентам кораллам. На грани полного вымирания оказались брахиоподы, уступив место донных фильтраторов двустворчатым моллюскам. Сильно пострадали морские лилии и морские рыбы.
Неспроста граница эр – палеозойской и мезозойской – проводится именно тут.
Столь масштабный катаклизм должен был иметь грандиозные причины; ясно, что ученые соревнуются в изобретении поводов сгинуть целым экосистемам.
Понятно, что первым делом в голову лезут ужасные катастрофы, и в них недостатка действительно не было.
Чего стоит хотя бы извержение вулканических Эмэйшаньских траппов и траппов в Западной Сибири! Бездны лавы, изливающейся из бездонных трещин, тянущихся от горизонта до горизонта, выбросы ядовитых газов, разрушение озонового слоя и выжигание всего живого вдесятеро усилившимся ультрафиолетом, пожары и пепел, покрывающий столь плодородные в прошлом равнины. Ужас! Одна проблема: в непосредственном соседстве с сибирскими траппами тысячи лет процветала богатейшая флора и фауна, не зная о том, что ей положено в ужасе вымирать в рекордные сроки. Непосредственно в вулканических базальтах пермско-триасовых траппов на плато Путорана полно насекомых, причем это не следы одномоментной гибели, жизнь кипела в таком режиме тысячелетиями.
А может, причина в метеорите или астероиде? Что ж, подходящий кратер всегда найдется, их на Земле сотни. Например, кратер диаметром в 500 км около Земли Уилкса – отличный кандидат и по времени, и по масштабу.
А что, если археи Methanosarcinales научились наконец перерабатывать органику с выделением метана и слишком увлеклись процессом? Газ тысячелетиями накапливался на дне морей, потихоньку травя там все живое. Но с наступлением потепления случилось ужасное: метан одномоментно всплыл несказанным смертоносным пузырем. Такой адский выброс даже пафосно назван метангидратным ружьем. Бабахнуло «ружье» так, что дышать на планете стало нечем. ОБЖ в школах еще не ввели, скомандовать «газы» было некому, вот все и задохнулись.
Что сказать, сценарии впечатляющие и достойные экранизации в многосерийном фильме-катастрофе. Но может, хватало и обычных причин, просто сложившихся вместе?
Ранее единая Пангея трескалась, опять расползаясь в разные стороны. Пик оледенения быстро сменился потеплением. Концентрация кислорода в воде упала. Одновременно потепление высвободило огромное количество пресной воды, а соль уже ушла в материковые отложения, отчего океан стал намного преснее прежнего. Из-за тектонических движений уровень моря в одних местах понизился, в других – вырос. Уже этого набора во все времена хватало для того, чтобы все умерли, но в перми появился еще один источник проблем.
Как уже говорилось, ко второй половине перми появилось множество насекомых с водными личинками – поденки, веснянки, ручейники и прочие. Личинки накапливали азот и фосфор в воде, а взрослые насекомые разлетались и разносили ценные вещества по водоразделам. Удобрение почв привело к успеху наземной растительности, заселившей былые пустыни. Деревья – а в перми это уже полноценные голосеменные – пускали свои могучие корни и закрепляли почву, что снижало размыв. Азот и фосфор и так безостановочно выносились из водоемов, так еще и перестали туда возвращаться. В сочетании с падением концентрации кислорода и опреснением это дало катастрофический эффект – продуктивность морей несказанно снизилась. Очевидно, что все это произошло не вдруг, а заняло тысячи и миллионы лет. До некоторых пор прочности биосферы хватало, но количество имеет свойство переходить в качество. Тут-то все и вымерли.
* * *
Пермь – время торжества палеозойской биоты и ее фатального краха. Все заделы, появившиеся еще в кембрии, ордовике, силуре, девоне и карбоне, реализовались в перми в полную силу. Планета наконец-то покрылась полноценными лесами, насекомые развернулись во всю мощь своих крылышек, четвероногие наконец-то освоили все возможные экологические ниши, кроме разве что воздушной. Пермские холода дали нам волосы и теплокровность, разные классы зубов и способность жевать, вторичное нёбо и две новые слуховые косточки, наружные уши и отличное обоняние, развитый мозг и сложное поведение. Ноги стали выпрямляться, а спина училась гнуться в вертикальной, а не горизонтальной плоскости. Казалось, до появления настоящих млекопитающих оставался один шаг, но ужасный катаклизм – потепление и вымирание – разрушил все надежды.
* * *
Альтернативы
Обилие пермских существ поражает. При этом надо помнить, что мы знаем лишь малую часть реального разнообразия, ведь большая часть находок происходит из Приуралья и Южной Африки, а что в это время происходило на остальной планете – покрыто мраком. Каждый полевой сезон приносит массу открытий, и темпы обновления наших знаний зависят только от усердия и возможностей палеонтологов, земля же скрывает еще тысячи удивительных окаменелостей. Но даже среди изученных существ есть масса кандидатов на лучшую судьбу, чем их постигла в реальности.
Великое множество рептилизовавшихся амфибий, среди коих первое место держат сеймуриаморфы, запросто могли реализовать свой потенциал лучшим образом. Им бы еще десяток-другой миллионов лет на каком-нибудь маленьком личном континентике – и настоящим рептилиям противостояла бы серьезная конкуренция. Но Пангея на то и «пан», что своих континентиков не оказалось, а условия даже в удаленных частях не слишком различались. Впрочем, несходство фаун даже из близких местонахождений показывает, что расстояние имело большое значение для расселения неторопливых амфибий и даже рептилий.
Среди зверообразных рептилий известно великое множество групп и жизненных форм. Суминии и, возможно, некоторые дицинодонты уже были готовы стать рептилио-приматами, развили бинокулярное зрение, хватательную кисть и всеядность – все важнейшие качества, отличавшие наших предков в начале кайнозоя. Конечно, лучшие альтернативные кандидаты на «маммализацию» – превращение в млекопитающих – тероцефалы. Какие-нибудь крысоподобные Nikkasaurus tatarinovi или барсукоподобные Hexacynodon purlinensis были в полушаге от настоящих зверей. Вибриссы, шерсть, теплокровность, большой мозг – не хватало только живорождения (а про это мы знаем слишком мало – может, оно уже у кого-то и появилось?).
Как жаль, что позднепермский кризис выкосил большую часть этих чудесных созданий. Однако, погибли не все. Спасшиеся основали новый, лучший мир. Жаль только, что нашим предкам в нем на долгие миллионы лет была уготована жалкая роль закуски. Наступал мезозой…
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК