Глава 5. Нуклеиновые кислоты и молекулярная репликация
Глава 5. Нуклеиновые кислоты и молекулярная репликация
Теперь, когда мы в несколько абстрактных выражениях описали требования к живой системе, мы должны подробнее рассмотреть, как осуществляются различные процессы в тех организмах, которые мы видим повсюду. Как мы уже видели, основное требование, безусловно, заключается в наличии некоторого довольно точного метода репликации и, в частности, копирования длинной линейной макромолекулы, образованной из стандартного набора субъединиц. На Земле эту роль играет то или иное из двух больших семейств нуклеиновых кислот: семейство ДНК и семейство РНК. Общее строение этих молекул является чрезвычайно простым, действительно, настолько простым, что ясно говорит о том, что они действительно восходят к самому началу появления жизни.
ДНК и РНК довольно похожи, они, можно сказать, молекулярные кузины, поэтому давайте сначала опишем ДНК, а затем то, чем отличается от нее РНК. Одна цепочка ДНК состоит из единообразного остова, последовательности атомов, повторяющейся снова и снова, с присоединенной при каждом повторе боковой группой. Химически в остове содержатся ...фосфат-сахар фосфат-сахар... и т.д., и так повторяется многие тысячи или даже миллионы раз. Сахар — это не тот сахар, что стоит у вас на обеденном столе, а более мелкий, который называется дезоксирибоза, то есть рибоза с одной отсутствующей «окси»-группой (следовательно, название ДНК означает Дезоксирибонуклеиновая Кислота; «нуклеиновая» — потому что найдена в ядре высших клеток, «кислота» — из-за фосфатных групп, каждая из которых в нормальных условиях несет отрицательный заряд). К каждому сахару присоединена одна боковая группа. Боковые группы различны, но всего насчитывается лишь четыре их основных типа. Эти боковые группы ДНК (по формальным причинам называемые основаниями) удобно обозначить по первым буквам их названий А, Г, Т и Ц (что означает, соответственно, Аденин, Гуанин, Тимин и Цитозин). Благодаря своему точному размеру и форме, а также характеру химических составляющих, А изящно соединится в пару с Т, а Г — с Ц. (А и Г большие, Т и Ц поменьше, поэтому каждая пара состоит из одного большого и одного малого оснований.)
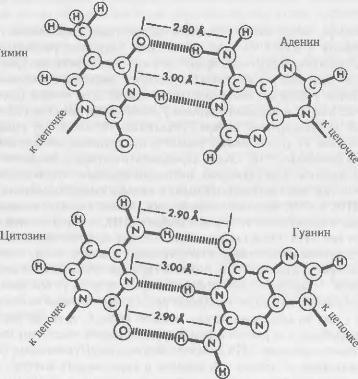
Пары оснований, которые образуют секрет структуры ДНК Основания соединены слабыми водородными связями, показанными прерывистыми линиями Тимин всегда объединяется с аденином; цитозин с гуанином.
И ДНК, и РНК довольно легко образуют двуцепочечные структуры, в которых обе цепочки лежат рядом, бок о бок, переплетаясь друг с другом для образования двойной спирали и связанные воедино своими основаниями. На каждом уровне существует одна пара оснований, образованная основанием одной цепочки, спаренного (на основе правил спаривания) с основанием другой. Каждая из связей, соединяющая эти пары друг с другом, довольно слаба, хотя в совокупности они образуют достаточно устойчивую двойную спираль. Но если структура нагревается, то возросшее тепловое возбуждение оттолкнет цепочки в сторону с тем, чтобы они разделились и отплыли друг от друга в окружающей их воде.
Генетическое сообщение передается точной последовательностью оснований вдоль одной цепочки. Тогда, зная эту последовательность, можно считывать последовательность ее комплементарной соседки, используя правила спаривания оснований (А с Т, Г с Ц). Генетическая информация записывается дважды, один раз на каждой цепочке. Это может оказаться полезным, если одна из цепочек повреждена, поскольку ее можно восстановить, используя информацию — последовательность оснований — другой цепочки.
Здесь есть одна неожиданная особенность. В обычной двойной спирали оба остова обеих цепочек не приблизительно параллельны, а антипараллельны. Если последовательность атомов в одном остове быстро увеличивается, то во втором — уменьшается. Это приводит к определенным сложностям, но не таким значительным как можно ожидать. По сути, это вытекает из того типа симметрии, которым обладает двойная спираль. Он создается псевдосимметрией спаривания оснований. Оказывается, что для этих конкретных химических соединений это удобный способ точно совпадать друг с другом.
Легко понять, что молекула такого типа, состоящая из пары цепочек, нерегулярные элементы которых (основания) совпадают друг с другом, идеальна для молекулярной репликации, особенно потому, что обе цепочки можно довольно легко отделить друг от друга щадящими методами Это происходит потому, что связи внутри каждой цепочки, скрепляющие каждую цепочку, являются прочными химическими связями, довольно невосприимчивыми к обычному тепловому разрушению, тогда как обе цепочки удерживаются вместе довольно слабыми связями, так что их можно без значительных затруднений отодвинуть друг от Друга, не нарушив при этом остов каждой из них. Две цепочки ДНК подобны двум любовникам, они удерживают друг друга в тесных объятиях, но их можно разделить, потому что как бы тесно они ни соприкасались друг с другом, у каждой есть единство, которое сильнее связей, их объединяющих.
Поскольку они столь точно совпадают друг с другом, то одну цепочку можно считать матрицей другой. Основной механизм репликации очень просто понять. Обе цепочки разделяются. Затем каждая цепочка действует как шаблон для сборки новой парной цепочки, используя в качестве сырья запас из четырех стандартных элементов. Когда эта операция завершится, мы будем иметь две пары цепочек вместо одной, и поскольку для того, чтобы выполнить работу аккуратно, сборка должна подчиняться правилам спаривания оснований (А с Т, Г с Ц), то последовательности оснований обязательно будут точно скопированы. В итоге мы получим две двойные спирали, где раньше имели лишь одну. Каждая дочерняя двойная спираль будет состоять из одной старой цепочки и одной вновь синтезированной цепочки, точно совпадающих друг с другом, и что более важно, последовательности оснований этих двух дочерних цепочек окажутся идентичными последовательности первоначальной родительской ДНК.
Основная идея вряд ли может быть проще. Единственная довольно неожиданная особенность заключается в том, что обе цепочки не идентичны, а комплементарны. Можно представить даже еще более простой механизм, в котором одинаковое спаривается с одинаковым с тем, чтобы обе парные цепочки оказались идентичными, но характер химических взаимодействий, скорее, несколько облегчает точное соответствие друг другу комплементарным молекулам, нежели абсолютно идентичным.
Как подобный процесс выдерживает сравнение с более крупными механизмами копирования, распространенными сегодня? Строка набора, подготовленная для печати, состоит (или обычно состояла) из определенного числа стандартных символов, организованных в строку или ряд строк. У каждой буквы в этом шрифте есть одинаковый для всех букв стандартный элемент, который вставляется в бороздки, удерживающие эту литеру на месте, и элемент, который характерен для каждой буквы. На этом сходство кончается. В репликации ДНК нет ничего, что соответствовало бы полиграфической краске. Буквы, напечатанные на листе, являются зеркальными изображениями очка литеры, но не комплементом (который остаётся, когда очко литеры вошло), и, что самое важное, получившуюся на печати строчку нельзя потом вернуть на место в ту же машину, чтобы воспроизвести очко литеры. Печатные прессы выпускают многие тысячи экземпляров газет, но газеты не копируются обратно в набор.
Репликация ДНК совсем не такая. Для того чтобы заработал естественный отбор, важно, чтобы саму эту копию можно было скопировать. Репликация ДНК больше похожа на отливку фрагмента скульптуры из литейной формы, так как если она достаточно простая, то саму скульптуру можно использовать для создания дополнительной формы. Основная разница заключается в том, что нить ДНК строится лишь из четырех стандартных отрезков. Очевидно, что это не относится к большинству фрагментов скульптуры.
Если мы исследуем процесс репликации ДНК, то увидим, что здесь есть ряд основных требований. Если мы начнем с двойной спирали, то обе цепочки должны каким-то образом разделяться. В наличии должен иметься запас из четырех элементов, каждый из которых состоит из характерного участка остова, — одна молекула сахара объединяется с одной молекулой фосфата, — и включает одно из четырех оснований, присоединенное к сахару. Такая состоящая из трех частей молекула называется нуклеотид. На практике эти первичные элементы имеют не просто один фосфат, а три, расположенные в ряд, причем два других отделяются в процессе полимеризации, предоставляя таким образом энергию для проведения синтеза в желаемом направлении. Хотя можно представить процесс, проходящий без дополнительных элементов, в развитой системе мы непременно рассчитываем обнаружить, по крайней мере, один фермент (то есть белок с каталитической активностью), который ускорит синтез и сделает его более точным.
Таковы в общих чертах требования. Когда исследуешь реальную систему репликации, то обнаруживаешь, что она значительно сложнее. Прежде всего, когда начинается синтез, обе цепочки еще полностью не разделились. Синтез новых цепочек происходит в процессе разделения, поэтому некоторые части двойной спирали копируются еще до того, как разделились другие более удаленные участки. Есть особые белки, функция которых состоит в том, чтобы раскрутить двойную спираль, и вместе с другими, которые могут создать ники[4] в остове, дать возможность одной цепочке вращаться вокруг другой, и затем вновь объединить разорванную цепь. Так как обе цепочки двойной спирали быстро двигаются в противоположных направлениях, и так как, говоря химическим языком, синтез проходит только в одном направлении, то мы обнаружим, что синтез происходит в прямом направлении на одну из цепочек и в обратном направлении на другую, поэтому механизм должен учитывать эту сложность. Более того, новый фрагмент цепочки ДНК обычно начинается как небольшой отрезок РНК, с которым затем объединяется более длинный фрагмент ДНК. Существуют добавочные белки, которые затем вырезают этот праймер РНК и заменяют его эквивалентным отрезком цепочки ДНК, и затем объединяют все воедино без разрыва. Мы знаем, что для синтеза одного вида небольшого вируса, созданного из ДНК, требуется почти двадцать различных белков; какие-то из них выполняют одну функцию, какие-то — другую. Это очень характерно для биологических процессов. Лежащий в их основе механизм должен быть простым, но если процесс биологически важен, тогда в длительном ходе эволюции естественный отбор усовершенствует и приукрасит его, с тем чтобы он мог функционировать как быстрее, так и точнее. Именно вследствие этой причудливой усложненности биологические механизмы часто так трудно разгадать.
К счастью, как мы отмечали ранее, нам не нужно задерживаться на этих сложностях. Когда зародилась жизнь, химия, должно быть, была относительно простой. Здесь важно уяснить, что четкая геометрия пар оснований, которая лежит в основе правил спаривания, предоставляет удобный случай особой репликации, даже в совершенно простых системах. Мы видим, что решающей в ДНК является не ее двойная спираль. Действительно, простой вирус может иметь в качестве своего генетического материала единственную нить ДНК, и она может быть настолько коротка (длиной лишь в пять тысяч оснований), что ей не требуется вторая цепочка в качестве страховки от повреждения. Основная особенность заключается в том, что в механизме репликации должна использоваться простота особых пар оснований для построения новой цепочки с комплементарной последовательностью оснований по отношению к старой. Именно эта простота заставляет нас считать, что она использовалась в самых древних живых системах. Вопрос о том, остаются ли обе цепочки, новая и старая, вместе после репликации, менее важен.
Здесь мы должны сказать несколько слов о близкой родственнице ДНК — РНК. (Подробнее различные виды РНК описаны в приложении.) Как мы уже объясняли, генетическая информация в каждой клетке высшего организма закодирована как подробная последовательность оснований ряда очень длинных молекул ДНК. В любое время многие более короткие участки этой последовательности копируются на однонитевые молекулы РНК и используются клеткой в качестве рабочих копий. Некоторые из них используются для структурных целей, но большинство как информационные РНК — инструкции для синтеза белка. Это происходит в очень сложных молекулярных структурах, называемых рибосомами, и для этого необходим дополнительный молекулярный аппарат, в частности, набор транспортных молекул РНК (тРНК).
Это, безусловно, очень сложная, но сложна она, в основном, потому, что должна выполнять сложную функцию. Процесс создания однонитевой копии РНК участка ДНК, который называется транскрипцией, относительно прост, и, чтобы направлять его, необходим лишь достаточно крупный белок. Процесс синтезирования белка с использованием фрагмента информационной РНК в качестве инструкций, который называется трансляцией, обязательно сложнее, так как инструкции написаны на языке РНК, состоящем из четырех букв, а их следует перевести с помощью химического механизма на язык белка из двадцати букв. Действительно, очень удивительно, что такой механизм вообще существует, и еще удивительнее, что любая живая клетка, животного ли, растения или же микроорганизма, содержит его вариант. Его открытие явилось одним из триумфов молекулярной биологии.
Таким образом, клетка — это миниатюрная фабрика, ведущая быструю, организованную химическую деятельность. В условиях соответствующих молекулярных воздействий фермент деловито синтезирует отрезки информационной РНК. Рибосома запрыгивает на каждую информационную молекулу РНК, двигается вдоль нее, считывая ее последовательность оснований и соединяя друг с другом аминокислоты (которые доставили ей молекулы тРНК) с тем, чтобы создать полипептидную цепь, которая, по завершении этого процесса, свернется и станет белком. Природа изобрела сборочный конвейер за несколько миллиардов лет до Генри Форда. Более того, этот сборочный конвейер производит много разных, весьма специфических белков, механических инструментов клетки, которые сами создают и восстанавливают органические химические молекулы для того, чтобы обеспечить сборочный конвейер сырьем, а также все молекулы, необходимые для устройства структуры фабрики, снабжают ее энергией, избавляют от отходов и выполняют другие Функции. Поскольку все это так сложно, то читателю не обязательно пытаться разобраться во всех этих тонкостях. Важно осознать тот факт, что несмотря на то, что генетический код почти универсален, механизм, необходимый для его реализации, слишком сложен для того, чтобы появиться внезапно. Он должен был развиться из какого-то более простого. Несомненно, что главная задача в понимании происхождения жизни — это попытаться разгадать, какой могла быть эта более простая система.
На этом этапе, возможно, стоит сравнить и сопоставить эти три больших семьи макромолекул: белок, РНК и ДНК. Молекулы белка, которые строятся из двадцати различных боковых цепочек, некоторые из которых химически довольно активны, более универсальны как класс, по сравнению с молекулами нуклеиновых кислот. Именно по этой причине все известные ферменты созданы из белков, хотя в некоторых случаях могут понадобиться мелкие органические молекулы, чтобы работать одновременно с ними в качестве фермента. Именно способность каждого фермента создавать или разрушать определенные химические связи позволяет современным клеткам вообще функционировать. Поскольку многие различные химические реакции необходимо подобным образом катализировать, то существует много различных видов ферментов.
В противоположность этому, не найдена ни одна молекула нуклеиновой кислоты, выполняющая функции катализатора. Обе, и РНК, и ДНК, имеют только четыре типа боковых групп вместо двадцати, и, несмотря на то, что они идеальны для репликации, потому что их основания так хорошо совпадают друг с другом, эти боковые группы не подойдут для химического катализа. Но РНК и ДНК могут делать то, чего не могут белки, — образовывать комплементарные структуры типа найденной в двойной спирали. Мы не знаем способа, которым молекула белка могла бы сделать то же самое, ну и, конечно же, этого не сделает современный белок с его двадцатью различными видами боковых цепочек.
Большинство химиков, занимающихся проблемой происхождения жизни, считают, что в начале первой появилась РНК, а ДНК является более поздним изобретением. РНК химически реактивнее ДНК, и, вероятно, ее было легче синтезировать в первозданных условиях Земли. Самые первые гены, должно быть, были созданы из РНК. Только позднее, когда генетическая информация увеличилась в длину, появилась необходимость в более устойчивой ДНК для обеспечения архивной копии.
Жизнь, какой мы ее знаем на Земле, представляется синтезом двух макромолекулярных систем. Белки, благодаря своей универсальности и химической реактивности, выполняют все функции, но неустойчивы для репликации самих себя любым простым способом. Нуклеиновые кислоты, по-видимому, специально созданы для репликации, но кроме нее могут успешно выполнять довольно небольшое количество функций по сравнению с более сложными и лучше оснащенными белками. РНК и ДНК — эти немые ткани биомолекулярного мира, которые, в основном, подходят для воспроизведения (с небольшой помощью со стороны белков), но от них мало пользы во многих действительно необходимых вещах. К проблеме происхождения жизни было бы намного легче подойти, если бы существовала только одна семья макромолекул, способная выполнять обе функции, репликацию и катализ, но жизнь, как мы знаем, задействовала две семьи. Возможно, это происходит благодаря тому, что не существует макромолекул, которые могли бы выгодно выполнять обе функции вследствие ограничений, накладываемых органической химией, то есть вследствие природы вещей.
Для того чтобы продвинуться дальше вперед, мы должны попытаться что-нибудь узнать о химических и физических условиях на первозданной Земле или на любой другой похожей планете К этому мы сейчас обратимся.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 2. Молекулярная архитектура белков
Глава 2. Молекулярная архитектура белков Не будем скрывать: покончив с первой главой, авторы (а возможно, и читатель) испытали некоторое облегчение. В конце концов цель ее заключалась лишь в том, чтобы дать читателю сведения, необходимые для понимания последующих глав,
ДНК — МОЛЕКУЛЯРНАЯ ОСНОВА ГЕНОМА
ДНК — МОЛЕКУЛЯРНАЯ ОСНОВА ГЕНОМА Сколько истин, признаваемых нами в настоящее время бесспорными, в момент провозглашения их казались лишь парадоксами или даже ересями! Екатерина II Что мыслимо — то возможно, что возможно — то мыслимо. Г. Лейбниц Тонкое устройство
Размножение ДНК (репликация)
Размножение ДНК (репликация) Важно то, что структура ДНК, открытая Уотсоном и Криком, многое прояснила относительно разных механизмов функционирования этой молекулы в клетке. ДНК не только кодирует генетическую информацию, но и самовоспроизводится (удваивается) при
ДНК — МОЛЕКУЛЯРНАЯ ОСНОВА ГЕНОМА
ДНК — МОЛЕКУЛЯРНАЯ ОСНОВА ГЕНОМА Сколько истин, признаваемых нами в настоящее время бесспорными, в момент провозглашения их казались лишь парадоксами или даже ересями! Екатерина II Что мыслимо — то возможно, что возможно — то мыслимо. Г. Лейбниц Тонкое устройство
Размножение ДНК (репликация)
Размножение ДНК (репликация) Важно то, что структура ДНК, открытая Уотсоном и Криком, многое прояснила относительно разных механизмов функционирования этой молекулы в клетке. ДНК не только кодирует генетическую информацию, но и самовоспроизводится (удваивается) при
Глава 13 Молекулярная биология. протеин
Глава 13 Молекулярная биология. протеин Энзимы и коэнзимЫXX в. открывал все новые и новые детали метаболизма клетки. Каждая метаболическая реакция, как выяснилось, катализируется каким-то определенным энзимом. Для того чтобы понять природу метаболизма, нужно исследовать
Глава 14 Молекулярная биология. нуклеиновые кислоты
Глава 14 Молекулярная биология. нуклеиновые кислоты Вирусы и геныКак только молекулы протеина вошли под контроль науки, неожиданно обнаружилось, что на роль первородных кирпичиков жизни претендуют совсем иные, нежели предполагали ученые, структуры. Эти структуры вышли
17. Часы и нуклеиновые кислоты
17. Часы и нуклеиновые кислоты Толковый словарь определяет «творческое мышление» как переосмысливание предшествующего опыта для создания новых «образов», подводящих к решению той или иной проблемы. Прекрасный пример тому являет собой работа Эрета.В 1948 году, окончив
Глава 6. Молекулярная генетика
Глава 6. Молекулярная генетика Поиски причин – занятие, не имеющее конца. У каждой причины есть своя причина, и нам приходится изучать все более и более отдаленные моменты в прошлом. Н. Тинберген (1907–1988), выдающийся голландский биолог, один из основателей этологии, лауреат
Путь глюкуроновой кислоты
Путь глюкуроновой кислоты Доля глюкозы, отвлекаемой на метаболизм по пути глюкуроновой кислоты очень невелика по сравнению с большим ее количеством, расщепляемым в процессе гликолиза или синтеза гликогена. Однако продукты этого вторичного пути жизненно необходимы
2.2. Нуклеиновые кислоты
2.2. Нуклеиновые кислоты Нуклеиновые кислоты обеспечивают хранение и воспроизведение наследственной информации. Этим определяется их фундаментальное значение для жизни на Земле.Нуклеиновые кислоты – это полимеры, мономерами которых являются нуклеотиды. Нуклеотид
Репликация ДНК
Репликация ДНК Репликацией называется процесс удвоения молекул ДНК. Уникальная способность ДНК к самоудвоению определяет ее ключевую роль в живом организме. Репликация лежит в основе воспроизведения себе подобных живыми организмами, что является главным признаком
Глава 3 Морфологическая и молекулярная эволюция
Глава 3 Морфологическая и молекулярная эволюция Я думаю, что отец наш небесный выдумал человека, потому что разочаровался в обезьяне. Марк
МОЛЕКУЛЯРНАЯ ЭКОЛОГИЯ
МОЛЕКУЛЯРНАЯ ЭКОЛОГИЯ В прессе часто появляются заметки о том, что торговцы дикими животными пытаются продавать запрещенные виды или продукты, изготовленные из вымирающих видов животных, прикрываясь торговлей вполне легальным товаром.Один из способов разрешения
9. Органические вещества. Нуклеиновые кислоты
9. Органические вещества. Нуклеиновые кислоты Вспомните!Почему нуклеиновые кислоты относят к гетерополимерам?Что является мономером нуклеиновых кислот?Какие функции нуклеиновых кислот вам известны?Какие свойства живого определяются непосредственно строением и