Глава 9. Мембраны и биохимия
Глава 9. Мембраны и биохимия
Электронный микроскоп показал, что биохимические реакции в живой клетке протекают с активным участием мембранных процессов. Это заключение относится и к нервной, и к глиальной клетке, и к внутриклеточным органеллам.
Следует признать, что биохимические изменения в мозге, возникающие при воздействии ЭМП, изучены еще недостаточно. Так, 10-минутное воздействие ПМП 50,0 мТ на крыс приводило к увеличению в ткани головного мозга содержания аммиака на 560%, глутаминовой кислоты — на 62,9% и аспарагиновой кислоты — на 41,2%. В то же время отмечали снижение содержания глутамина на 71,3%. Знаменательно, что содержание гамма-аминомасляной кислоты, которую связывают с тормозным процессом НС, увеличивалось при этом на 60%.
В опытах на крысах ростовские исследователи изучали тканевое дыхание, окислительное фосфорилирование, анаэробный и аэробный гликолиз гипоталамуса с прилегающими к нему участками таламуса при воздействии ПеМП частотой 50 Гц и индукцией 200 мТл. При воздействии ПеМП на интактных крыс в гипоталамо-таламическом отделе мозга происходило значительное усиление тканевого дыхания и некоторое усиление фосфорилирования. ПеМП увеличивало в мозге гликолиз, особенно аэробный. Оказалось, что непосредственно после воздействия ПеМП на голову крыс происходит повышение уровня холинэстеразы в крови до 32%, а спустя 6 ч содержание холинэстеразы возвращается к исходному.
При изучении результатов воздействий ЭМП на целостный организм или локально на голову остается неясным вопрос, происходят ли наблюдаемые изменения за счет процессов, протекающих в самой нервной ткани, или они осуществляются за счет других механизмов. Поэтому представляет большой интерес изучение влияния ЭМП на деятельность изолированного мозга или одиночной изолированной переживающей нервной клетки. Нейроны являются образованиями, хорошо изолированными от электрических и магнитных влияний межклеточной жидкостью и другими клеточными элементами. Однако, так как средняя ионная концентрация межклеточной жидкости поддерживается постоянно механизмами активного транспорта и гемато-энцефалическим барьером, то каждый раз, когда возникает потенциал действия, происходят локальные изменения концентраций ионов вследствие обмена ионами между нейронами и окружающей средой. В то же самое время внеклеточная жидкость перемешивается и движется под действием эпиндимных клеток, которые снабжены ресничками. Тогда можно заключить, что локальные концентрации ионов, в частности ионов кальция и магния, могут быть изменены ЭМП и соответственно могут значительно влиять на синаптические передачи.
Ионы кальция важны для регуляции мембранного потенциала покоя и для синаптического возбуждения. Оказывается, кальций распределен в мозговой ткани дифференциально. Ионам кальция отводится важная роль в реализации воздействия ЭМП на физико-химические (Л. Д. Кисловский) и биологические системы (А. Г. Карташев). Более высокие концентрации его находят в нейронных мембранах, в синаптических районах, в нейроглиальной цитоплазме и в нейрональных органеллах, но в нейрональной эктоплазме его относительно мало. Глюкопротеины мембранной поверхности обнаруживают сильное средство к катионам, и особенно к ионам кальция и водорода. Таким образом, оказалось, что кальций играет существенную роль в классических механизмах нервного возбуждения. Помимо этого, вероятно его участие в более слабых событиях на поверхности мембраны, включая редкое взаимодействие с рассеянными молекулами гормонов и восприятие слабых электрических градиентов.
В лаборатории американского нейрофизиолога У. Эйди было показано, что слабые синусоидальные ЭП изменяли выход кальция из свежей изолированной ткани мозга цыпленка или кошки. Вслед за инкубацией (30 мин) с радиоактивным кальцием (45Са2+) каждая проба помещалась в свежий физиологический раствор и подвергалась 20-минутному воздействию полей 1, 6, 16, 32 или 75 Гц с электрическим градиентом 5, 10, 56 и 100 В/м. Выход 45Са2+ в раствор измеряли затем в 0,2 миллилитровой навеске и сравнивали с выходом из контрольных проб. Все пробы находились при температуре 36° С. Воздействие ЭП приводило к уменьшению освобождения кальция. Максимум снижения наблюдали при 6 и 16 Гц (от 15 до 20%). Пороги были около 10 и 56 В/м для мозга цыпленка и кошки соответственно (рис. 12).
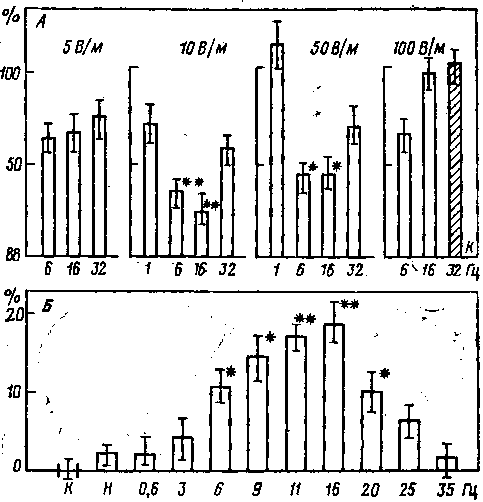
Рис. 12. Выход Са из ткани мозга цыпленка после воздействия ЭП (А) в поля СВЧ (Б) [Эйди, 1977]
Абсцисса — частота (Гц); ордината — величина реакции (%). Звездочками отмечены достоверные отличия от контроля. Цифры сверху — напряженность ЭП (В/м), к — контроль
Ранее описано резкое увеличение выхода кальция из изолированной ткани мозга цыпленка при действии модулированных радиочастотных ЭМП. Эти исследования показали, что ответ наблюдался в узкой полосе низкочастотных модуляций (6—25 Гц) и, что характерно, отсутствовал при действии немодулированной несущей волны (147 МГц).
Исследования различных участков изолированной коры больших полушарий мозга кошки показали значительное снижение выхода кальция при действии ЭП частотой 6 и 16 Гц с градиентом 56 В/м. Незначительные тенденции к снижению выхода наблюдали также в поле данной напряженности при частотах 1,32 и 75 Гц. Никаких существенных эффектов не наблюдали не только в поле 10 В/м с частотой 6,16 и 32 Гц, но и в поле 100 В/м с частотой 6 и 16 Гц. Следовательно, мы имеем «резонансную» кривую с максимальной чувствительностью ткани в области 6 и 16 Гц и с амплитудным окном между 56 и 100 В/м.
Не было обнаружено разницы между различными корковыми участками, которые относились к преимущественно сенсорным, двигательным или ассоциативным областям.
Данные по выходу кальция при действии низкочастотных ЭМП на мозговую ткань цыпленка и кошки могут быть объединены в некоторой последовательности. Начальные изменения происходят на продольной оси мембраны, включая, возможно, макромолекулярные конформационные изменения, зависящие от изменений связей кальция с полианионными гликопротеинами. Слабое изменение в одной точке может вызвать макромолекулярные конформационные изменения на значительном расстоянии вдоль мембраны.
Для низкочастотных и радиочастотных ЭМП обнаружено существование минимальной чувствительности к полям «биологической» частоты, но результаты четко показывают, что способ взаимодействия зависит от амплитуды поля. Возможное основание для этой амплитудной избирательности может лежать в способе связывания кальция с цепями биополимеров.
Если, как предполагали раньше, кооперативная динамика связывания кальция на возбудимых мембранах определяет предельный цикл поведения, то субстратно-кальциевое равновесие будет колебаться с фиксированной частотой и амплитудой.
Таким образом, слабое нарушение любого параметра может легко разрушить электрохимическое равновесие. Однако имеющиеся результаты не могут указать на дипольный момент или на другую молекулярную чувствительность в таком узком частотном диапазоне. Существованием амплитудного окна для этих прямых взаимодействий внешних низкочастотных ЭП с тканью мозга можно объяснить описанные ранее поведенческие и нейрофизиологические изменения.
Спонтанные циркадные ритмы у человека и птиц, живущих в экранированных камерах, становились короче при наложении электрического поля 10 Гц прямоугольной волны с градиентом 2,5 В/м в воздухе. Оценки интервалов времени (5 с) у обезьян ускоряются на 10% при наличии 7 Гц, 10 В/м синусоидального электрического поля. Дальнейшие исследования расширили эти представления, показав максимальную чувствительность в области 7 Гц с прогрессивным увеличением порогов к 45 и 75 Гц.
При исследовании распределения РНК в рецепторе рака микрофотометрическим методом выявились изменения, особенно резко выраженные в зонах отхождения дендритов и аксона. К 30-й минуте действия ПМП в области отхождения дендритов наблюдается уменьшение плотности, а в области отхождения аксона, наоборот, увеличение плотности РНК. После снятия ПМП через 30 мин наступает некоторое восстановление исходной картины. Фотометрирование тела нейрона по поперечному диаметру выявило увеличение плотности РНК к 30-й мин действия ПМП в перинуклеарной области, сглаживающееся после прекращения действия ПМП.
Результаты этих опытов говорят о том, что реакции нервной ткани на ПМП могут иметь в основе изменения свойств генерации импульсов нейронами. Нарушение функциональной деятельности нейронов, по-видимому, является отражением изменения характера их внутриклеточного метаболизма, которое наблюдали в данном случае по показателям динамики РНК, имевшим место при действии ПМП.
Влияние на метаболизм НС других диапазонов ЭМП интенсивно исследуют харьковские гигиенисты под руководством Г. И. Евтушенко. Были отмечены изменения разных сторон обмена НС при воздействии ИМП и при воздействии поля УВЧ. В последнем случае нужно отметить большое количество обследованных животных (500 крыс), широкий набор биохимических методик, изучение раздельного воздействия электрической и магнитной компонент ЭМП, вариации в способах воздействия (однократный, многократный и хронический) и в его интенсивности. Интересно, что при воздействии ЭП наиболее чувствительным показателем оказалось содержание лактата в мозгу и крови, а для МП — содержание преформированного аммиака и повышение содержания амидного азота в белках мозга крыс. Эти результаты дают дополнительный материал для выяснения специфичности воздействия магнитных и электрических полей. В будущем желательно сопоставить эти оригинальные биохимические данные с результатами физиологических и морфологических исследований.
Можно надеяться, что именно на этом уровне изучения функций НС будет выявлена специфичность действия различных диапазонов ЭМП, как она начинает выявляться при изучении разных компонент ЭМП.
Поскольку показано, что внешние ЭМП изменяют состояние мозга, возникают вопросы, касающиеся роли внутренних мозговых низкочастотных ЭМП.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
§ 8. Заряды мембраны нервных клеток
§ 8. Заряды мембраны нервных клеток Однако основные свойства нервной системы обусловлены способностью быстро реагировать на изменение ситуации внутри или вне организма. Скоростные процессы не могут осуществляться по медленным гуморальным законам, они происходят по
2. Строение клеточной стенки и цитоплазматической мембраны
2. Строение клеточной стенки и цитоплазматической мембраны Клеточная стенка – упругое ригидное образование толщиной 150–200 ангстрем. Выполняет следующие функции:1) защитную, осуществление фагоцитоза;2) регуляцию осмотического давления;3) рецепторную;4) принимает
Глава 9. Биологические мембраны
Глава 9. Биологические мембраны Клетка представляет биологическую систему, основу которой составляют мембранные структуры, отделяющие клетку от внешней среды, формирующие ее отсеки (компартменты), а также обеспечивающие поступление и удаление метаболитов, восприятие и
Глава 12. Биохимия гормонов
Глава 12. Биохимия гормонов Гормоны (от греческого hormaino – побуждаю) – это биологически активные вещества, которые выделяются эндокринными клетками в кровь или лимфу и регулируют в клетках-мишенях биохимические и физиологические процессы.В настоящее время предложено
Глава 14. Биохимия питания
Глава 14. Биохимия питания Наука о пище и питании называется нутрициологией (от греч. нутрицио - питание). Нутрициология или наука о питании – это наука о пище, пищевых веществах и других компонентах, содержащихся в продуктах питания, их взаимодействии, роли в поддержании
Глава 22. Метаболизм холестерола. Биохимия атеросклероза
Глава 22. Метаболизм холестерола. Биохимия атеросклероза Холестерол – стероид, характерный только для животных организмов. Основное место его образования в организме человека – печень, где синтезируется 50% холестерола, в тонком кишечнике его образуется 15–20%, остальное
Биохимия атеросклероза
Биохимия атеросклероза Атеросклероз – это патология, характеризующаяся появлением атерогенных бляшек на внутренней поверхности сосудистой стенки. Одна из основных причин развития такой патологии – нарушение баланса между поступлением холестерола с пищей, его
Глава 28. Биохимия печени
Глава 28. Биохимия печени Печень занимает центральное место в обмене веществ и выполняет многообразные функции:1. Гомеостатическая - регулирует содержание в крови веществ, поступающих в организм с пищей, что обеспечивает постоянство внутренней среды организма.2.
Глава 30. Биохимия крови
Глава 30. Биохимия крови Кровь – жидкая подвижная ткань, перемещающаяся по сосудам. Выполняет роль транспортного и коммуникативного средства, интегрирующего обмен веществ в различных органах и тканях в единую систему. Общая характеристика Общий объем крови у взрослого
Глава 31. Биохимия почек
Глава 31. Биохимия почек Почка – парный орган, основной структурной единицей которого является нефрон. Благодаря хорошему кровоснабжению почки находятся в постоянном взаимодействии с другими тканями и органами и способны влиять на состояние внутренней среды всего
Глава 33. Биохимия мышечной ткани
Глава 33. Биохимия мышечной ткани Подвижность является характерным свойством всех форм жизни - расхождение хромосом в митотическом аппарате клеток, воздушно-винтовые движения жгутиков бактерий, крыльев птиц, точные движения человеческой руки, мощная работа мышц ног. Все
Биохимия мышечного утомления
Биохимия мышечного утомления Утомление – состояние организма, возникающее вследствие длительной мышечной нагрузки и характеризующееся временным снижением работоспособности.Центральная роль в развитии утомления принадлежит нервной системе. В состоянии утомления в
Глава 34. Биохимия соединительной ткани
Глава 34. Биохимия соединительной ткани Соединительная ткань составляет около половины от сухой массы тела. Все разновидности соединительной ткани, несмотря на их морфологические различия, построены по общим принципам:1. Содержит мало клеток в сравнении с другими
Глава 2 Биохимия экономики
Глава 2 Биохимия экономики Также любят они соседа и жмутся к нему, ибо им необходимо тепло. Ницше Ф. Так говорил Заратустра Как правило, люди отвечают добром на добро и испытывают непроизвольную симпатию к тем, кто относится к ним хорошо. Это естественное чувство симпатии
Статус, самооценка и биохимия
Статус, самооценка и биохимия В глубине поведенческих параллелей между человеком и человекообразными обезьянами лежат параллели биохимические. В стаях обезьян-верветок у доминирующих самцов обнаруживается более высокий уровень нейротрансмиттера серотонина, чем у
ЧАСТЬ ЧЕТВЕРТАЯ. БИОХИМИЯ ГОЛОДАНИЯ (Под редакцией Академика АМН СССР А. А. ПОКРОВСКОГО)
ЧАСТЬ ЧЕТВЕРТАЯ. БИОХИМИЯ ГОЛОДАНИЯ (Под редакцией Академика АМН СССР А. А. ПОКРОВСКОГО) Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва) В течение последних лет в нашей стране и