Глава 7. Непосредственное действие электромагнитных полей на мозг
Глава 7. Непосредственное действие электромагнитных полей на мозг
Мозг как наиболее совершенная система управления и как орган тонкого приспособления организма к условиям окружающей среды привлекает пристальное внимание естествоиспытателей различных специальностей. Психологи изучают образование индивидуальных и видовых приспособительных поведенческих реакций; нейрофизиологи, регистрируя электрическую активность отдельных нейронов, хотят понять особенности формирования нейронных цепей; биохимики стремятся выяснить механизмы обучения и памяти на молекулярном уровне.
Любому исследователю, изучающему деятельность мозга, необходимо знание способов воздействия на центральную нервную систему, чтобы уметь объективно оценивать и нормальную деятельность мозга, и ее изменения под влиянием определенных внешних воздействий. Следует признать, что это необходимое предварительное условие в реальных исследованиях не может быть выполнено, так как нам еще неизвестны все параметры среды, определяющие деятельность мозга, условно принимаемой за «нормальную».
Более полно способы воздействия и регистрации функций головного мозга в естественных и искусственных условиях указаны в табл. 2.
До сих пор мы рассматривали результаты изучения деятельности мозга целостного организма при воздействии ЭМП-методами, отмеченными в верхних строчках таблицы. Нужно заметить, что среди воздействующих факторов почетное место занимает прямое электрическое раздражение мозга, но эта процедура связана с частичным разрушением мозга.
Большие надежды возлагают на способы воздействия, связанные с проникающими ЭМП разных параметров. В начале нашего века известный русский физиолог В. Я. Данилевский называл этот способ «действие электричества на расстоянии». В настоящее время электромагнитный способ воздействия на мозг менее разработан, чем контактный электрический способ. Однако дистантный путь воздействия сулит большие перспективы в будущем. Надежды на плодотворность этого пути исследования поддерживают недавние убедительные данные по регистрации электрических и магнитных полей вокруг живого мозга, о чем подробнее мы поговорим позднее. Пока зарегистрировать такие поля удалось на расстоянии нескольких сантиметров от мозга, но эти факты свидетельствуют о том, что «полевое» воздействие имитирует в какой-то степени естественные нервные процессы, а следовательно, такой путь исследований перспективен.
Следует заметить, что прямое изучение деятельности мозга при использовании биохимических и биофизических методов началось сравнительно недавно (примерно с середины XX в). Эти возможности появились и появляются в связи с общим научно-техническим прогрессом, затрагивающим в большей мере смежные области знания. К примеру, сегодняшние достижения в учении о мозге нельзя вообразить без учета возможностей радиоэлектроники, позволившей зарегистрировать и обработать различные биоэлектрические процессы мозга.
Таблица 2. Способы воздействия на мозг и методы регистрации его функций в интактном организме и в искусственных условиях
Объект Воздействие Реакции через органы чувств прямо на мозг Интактный мозг Социальные воздействия (речь), естественные раздражители (свет, звук, запах, вкус, осязание и т. д.), химические факторы, физические факторы Химические факторы, физические факторы (электричество, ЭМП, вибрация, ультразвук и т. д.) Субъективный отчет. Двигательные реакции. Секреторные реакции. Сосудистые реакции. Электрическая активность мозга. Биохимические реакции мозга Изолированный мозг Естественные раздражители, химические факторы, физические факторы Химические факторы, физические факторы Электрическая активность мозга. Биохимические реакции мозга Переживающая нервная ткань и культура нервной ткани — Химические факторы, физические факторы Электрическая активность мозга. Биохимические реакции мозга Мертвый мозг — — Морфологические, гистологические, цитологические и молекулярные измененияЛюбой здоровый человек может оценивать работу своего мозга по ощущениям. Он давно выделил пять основных органов чувств, эти своеобразные окна в мир, и, основываясь на здравом смысле, вполне резонно, на его взгляд, полагает, что разговоры о дополнительных источниках информации не нужны.
Особенность сегодняшнего этапа научного познания заключается в углубленном изучении всех природных явлений. Факты, лежащие на поверхности, уже получили свое объяснение. Как физика перешла к объяснению мира на невидимый невооруженному глазу атомный уровень, так и нейрофизиология начинает оперировать явлениями, недоступными прямым человеческим ощущениям.
Хотя мозг в настоящее время интенсивно изучается на разных уровнях его деятельности, наиболее перспективными считаются новые аналитические методы, крайним выражением которых является изучение изолированной культуры нервной ткани.
Детальное познание всех свойств живого мозга человека и животных является очень сложной задачей. Переплетения миллиардов нервных клеток, каждая из которых связана с тысячами других, только приблизительно характеризуют сложность проблемы. Сюда необходимо добавить десятки миллиардов клеток глиальных, ибо по современным воззрениям мозг без глии не может функционировать. Физиолог укажет также на необходимость учета функций кровеносных сосудов при анализе мозговой деятельности, на существенную роль спинно-мозговой жидкости (ликвор обновляется 5—6 раз за сутки!) и на массу других подробностей, усложняющих и без того сложную работу мозга.
Недавним приобретением науки считается взгляд, согласно которому структурно-функциональной единицей нервной деятельности следует считать не отдельный нейрон, а сосудо-глионейрональный комплекс.
История применения электрофизиологических методов в электромагнитной нейрологии еще коротка. Относительно изменения электрической активности спинного мозга под влиянием МП известно, что при частотах внешнего ПеМП, близких параметрам электрической активности спинного мозга спинальной лягушки, отмечалось увеличение амплитуды, а при других частотах ПеМП изменяло форму регистрируемых биопотенциалов.
При исследованиях электрической активности головного мозга человека отмечали ее изменения после воздействия МП, поля УВЧ и поля СВЧ. Влияние слабых ЭМП отмечали в эпизодических исследованиях коллективов физической ориентации. В одном исследовании белорусских физиков [Михайлова-Лукашева и др., 1972] на головы 7 испытуемым в возрасте 20—30 лет воздействовали 10—25 с слабыми перепадами ЭМП (10-14 Дж) частотой следования 2,8—10, 400 Гц когерентного излучения в полосе до 6 МГц. Регистрировали биоэлектрическую активность теменной области при биполярном отведении. Сравнивали средние значения интегральной плотности ЭЭГ до и после воздействия.
На ЭМП 400 Гц изменения ЭЭГ отмечали у 6 испытуемых, а на ЭМП 2—10 Гц — только у 4 из 7. Хотя детальных сведений о результатах экспериментов авторы не приводят, можно предполагать, что низкочастотные ЭМП (2—10 Гц) вызывали преобладание колебаний биопотенциалов в диапазоне дельта- и тета-ритмов, а ЭМП 400 Гц — преобладание высокочастотных колебаний в бета-диапазоне.
В сообщении украинских физиков отмечалось, что некоторые люди (3 из 10) могут воспринимать искусственные низкочастотные (0,01—2,00 Гц) МП с индукцией 2*10-7—2*10-6 Тл, судя по изменению выраженности медленных волн в их ЭЭГ. В загородных условиях, где высшие магнитные помехи были на 2 порядка ниже, чем в городе, отмечали большую чувствительность к МП. Изменения в ЭЭГ наступали не раньше чем через 1 мин после начала воздействия.
Более подробно изучали ЭЭГ-изменения у человека под действием ЭМП врачи-гигиенисты, отмечая чаще всего преобладание медленных волн в ЭЭГ людей, подвергающихся электромагнитному воздействию в условиях производства.
Наблюдаемые изменения являлись результатом длительных (иногда многолетних) воздействий ЭМП, параметры которых значительно варьировали. Отсюда выводы при исследовании ЭЭГ человека обычно заключались в констатации изменений, а более подробный физиологический анализ возникающей ЭЭГ-реакции можно было получить только в опытах на животных.
В качестве объекта исследований использовались обезьяны, собаки, кошки, кролики, крысы, голуби, саламандры, лягушки и беспозвоночные животные. Эксперименты на животных давали возможность вживлять электроды в различные отделы головного мозга и тем самым исследовать межцентральные отношения, изолировать и разрушать отдельные участки ЦНС, а также позволяли изучать реакцию «под лучом». Правда, на этом пути возникали методические осложнения, связанные с тем, что металлические электроды, концентрируя вокруг себя высокочастотные ЭМП, вызывали нагревание мозговой ткани. Для избавления от таких артефактов исследователи переходили к созданию съемных металлических электродов, к использованию угольных или стеклянных электродов, а также к применению низкочастотных ЭМП и постоянных полей, когда нагревание исключается. Электрофизиологический анализ влияния ЭМП на головной мозг чаще проводили советские исследователи.
В наших исследованиях, где изучались ЭЭГ-реакции на ЭМП, основные опыты проведены на неанестезированных кроликах. Хотя не исключена возможность, что другие животные могли быть более чувствительны к ЭМП, нельзя считать выбор объекта совершенно случайным: у животных с высокой степенью синхронизации мозговых потенциалов такие относительно слабые раздражители, как ЭМП, могли не повлиять на ЭЭГ. Важно подчеркнуть, что животные не подвергались никаким фармакологическим воздействиям и потому речь может идти о нормальных реакциях организма.
Длительность воздействия ЭМП чаще всего ограничивалась 1 или 3 мин с интервалом 10—20 мин. В отдельных экспериментах длительность воздействия варьировала от нескольких секунд до нескольких часов. Длительность эксперимента с каждым животным в зависимости от цели исследования менялась от нескольких часов до нескольких месяцев.
В 1960 г. были начаты электрофизиологические исследования влияния на мозг кролика полей УВЧ и СВЧ. При использовании съемных электродов отмечалось возникновение неспецифической (одинаковой при разных ЭМП) диффузной (возникающей сразу во всех участках мозга) ЭЭГ-реакции синхронизации, заключающейся в увеличении числа медленных волн и веретен в ЭЭГ. Наиболее интенсивная реакция на поле СВЧ с ППМ, около 40 мВт/см2, наблюдалась в коре больших полушарий, гипоталамусе и в неспецифических ядрах таламуса.
В подобных условиях опытов другие исследователи [Лапин, 1970] обнаружили следующую последовательность в интенсивности ЭЭГ-реакции на поле СВЧ, если перечислять в убывающем порядке: гипоталамус, кора, таламус, гиппокамп, ретикулярная формация.
Недавно появились сообщения о том, что модулированное (50 Гц) ЭПМ с частотой 39 МГц и напряженностью 30—400 В/м вызывает появление изолированной судорожной электрической активности в гиппокампе крыс после воздействия длительностью от нескольких секунд до 2 ч.
Эти интересные электрофизиологические данные, которые согласуются и с результатами, полученными условнорефлекторной методикой, показывают, что при воздействии ЭМП на ЦНС возникает реакция, характеризующаяся сложными межцентральными отношениями. Их изучение только начинается. Для проведения сравнительного анализа результатов, полученных разными авторами, важно, чтобы эксперименты проводились на животных одного вида и анализ осуществлялся по одному параметру (у крыс лимбическая система более реактивна к ЭМП, чем у кроликов).
Изучение реакции мозга на каждый новый раздражитель освещает какую-то неизвестную прежде грань в многогранной деятельности ЦНС, помогая тем самым глубже познавать особенности функционирования этой важнейшей системы организма. Здесь уместно описать подробнее реакции на ПМП, которые не вызывают нагревания, относительно легко создаются и часто встречаются в природных условиях.
В 60-х годах (публикации появились с 1963 г.) мы исследовали влияние ПМП, создаваемого электромагнитом, который питался постоянным током от аккумуляторов. Индукцию ПМП варьировали от 20 до 100 мТл. Длительность воздействия измерялась 1—20 мин. Направление силовых линий было горизонтальным. Регистрируя электрическую активность мозга, мы располагали голову кролика между полюсами электромагнита.
Во многих экспериментах одновременно с записью ЭЭГ регистрировали электрокардиограмму (ЭКГ) и частоту дыхания кроликов. Однако не удалось отметить изменений со стороны дыхания или сердечной деятельности при кратком воздействии ПМП. Можно полагать на этом основании, что запись ЭЭГ является достаточно чувствительным методом регистрации реакций на ПМП.
Если между полюсами электромагнита помещали туловище или таз кролика, то изменений в ЭЭГ при 1—3-минутном воздействии не наблюдали, что заставило предполагать непосредственное действие ПМП на ЦНС.
Многочисленные литературные данные доказывают существование и рефлекторного пути бездействия ПМП на ЦНС, но нас интересовало именно непосредственное влияние, так как в большинстве работ оно только отмечалось, но не анализировалось.
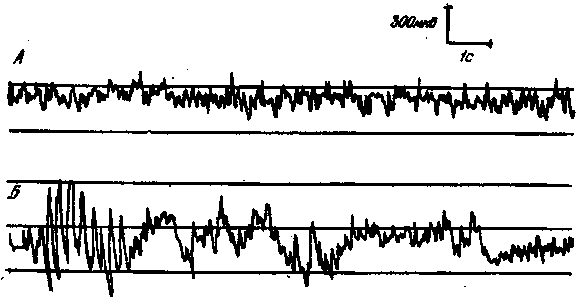
Рис. 9. Изменение ЭЭГ крысы под влиянием ПМП 100 мТ
А — фон; Б — во время действия ПМП
В опытах с ПМП 147 кроликам было дано более 5000 воздействий. При использовании ПМП возможное биологическое действие ЭДС индукции в момент включения и выключения электромагнита проверялось в опытах с постепенным включением ПМП (через реостат). Оказалось, что ЭЭГ-реакция на ПМП не зависит от скорости создания ПМП, а определяется интенсивностью ПМП и длительностью его воздействия. При длительности воздействия менее 20 с мы не обнаружила ЭЭГ-реакции на ПМП.
В отличие от привычной для электрофизиолога генерализованной реакции десинхронизации, возникающей с латентным периодом в доли секунды на применение обычных раздражителей (свет, звук, и т. д.) средней интенсивности, основная ЭЭГ-реакция на ПМП выражалась в достоверном увеличении числа веретен и медленных волн (рис. 9). Такая генерализованная неспецифическая реакция синхронизации выявлялась не при каждом применении ПМП (примерно в 60% случаев), а ее средний латентный период располагался в диапазоне 20—40 с.
Возникшая ЭЭГ-реакция длилась некоторое время и после выключения электромагнита. Часто последствие выражалось в дополнительном увеличении числа веретен и медленных волн, которое характеризуется нами как реакция на выключение. Эта реакция могла появляться независимо от основной реакции и обладала меньшим латентным периодом (в среднем около 15 с).
Автоматический частотный анализ ЭЭГ подтвердил результаты визуальной обработки, показав, что во время воздействия ПМП происходит обогащение ЭЭГ на медленных частотах и на тех, которые составляют веретена. Подобные результаты дал автоматический анализ ЭЭГ, записанный на протяжении 1 мин после выключения ПМП, подтверждая наличие эффекта последействия.
При регистрации электрической активности различных отделов головного мозга кролика было обнаружено, что реакции в виде увеличения числа медленных волн и веретен возникают одновременно во всех отведениях (при скорости записи 15 мм/с), а наиболее интенсивную реакцию наблюдали в гипоталамусе, затем следовала кора сензомоторная, зрительная, специфические ядра таламуса, неспецифические ядра таламуса, гиппокамп и ретикулярная формация среднего мозга. Можно заключить, что наиболее реактивными образованиями ЦНС при воздействии ПМП являются кора и гипоталамус.
Результаты данной серии экспериментов привели нас к предположению о том, что ПМП может влиять на любой участок головного мозга, но интенсивность реакции определяется его реактивностью. В нескольких опытах искусственно повышали реактивность, к примеру, ретикулярной формации среднего мозга путем внутривенного введения адреналина, и реакция этою участка мозга на ПМП становилась более интенсивной, чем реакции других отделов.
При физиологическом анализе обнаруженной реакции мы пытались выяснить, все ли участки тела кролика чувствительны к различным ПМП. Для этой цели воздействию попеременно подвергались задние ноги кролика, живот, грудь или голова. Понятно, что в этих экспериментах локализация ПМП была относительной, так как уменьшенное поле выходило за пределы интересующей нас области, а длительность невелика (1—3 мин). Но результаты опытов были достаточно определенны: изменения ЭЭГ мы наблюдали только в случае воздействия ПМП на голову.
В дальнейшем выяснялась роль в восприятии ПМП известных дистантных рецепторов, расположенных в головной части животного: зрительного, слухового и обонятельного. Хирургическим путем производилась та или иная деафферентация. Оказалось, что ЭЭГ-реакция сохранялась после разрушения дистантных рецепторов. Таким образом, целостность дистантных рецепторов не является необходимым условием воздействия ПМП на ЦНС.
Одностороннее разрушение ретикулярной формации среднего мозга, заднего вентролатерального ядра таламуса или заднего гипоталамуса не препятствовало возникновению ЭЭГ-реакций на ПМП и не вызывало асимметрии этих реакций. Можно предполагать, что одностороннее прерывание путей температурной, болевой, тактильной и проприоцептивной чувствительности не влияло на ЭЭГ-реакции при действии ПМП. Если вспомнить, что разрушение дистантных рецепторов также не влияло на осуществление этих реакций, то еще раз напрашивается заключение, что ПМП обладает преимущественно непосредственным действием на ЦНС. Проверку этого предположения мы решили провести на изолированных участках головного мозга.
Оказалось, что препарат изолированного мозга, получаемый после перерезки на уровне среднего мозга, реагирует на ПМП более часто, с меньшим латентным периодом и более интенсивно (иногда в ответ на воздействие возникали судорожные разряды), чем интактный мозг. ЭЭГ-реакция заключалась в увеличении числа медленных волн, веретен и судорожных разрядов.
Таким образом, опыты с препаратом изолированного мозга показали, что денервированные структуры промежуточного и конечного мозга могут реагировать на ПМП. Однако оставалось неясным, реагирует ли на ПМП целостная структура переднего мозга или любой участок мозговой ткани. Для ответа на этот вопрос были проведены опыты на нейронально изолированной полоске коры больших полушарий мозга кролика.
Полоску коры размером 5?15?5 мм мы изолировали в сензомоторной или зрительной области. Регистрацию электрокартикограмм (ЭКоГ) производили биполярным способом с помощью фитильковых электродов. В большинстве случаев регистрировали спонтанную электрическую активность полоски в виде нерегулярных или регулярных высоковольтных медленных потенциалов, а также в виде судорожных разрядов.
ЭКоГ-реакция на ПМП чаще выражалась в активации электрической активности. Эта реакция характеризовалась малым латентным периодом в сравнении с интактным мозгом и препаратом изолированного мозга, хотя по степени прочности ЭКоГ-реакция полоски на ПМП была близка к ЭЭГ-реакции интактного мозга. Реакция полоски коры на ПМП не зависела от места изоляции.
Настоящая серия экспериментов показала, что нейронально изолированная полоска коры реагирует на ПМП с меньшим латентным периодом, чем кора интактная, а следовательно, в условиях целостного мозга ЭЭГ-реакция на ПМП формируется при участии как коры, так и подкорковых отделов. Этот вывод подтверждается как формой ЭЭГ-реакции на ПМП целостного мозга, так и величиной ее латентного периода.
Электрическая реакция на ПМП может возникать в небольшом участке мозговой ткани, снабжаемом кровью через паутинную оболочку мозга. Возможная роль гуморального фактора в появлении изучаемых реакций исключалась в опытах по изучению электрической активности переживающей изолированной нервной системы беспозвоночных. Речь пойдет об электрической реакции на ПМП изолированного брюшного участка нервной цепочки речного рака.
Исследуемый препарат состоял из 8 ганглиев и соединяющих их коннектив брюшной нервной цепочки речного рака. Препарат помещали во влажную камеру с физиологическим раствором и с помощью серебряных электродов регистрировали спайковые разряды нейронов. Частота разрядов широко варьировала в зависимости от особенностей препарата, срока переживания, времени года и т. д.
50 препаратам было дано 103 3-минутных воздействия ПМП индукцией около 200,0 мТл. Наблюдали обратимое уменьшение частоты спайков. Латентный период реакции превышал 3 с. В остальных случаях изменения электрической активности цепочки были незначительными. Необходимо заметить, что характер эффекта ПМП зависел от фоновой частоты разрядов нейронов, торможение возникало при высокой исходной частоте. В контрольных опытах не удавалось зарегистрировать торможения электрической активности, которое возникало при действии ПМП.
Таким образом, опыты на полностью изолированной нервной цепочке речного рака показали, что нервная ткань, лишенная гуморальной связи с остальным организмом, может реагировать на ПМП. Реакция чаще выражалась в торможении спонтанной электрической активности нейронов. Подобные результаты получили американские исследователи, влияя ПМП на изолированный подглоточный ганглий тараканов.
Воздействуя непосредственно на основное звено регуляторных процессов, мы сможем управлять деятельностью всего организма. Важность непосредственного действия раздражителя на ЦНС неоспорима; речь может идти о распространенности этого явления, о его нормальном или патологическом характере. Говоря о распространенности, мы должны иметь в виду как организмы с различным уровнем биологической организации, так и факторы различной физической природы. Известно, что одноклеточные воспринимают любой раздражитель непосредственно своей одной клеткой. Только после значительного периода развития многоклеточные организмы приобретают органы чувств.
Следовательно, интересующая нас форма взаимодействия организма со средой является старым и единственным способом взаимодействия на низших этапах эволюционного развития. В какой-то мере каждая живая клетка сложного организма, несмотря на высокую степень специализации, сохранила способность непосредственно реагировать на отдельные факторы среды. И, конечно, в ряду клеток различных систем нервная клетка в этом отношении занимает одно из первых мест.
Проведенные эксперименты показывают, что исследуемые ПМП вызывают сложные неспецифические реакции, которые выявляются на уровне организма, ЦНС и нервных клеточных образований. В отличие от раздражителей, для восприятия которых имеются специализированные рецепторы, ПМП, обладая проникающим действием, может одновременно влиять на различные ткани организма. Однако из-за разной чувствительности тканей и органов, а также из-за включения репарационных процессов общая реакция организма меняется на разных этапах воздействия ПМП. Но на всех этапах реакции в ней принимает участие ЦНС. Нас интересовала начальная непосредственная реакция ЦНС на ПМП, так как влияние на этот центральный регуляторный орган является основным звеном в цепи ответов организма.
Другие авторы изучали длительное воздействие ЭМП. Так, киевские гигиенисты исследовали ЭЭГ сензомоторной и зрительной областей коры больших полушарий кроликов при воздействии поля УВЧ. Животные были разделены на 7 равных групп, одна из которых была контрольной, а 6 других подвергали 60-дневному воздействию (10—12 ч ежедневно) поля УВЧ напряженностью 0,05; 0,2; 0,5; 1,0; 3,0 и 6,0 В/м. Обнаружили, что действенными оказались поля УВЧ трех наибольших напряженностей. Начиная со второго дня и достигая вершины через 14 дней после начала воздействия, в ЭЭГ трех указанных групп кроликов увеличивался в 2—3 раза индекс альфа-ритма. Через 45 и 60 дней воздействия наблюдали увеличение индекса дельта-ритма. Характер ЭЭГ-реакции при длительном хроническом воздействии поля УВЧ и наличие пороговой напряженности поля УВЧ (между 0,5 и 1,0 В/м) показывают сходство в реакциях мозга на различные ЭМП.
Было отмечено, что, кроме основной реакции, возникала реакция на выключение (РНВ). Впервые о РНВ после выключения ПМП мы заговорили, когда изучали ЭЭГ-реакцию кроликов при воздействии ПМП на голову животного. В опытах длительность воздействия равнялась 60 с, а интенсивность достигала 45,0 мТл. Кроликам было дано 1113 воздействий ПМП и было зарегистрировано 360 РНВ, прочность которых составила 32,3%. Распределение латентных периодов РНВ по ЭЭГ показателям кроликов приведено на рис. 10, где за 100% взято общее число РНВ.
На том же рисунке показано распределение латентных периодов РНВ по сенсорному показателю для 5 испытуемых, которым давали 139 воздействий на руку ПМП интенсивностью 20,0 мТл с экспозицией 60 с. Хотя воздействие на человека было по индукции примерно в 2 раза ниже, чем воздействие на кролика, это обстоятельство нельзя считать существенным, так как, согласно нашим прежним данным, ПМП интенсивностью от 10 до 90 мТл вызывает сходные сенсорные реакции у человека.
Несмотря на разницу в интенсивности ПМП и в способе воздействия (прямое — на мозг и рефлекторное — на руку), в регистрируемых реакциях (сенсорная реакция и ЭЭГ-реакция) и в объектах исследования (человек и кролик) представленные кривые распределения поразительно похожи друг на друга. Кроме того, совпадение касается и частоты возникновения реакции на выключение (у кроликов она зарегистрирована в 32,3% случаев, а у людей — в 31,6%).
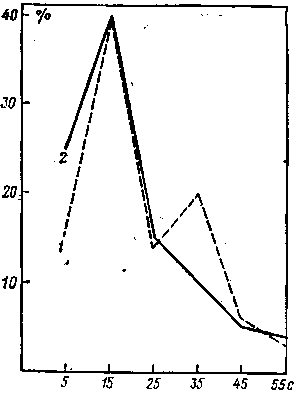
Рис. 10. Распределение величины латентных периодов ЭЭГ-реакций на выключение электромагнита у кроликов (1) и сенсорных реакций человека (2)
Можно предполагать, что организм позвоночных обладает общей медленной системой реагирования, которая может подключаться как при центральном, так и при периферическом воздействии.
Советский исследователь Н. А. Аладжалова, перечисляя свойства этой системы, указывает, что она может не реагировать на кратковременные стимулы, функционирует длительное время после прекращения воздействия и может менять уровень деятельности быстродействующей системы. Опыты показали, что реакции на ЭМП отличаются большим латентным периодом, длительным последействием и корригирующим влиянием. Иными словами, свойства реакции ЦНС на ЭМП совпадают со свойствами медленной системы реагирования. Неожиданным оказалось то обстоятельство, что электрические реакции на ЭМП нейронально изолированных структур мозга происходили чаще, скорее и интенсивнее в сравнении с интактным мозгом. Это можно объяснить снятием тормозящего действия со стороны рецепторов и нижележащих отделов ЦНС, а также неспецифическим повышением чувствительности нейронально изолированных мозговых структур.
Только ли в целостном мозге возникает реакция на выключение после прекращения воздействия различных ЭМП? При ответе на этот вопрос следует учитывать, что для получения яркой реакции на выключение необходима определенная интенсивность и длительность ЭМП.
В лаборатории украинского физиолога П. Г. Богача изолированный ганглий моллюска Планорбис помещали на 6 ч между полюсами электромагнита, создавая в разных сериях опытов ПМП 18,0, 60,0; 120,0 или 180,0 мТл. Отмечали снижение мембранного потенциала во всех случаях. Если на протяжении первых 4 ч процесс деполяризации при индукции ПМП 18,0 и 60,0 мТл проходил медленнее, то через 6 ч наблюдали одинаковое снижение мембранного потенциала во всех опытных сериях (в среднем на 40—47%) в сравнении с контролем. После прекращения воздействия намечалась тенденция к восстановлению мембранного потенциала нервных клеток, но на протяжении 3 ч его величина не достигала контрольного уровня. После воздействия ПМП 180,0 мТл наблюдали что-то похожее на описанную выше реакцию на выключение.
Использование электрографического метода для изучения этой системы совместно с фармакологическими воздействиями является перспективной областью исследования также межцентральных отношений в ЦНС при действии ЭМП. Однако при формулировке этой проблемы нельзя забывать, что реакция голодного мозга на ЭМП возникает одновременно во многих районах, а последовательность вовлечения тех или иных областей вероятнее всего определяется не параметрами воздействующего фактора, а состоянием реагирующего субстрата.
Эксперименты с изоляцией различных участков головного мозга при сохранении кровообращения показали, что деафферентированные участки ЦНС реагируют на ЭМП скорее, интенсивнее и чаще, чем интактный мозг. Необходимо в будущем проверить чувствительность и реактивность различных изолированных участков ЦНС млекопитающих к ЭМП.
Общий вывод из опытов с изолированным мозгом млекопитающих говорит о том, что система, включающая нейроны, глиальные образования и кровеносные сосуды, может реагировать на ЭМП, и в этой связи резонно проследить изменения при действии ЭМП гематоэнцефалического барьера, в который входят указанные структурные элементы мозга. Гематоэнцефалический барьер является частным случаем гистогематических барьеров, свойственных различным системам организма. Детальное изучение этой проблемы было начато академиком Л. С. Штерн в 20-х годах нашего столетия.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 1 МОЗГ
Глава 1 МОЗГ 1. ОБЩИЕ СВЕДЕНИЯ Традиционно со времён французского физиолога Биша (начало XIX в.) нервную систему разделяют на соматическую и вегетативную, в каждую из которых входят структуры головного и спинного мозга, называемые центральной нервной системой (ЦНС), а также
Глава 6. Как обмануть свой мозг?
Глава 6. Как обмануть свой мозг? Более 90 процентов всех решений мозг принимает самостоятельно. Наш разум не участвует в координации работы сердца или желудка, мы не задумываемся, сколько мышц следует задействовать, чтобы взять в руки яблоко, и не всегда способны
6.2. Полярность морфогенетических полей[141]
6.2. Полярность морфогенетических полей[141] Большинство биологических морфических единиц поляризовано по крайней мере в одном направлении. Их морфогенетические поля, содержащие поляризованные виртуальные формы, будут автоматически принимать подходящие ориентации, если
6.3. Размеры морфогенетических полей
6.3. Размеры морфогенетических полей Размеры индивидуальных атомных или молекулярных морфических единиц более или менее постоянны; это относится также к размерам кристаллических решеток, хотя они повторяются неограниченное число раз, образуя кристаллы различных
1. О природе М-полей
1. О природе М-полей Итак, согласно рассматриваемой гипотезе, фактор, определяющий образование форм на всех уровнях организации материи, — это морфогенетические поля. Утверждается, что это поля нового типа, не известного физике; соответственно, они не могут быть измерены
Глава 8 ИСКУСНЫЙ МОЗГ: УНИВЕРСАЛЬНЫЕ ЗАКОНЫ
Глава 8 ИСКУСНЫЙ МОЗГ: УНИВЕРСАЛЬНЫЕ ЗАКОНЫ Искусство это исполнение нашего желания найти самих себя среди явлений внешнего мира. Ричард Вагнер Прежде чем продвигаться дальше к следующим семи законам, я хотел бы разъяснить, что я понимаю под «универсальностью». То, что
Джордж Беркли Теория зрения, или зрительного языка, показывающая непосредственное присутствие и провидение божества; защищенная и объясненная. В ответ анонимному автору[18]
Джордж Беркли Теория зрения, или зрительного языка, показывающая непосредственное присутствие и провидение божества; защищенная и объясненная. В ответ анонимному автору[18] <…> 6. То, что атеистические принципы пустили глубокие корни и распространяются дальше, чем
Глава 7. Специфическое динамическое действие пищи
Глава 7. Специфическое динамическое действие пищи 7.1. Вводные замечания У большинства организмов можно выделить состояние голода, при котором используются эндогенные пищевые ресурсы (эндотрофия), и состояние сытости после потребления экзогенных пищевых веществ
Глава 7. Специфическое динамическое действие пищи
Глава 7. Специфическое динамическое действие пищи 7.1. Вводные замечания У большинства организмов можно выделить состояние голода, при котором используются эндогенные пищевые ресурсы (эндотрофия), и состояние сытости после потребления экзогенных пищевых веществ
Использование рыбами электрических полей
Использование рыбами электрических полей Электрическая локацияЕще Дарвину было известно, что некоторые рыбы имеют небольшие электрические органы. Разряды, излучаемые ими, настолько слабы, что казались естествоиспытателям совершенно бесполезными. Эти органы считались
Глава 4. Терапевтическое действие
Глава 4. Терапевтическое действие Для медицинских целей ЭМП важны не только при гигиенической оценке фактора или при использовании их в диагностике, но и для физиотерапии. Если ЭМП радиочастотного диапазона (поля УВЧ и СВЧ) в нашем веке широко используются врачами и
Глава 5. Экологическое значение геофизических электромагнитных полей
Глава 5. Экологическое значение геофизических электромагнитных полей Медицинская сторона экологического значения естественных ЭМП прежде всего находит выражение в корреляции между изменениями ЭМП и обострением различных заболеваний, хотя здоровые люди тоже
Глава 1 Мозг: мужской или женский?
Глава 1 Мозг: мужской или женский? Чуть более шестидесяти лет назад Симона де Бовуар в своей книге «Второй пол» написала: «Человек не рождается женщиной, а становится ею». Высказывание де Бовуар превратилось в универсальный девиз феминисток и дизайнеров моды. Вероятно,