Глава 8. Нейрон или глия?
Глава 8. Нейрон или глия?
Гемато-энцефалический барьер представляет собой сложную анатомическую, физиологическую и биохимическую систему, определяющую скорость проникновения отдельных веществ в мозг. На рис. 11 приведена схема сосудо-глио-нейронного комплекса, из которой следует, что важную барьерную функцию выполняет астроцит.
Хотя обсуждение вопросов влияния ЭМП на гемато-энцефалический барьер началось с регистрации морфологических изменений глиальных элементов не позже 1966 г. в работах М. М. Александровской, остро подчеркивалась его важная роль в реакциях ЦНС на ЭМП в работе А. Фрая в 1974 г., а на международном симпозиуме в 1977 г. эта проблема уже в качестве одной из важных обсуждалась за круглым столом.
Эксперименты электромагнитобиолога А. Фрая предназначены были определить влияние ноля СВЧ на мозг крыс. Оказалось, что импульсное поле СВЧ было более эффективным, чем непрерывное, судя по флюоресценции срезов мозга в ультрафиолетовом свете после введения в вену 0,15 мл 4%-ного раствора флюоресцина натрия. Картина флюоресценции наводит на мысль, что сосудистое сплетение желудочков мозга участвует в реакциях ЦНС на ЭМП, но этот эффект не всегда ограничивается только желудочками, но наблюдается также и в других областях мозга, особенно в промежуточном мозге. Поскольку облучение животных было общим, нельзя с уверенностью сказать, что эффект, наблюдаемый в мозге, прямой. Он мог быть результатом рефлекторного механизма. Сосудистое сплетение мозга явилось для советского биофизика С. Е. Бреслера и соавторов [1978] объектом исследования влияния ПМП на проницаемость биологических мембран.
Изменения гистогематического барьера и микроциркуляции наблюдали советские гигиенисты у лиц, работавших в условиях воздействия неблагоприятных факторов производственной среды (химическая интоксикация, вибрация, ЭМП радиочастот), сходные изменения свидетельствуют о неспецифическом характере реакций не указанные факторы среды.
В работах американского морфолога Э. Альберта, проведенных на крысах и хомячках с использованием электронно-микроскопической методики, показаны изменения нейроглии и микроциркуляции в мозге, связанные с повышением проницаемости гемато-энцефалического барьера, после воздействия поля СВЧ 2450 МГц 10 мВт/см2.
Описанные Э. Альбертом нарушения ультраструктуры головного мозга млекопитающих совпадают с данными советских исследователей, которые изучали действие ПМП на мозг. Грузинские морфологи электронно-микроскопическим методом исследовали изменения, возникающие в средней супрасильвиевой извилине коры больших полушарий головного мозга кошки после 1-часового воздействия ПМП индукцией 60 мТл. Изменения наблюдали не во всех нервных и глиальных клетках и не во всех синапсах. В нейронах разных слоев коры отмечали сходные реакции: появление инвагинаций ядерной оболочки, расширение цистерн и каналец эндоплазматической сети, уменьшение числа рибосом, набухание митохондрий, гипертрофию комплекса Гольджи и увеличение количества лизосом. Таким образом, под влиянием ПМП изменяются почти все структурные элементы тела нейрона. Причем изменения со стороны ядерной оболочки, митохондрий и комплекса Гольджи свидетельствуют о повышении метаболических процессов, а состояние эндоплазматической сети и рибосом говорит о подавлении белкового синтеза.
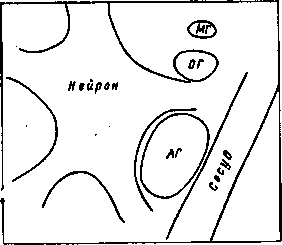
Рис. 11. Схема нейро-глио-сосудистого комплекса, являющегося структурной основой мозговой ткани
МГ — микроглин, ОГ — олигодендроглия, АГ — астроцитарная глия.
Наиболее заметные изменения возникали в ультраструктуре синапсов, особенно со стороны пресинаптических терминалей. Отмечали набухание митохондрий и появление крупных полиморфных вакуолей. Такие вакуоли достигали особенно больших размеров в постсинаптических областях: дендритах и шипиках.
В аксодендритных синапсах возникала значительная вариабельность размеров синаптических пузырьков, их перераспределение в бутоне и уменьшение общего числа. Встречались терминали, в которых вообще не было синаптических пузырьков. Эти картины свидетельствуют о значительном нарушении синаптической передачи после воздействия ПМП.
Недавно возникла гипотеза о возможном влиянии ЭМП на микротрубочки и в связи с этим на аксональный транспорт. Это предположение связано не только с фактами, свидетельствующими об изменении функций НС под влиянием ЭМП, но и с сообщениями о возникновении при действии ЭМП катаракты в хрусталике глаза, нарушении ретины, снижении сперматогенеза и ненормальном развитии куколок насекомых. Во всех перечисленных процессах важную роль играют микротрубочки.
Если в опытах в пробирке обсуждаемая гипотеза получила подтверждение при электронно-микроскопическом исследовании мозга крыс (исчезали микротрубочки в дендритах нейронов гипоталамуса), то в экспериментах с изолированной нервной тканью (ткань головного мозга или вагусного нерва кроликов-альбиносов) воздействие импульсного ЭМП 3,1 ГГц 240—900 Вт/м2 не вызвало эффекта.
Как указывалось в предыдущей главе, при регистрации электрической реакции головного мозга на различные ЭМП исследователи отмечали появление медленных высокоамплитудных колебаний биопотенциалов. В последнее время отводят важную роль в формировании именно медленных компонентов ЭЭГ глиальным элементам.
В опытах, проведенных нами совместно с М. М. Александровской методами световой микроскопии, реакция нейроглии и нейронов сензомоторной области коры больших полушарий животных, побывавших в ПМП 20,0—30,0 мТл, определялась морфологическими методами.
Уже через 3 мин после начала действия МП на голову кролика обнаружено увеличение числа окрашенных астроцитов в поле зрения микроскопа с 9,0±0,5 (контроль) до 16,7±0,4 и увеличение числа клеток микроглии с 11,5±0,4 (контроль) до 19,7±0,4. Отмечалось также увеличение числа клеток олигодендроглии. Нейроны при тех же условиях оставались сохранными, с хорошо выраженным ядром и ядрышком.
Через 1 ч после начала воздействия ПМП у кроликов отмечали более резкое увеличение (до 23,0±0,5) числа окрашенных клеток астроцитарной глии с увеличением тел и отростков. Число клеток других видов глии также увеличивалось. Нейроны оставались интактными.
Через 10—12 ч после начала воздействия у кроликов и кошек число клеток глии в сравнении с нормой оставалось большим. Нейроны претерпевали обратимые изменения в виде набухания и гиперхроматоза.
Через 60—70 ч после начала воздействия ПМП в головном мозге у кроликов, кошек и крыс наблюдали продуктивно-дистрофические нарушения нейроглии с набуханием олигодендроцитов и появлением дренажных клеток. Дистрофические нарушения захватывали и нервные клетки. Морфологически определялась картина гипоксической энцефалопатии с дистрофическими изменениями как астроцитов, так и микроглии.
Нужно заметить, что реакцию со стороны глиальных элементов в головном мозге при действии МП отмечали многие исследователи.
Сотрудник Куйбышевского медицинского института С. А. Батурина изучала особенности гипоталамической нейросекреции крыс после действия ПМП малых интенсивностей — от 1 до 10 мТл. Длительность воздействия продолжалась несколько минут. Материал взят непосредственно после сеанса ПМП через 15, 30 мин, 1, 2 ч, 1, 3, 5, 7, 10, 12, 15, 20 сут. Об интенсивности нейросекреции судили по количеству нейросекрета в клетках и отростках, размерам клеток, ядер и ядрышек, соотношению нейросекреторных клеток, находящихся в различных фазах нейросекреторного цикла. Полученные количественные данные обработаны статистически и составлены вариационные гистограммы. В работе также использовали метод гистоавторадиографии с введением тимидина-Н3.
Однократное 5-минутное воздействие ПМП приводило к быстрому выводу нейросекрета из нейрогипофиза: через 5—15 мин количество нейросекрета быстро снижалось. Выведение нейросекрета из нейрогипофиза, являющегося депо для нейрогормонов, происходило в течение 1 ч.
Через 1—2 ч после первою сеанса отмечался выход нейросекрета из нейроцитов супраоптических ядер, а также значительное усиление нейросекреции. В стадию синтеза переходила большая часть клеток — 48,8% (в контроле — 11,4%). Наряду с этим отмечалось значительное количество нейросекреторных гранул в отростках нервных клеток, где образуются довольно крупные расширения в виде капель диаметром до 10 мкм и более. Возрастал объем нейросекреторных клеток.
В паравентрикулярном ядре большая часть клеток переходила в стадию синтеза и накопления нейросекрета. Активное выведение нейросекрета отмечалось черев 1 сут после сеанса ПМП.
Первое воздействие ПМП приводило к тому, что большая часть клеток ядер начинала функционировать в одинаковой фазе. Под влиянием ПМП в ядрах гипоталамуса резко снижалось количество клеток, находящихся в состоянии покоя. Стадии покоя и накопления кратковременны, поэтому на препаратах большая часть клеток находилась или в состоянии синтеза, или в состоянии выведения. После действия ПМП в 10 мТ отмечался одновременный выброс нейросекрета из ядер гипоталамуса через 1—2 ч. В связи с этим нейросекрет неравномерно поступал в нейрогипофиз: в стадии выведения его много, а в стадии синтеза он почти полностью отсутствует. Этим могут быть объяснены циклические изменения, отмечаемые другими авторами в функциональной активности периферических эндокринных органов под влиянием ПМП.
Повышенная нейросекреторная активность гипоталамических ядер сохранялась в течение 10—12 сут.
Увеличение количества сеансов ПМП не вызывало дальнейшего усиления нейросекреторной активности. Стимулирующим действием на нейросекрецию обладало первое воздействие, а последующие сеансы оказывали поддерживающее действие. Повышение количества сеансов ПМП (в данном случае до 10) приводило к постепенному снижению эффекта стимуляции нейросекреции. Это свидетельствует о возможности адаптации организма к действию ПМП.
Под влиянием ПМП усиливается функция всех долей гипофиза, о чем косвенно свидетельствует повышение индекса меченых Т-Н3-ядер клеток аденогипофиза и нейрогипофиза.
Ни в одном из изученных случаев повреждающего действия ПМП на нейроциты не отмечено, что свидетельствует о высоком функциональном напряжении гипоталамуса.
В этой работе показано изменение под влиянием ПМП нейросекреторной активности крупноклеточных ядер гипоталамуса, в которых происходит синтез гормонов вазопрессина и окситоцина, обладающих прессорным и антидиуретическим действием.
Суммируя гистологические данные, мы составили табл. 3, из которой следует, что глия первой вовлекается в реакцию, первой среди равно реагирующих на МП элементов нейро-глио-сосудистого комплекса. Правда, роль капилляров в этой реакции изучена еще недостаточно (о чем свидетельствует знак вопроса в таблице), особенно в начальные периоды воздействия МП. Можно надеяться, что в скором времени заполнится и этот пробел в наших знаниях.
Участие глиальных образований в реакциях мозга на МП подтвердили недавно исследования [Думбадзе и др., 1980], проводимые в лаборатории известного физиолога члена-корреспондента АН СССР А. И. Ройтбака, который наиболее активно в нашей стране изучает роль глии в деятельности нервной системы. Суть упомянутых исследований заключается в том, что в вызванном электрическом ответе коры больших полушарий головного мозга при ее прямом электрическом раздражении во время действия ПМП отмечали ослабление медленного отрицательного потенциала. Такой эффект объясняют деполяризующим действием ПМП на глиальные клетки.
Знаменательно, что физиологические исследования проводятся параллельно с электронно-микроскопическими, которые показали, что в олигодендроцитах отмечается повышение электронно-оптической плотности ядра и цитоплазмы. У отдельных дендроцитов набухали тела и отростки; в них наблюдали уменьшение отдельных органоидов. Изредка попадались шиповидные выросты астроцитов, в которых имелись синаптические пузырьки. Скорее всего, ПМП приводило к распаду мембран астроцитарного отростка.
Таблица 3. Параметры морфологических реакций нейро-сосудистого комплекса центральной нервной системы на магнитные поля свыше 20 мТл в зависимости от длительности экспозиции
Экспозиция Нейрон Глия Сосуд Длительность последействия Аккумуляция Десятки секунд — — — — — Минуты — + ? Десятки минут — Десятки минут + + + Сутки + Часы + + + Недели + Сутки + + + Месяцы +Из приведенного описания следует, что в зависимости от целевой установки исследователя одни и те же морфологические изменения в головном мозге после воздействия ЭМП трактуются по-разному, исходя из ориентации в объяснении тех или иных интегральных функций ЦНС, проявляющихся в поведении целостного организма.
Впервые электронно-микроскопические изучения реакций мозга на МП начал Ю. М. Ирьянов в г. Ижевске. Он использовал длительные (до 75 ч) воздействия ПМП индукцией 2; 20; 200 и 400 мТл, а также ПеМП 50 Гц с индукцией 100 мТл. Исследовалась не только непосредственная реакция, но и репарационные процессы, возникающие через разные сроки после прекращения магнитного воздействия. Опыты проводились на мышах, так как эти животные становятся основным объектом магнитобиологических исследований и возможность сравнения данных различных исследователей значительно увеличивается. Изучали ультрамикроскопические изменения в нейронах и мантийных клетках гассерова ганглия, а также в седалищном нерве.
Наблюдаемый эффект зависел от индукции МП (2,0 мТл эффекта не вызывали), от характера, МП (переменное более эффективно, чем постоянное) и от длительности воздействий. Магнитобиологический эффект не ограничивался только временем воздействия МП, но длился еще долгое время (несколько суток) после прекращения воздействия. Эти выводы согласуются с многочисленными магнитобиологическими данными, полученными другими методами исследования. Однако оригинальность этой работы заключается в том, что в ней впервые показана высокая чувствительность к МП митохондрий, эндоплазматического ретикулума других органоидов нервной клетки.
Гипотеза о первичном действии МП на проницаемость биологических мембран сейчас наиболее распространена. Ю. М. Ирьянов [1971] также склоняется к этому мнению, основываясь на своих достаточно убедительных электронно-микроскопических данных. Однако он идет дальше указанного общего заключения, доказывая наибольшую чувствительность к МП мембран митохондрий и эндо-плазматического ретикулума в сравнении с другими мембранами нервной клетки. Более того, чувствительность к МП мембран этих органоидов зависит и от их локализации. К примеру, в теле нейрона изменения были более выраженными, чем в нервном волокне.
Высокая реактивность нейронов связывается с характером липопротеидных связей. Ю. М. Ирьянов не расшифровывает возможную структуру этих связей, в то время как его научный руководитель К. К. Сергеев, автор липопротеидной гипотезы нейроплазмы, указывает на возможность влияния внешних МП на электромагнитные силы, создающие липопротеидные и нуклеопротеидные комплексные соединения.
Если полученные в 60-х годах электронно-микроскопические данные Ю. М. Ирьянова можно рассматривать как эпизодические (сейчас эти исследования в Ижевске не проводятся) достижения одиночки, то 70-е годы характеризуются включением в русло нейромагнитобиологических исследований нескольких гистологических школ [Сперанский и др., 1976].
В этой связи прежде всего следует указать на электронно-микроскопические работы [Эрназаров, 1979], проводимые в Ташкентском медицинском институте под руководством М. С. Абдуллаходжаевой. В этих работах основное внимание уделяется зависимости эффекта от параметров воздействующего МП и от региональных особенностей участков головного мозга крыс (кора больших полушарий и гиппокамп). Исследователи концентрируются на детализации внутриклеточных изменений в центральном нейроне.
Представители грузинской нейроморфологической школы Т. И. Кикнадзе и Н. Л. Лазриев подчеркивают роль глии в реакциях головного мозга кошки на ПМП, хотя также тщательно описывают структурные изменения в нервной клетке. Эти работы отличаются комплексным морфологическим и электрофизиологическим подходом к наблюдаемым реакциям.
Среди зарубежных исследователей уже упомянутый Э. Альберт, ставший недавно президентом Биоэлектромагнитного общества, уделяет много внимания структурным изменениям головного мозга после воздействия на животных полем СВЧ.
При обсуждении сложного и важного вопроса о специфичности биологического действия МП Ю. И. Ирьянов приходит к выводу о специфическом действии МП на структуру митохондрий. Основанием для такого вывода послужили немногочисленные литературные сведения об угнетении дыхания и окислительного фосфорилирования в МП. Однако другие данные показывают, что характер измерения процессов дыхания в МП носит фазный характер, а также зависит от интенсивности МП и особенностей биообъекта. Так что на субклеточном уровне мы еще не можем четко обнаружить специфичность в действии МП.
Анализ изменений рибосом и ядрышка в МП приводит к заключению о снижении интенсивности синтеза белка в клетке. Вероятно, здесь уместно было охарактеризовать и степень изменения синтеза РНК.
Заканчивая эту часть книги, мы должны отметить, что анализ реакций клеток ЦНС на ЭМП подтвердил основные выводы, полученные при исследовании на системном и организменном уровнях. Эти реакции могут возникать при непосредственном действии ЭМП на структуры нервной ткани. Они отличаются небольшой прочностью, значительным латентным периодом и длительным последействием. Применение гистологического метода исследований дало дополнительные сведения о характере изучаемых реакций, многие особенности которых можно объяснить вовлечением глиальных элементов ЦНС.
Следует заметить, что, изучая влияние ПМП и ПеМП индукцией от 16,0 до 32,0 мТл, американские исследователи не могли обнаружить у кроликов «нейропатологии», хотя воздействовали МП по 3—4 ч в день на протяжении 15—20 сут. Они справедливо отмечают, что нейроглия может давать подобную реакцию на воспаление (энцефалит), которое часто встречается у кроликов. Для дальнейшего анализа глиальной гипотезы, характеризующей реакцию ЦНС на ЭМП, необходимо стандартизировать объект исследования (лучше не кролики), методы окраски (они не совпадают) и способы обработки (имеются количественные методы). Напомним, что совокупность данных, которые нам известны, чаще свидетельствует об участии нейроглии в реакциях НС на ЭМП.
Косвенные данные показывают, что под влиянием ЭМП претерпевают значительные изменения окислительные процессы. Но на этом пути мы уже должны выходить за пределы физиологического анализа и приниматься за биохимические исследования.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ГЛАВА XV.
ГЛАВА XV. Обработка земли под яровые хлеба.Обработку земли под яровые хлеба я начинаю тотчас после уборки озими . Только при соблюдении этого условия можно рассчитывать на самый обильный урожай.Поля, поросшие сорными травами и покрытые густым жнивьем, я вспахиваю
Глава I
Глава I Восемь лет назад я написал небольшую книгу «Аксиомы биологии»[1], в конце которой высказал предположение, что возможно создание общей теории эволюции последовательно реплицирующихся систем. Завершил я книгу словами: «Под эту категорию попадают не только объекты
2. НЕЙРОН. ЕГО СТРОЕНИЕ И ФУНКЦИИ
2. НЕЙРОН. ЕГО СТРОЕНИЕ И ФУНКЦИИ Мозг человека состоит из 10 12 нервных клеток. Обычная нервная клетка получает информацию от сотен и тысяч других клеток и передаёт сотням и тысячам, а количество соединений в головном мозге превышает 10 14 - 10 15 . Открытые более 150 лет тому назад
3.1. Парадигма реактивности: нейрон, как и индивид, отвечает на стимул
3.1. Парадигма реактивности: нейрон, как и индивид, отвечает на стимул Как мы уже отмечали, с позиций парадигмы реактивности поведение индивида представляет собой реакцию на стимул. В основе реакции лежит проведение возбуждения по рефлекторной дуге: от рецепторов через
3.2. Парадигма активности: нейрон, как и индивид, достигает «результат», получая необходимые метаболиты из своей «микросреды»
3.2. Парадигма активности: нейрон, как и индивид, достигает «результат», получая необходимые метаболиты из своей «микросреды» Упомянутые ранее этапы приобретают смысл в том случае, если принять, что процесс, обеспечивающий переход от прек постсинаптическим образованиям,
ГЛИЯ – МОРФОЛОГИЯ И ФУНКЦИЯ
ГЛИЯ – МОРФОЛОГИЯ И ФУНКЦИЯ Головной мозг человека состоит из сотен миллиардов клеток, причем нервные клетки (нейроны) не составляют большинство. Большая часть объема нервной ткани (до 9/10 в некоторых областях мозга) занята клетками глии (от греч. склеивать). Дело в том, что
НЕЙРОН
НЕЙРОН Нейрон является главной клеткой центральной нервной системы. Формы нейронов чрезвычайно многообразны, но основные части неизменны у всех типов нейронов. Нейрон состоит из следующих частей: сомы (тела) и многочисленных разветвленных отростков. У каждого нейрона
Глава 3
Глава 3 Описание метода лечебной кинологии и результатов исследования В предыдущих главах описывались теоретические аспекты метода лечебной кинологии. Постараемся схематически описать практический аспект методологии. Как уже отмечалось, данный метод находится на
Глава 2. ДНК
Глава 2. ДНК На стене паба “Орел” в Кембридже висит синяя мемориальная доска, установленная в 2003 году в честь пятидесятилетия одного случая, когда разговоры в этом пабе приняли не совсем обычный оборот. Во время обеда 28 февраля 1953 года два завсегдатая “Орла”, Джеймс
Глава III. Мир РНК-ДНК
Глава III. Мир РНК-ДНК 3.1. Миры до РНК и мир РНК Многие исследователи полагают, что первым клеточным миром был мир РНК (Ferris, 1999; Hoenigsberg, 2003). Однако по причинам, рассмотренным выше, более правдоподобна версия, согласно которой в ранних клетках функционировали информационные
Глава 1. Вид
Глава 1. Вид ТЕМЫ• История эволюционных идей• Современное эволюционное учение• Происхождение жизни на Земле• Происхождение человекаВ настоящее время на нашей планете обитает несколько миллионов видов живых организмов, каждый из которых по своему уникален. Каким
Глава 10. Уши{10}
Глава 10. Уши{10} Того, кто заглянет поглубже в ухо, чтобы увидеть, как устроен наш орган слуха, ждет разочарование. Самые интересные структуры этого аппарата скрыты глубоко внутри черепа, за костяной стенкой. Добраться до этих структур можно только вскрыв череп, удалив мозг,