Глава 10. Собственные электромагнитные поля мозга
Глава 10. Собственные электромагнитные поля мозга
О биопотенциалах мозга написаны тысячи научных и популярных статей и книг за те полвека, что прошли со времени начала регистрации электроэнцефалограммы человека немецким физиологом Г. Бергером.
В те же годы советским академиком А. В. Леонтовичем была выдвинута гипотеза о нейроне как аппарате переменного тока. На основании нейрогистологических исследований он пришел к выводу, что у нейрона имеются катушки и емкости, иными словами все то, что надо для контура Томпсона, предназначенного для генерации ЭМП. Поэтому он решил, что передача информации с нейрона на нейрон может происходить двумя путями: химическим и электрическим.
Советские академики П. П. Лазарев и В. М. Бехтерев считали принципиально возможным зарегистрировать ЭМП биологического происхождения, а известный английский электрофизиолог. Е. Д. Эдриан скептически относился к этой идее.
Ученик А. В. Леонтовича Б. В. Краюхин сообщил в 1945 г., что ему удалось зарегистрировать магнитное поле возбужденного нерва лягушки с помощью специальной катушки. Но эти результаты не были подтверждены. Попытки намагнитить иглу, помещенную рядом с мышцей или нервом животного, не увенчались успехом.
Много копий поломано при обсуждении роли электрических полей в целостной деятельности мозга. Поскольку показано, что внешние электрические поля изменяют состояние мозга, возникают вопросы, касающиеся роли внутренних мозговых низкочастотных ЭМП.
Нелишне напомнить, что вся громадная сегодняшняя электротехника зародилась от биоэлектричества рыб и лягушек. За два века своего интенсивного развития электротехника успела позабыть свои истоки и черпает новые идеи для прогресса из собственного арсенала. В этой колыбели возникла и радиотехника. В наши дни мы наблюдаем обратный процесс: влияние радиотехнических идей на объяснение механизма самых разных биологических процессов.
В 40-х годах нашего века при использовании метода перерезок удалось получить факты, которые позволили выдвинуть теорию об электрическом (эфаптическом) взаимодействии нервных клеток коры больших полушарий головного мозга. Отмечается роль эфаптической передачи в деятельности рядом расположенных нервных волокон. Кроме того, эта проблема обсуждается и при изучении перехода нервного возбуждения на эффекторный орган, в частности на мышцу.
При изучении условий перехода на нейронально изолированную полоску коры головного мозга высокоамплитудных судорожных разрядов, вызванных отравлением стрихнином интактной части полушария, констатировали, что такой переход возможен, если генерируются биопотенциалы не ниже 500 мкВ.
Нужно отметить, что нейрофизиологи, изучающие эфаптическое распространение нервного возбуждения, обычно не обсуждают вопросы возможного моделирования такого распространения с помощью воздействия искусственных ЭМП. Эфаптическое воздействие для них значит прежде всего несинаптическое. На доказательстве различий в этих воздействиях они и сосредоточиваются, и о параметрах ЭМП, осуществляющих эфаптическое воздействие, мы имеем очень смутные представления.
Пока только на примере рыб убедительно показано биологически значимое существование разных по силе живых генераторов ЭМП. Здесь мы ограничимся обсуждением электрической составляющей ЭМП. Вероятно, впервые его замерил английский физик Джон Деви в 1832 г. у одной из электрических рыб, хотя этих экзотических животных с усердием изучали такие всемирно известные естествоиспытатели прошлого века, как А. Гумбольдт, М. Дюбуа-Раймон, М. Фарадей и др.
Начало современного этапа в изучении биоэлектрических полей рыб связывают с именем английского эколога X. Лиссмана, который в 1958 г., используя современную электронную технику, показал, что некоторые рыбы используют свои ЭМП для локации и связи. Сейчас такие работы интенсивно развиваются.
Еще раньше морфологи, изучавшие строение рыб, отметили у некоторых их представителей наличие специальных органов, которые генерируют ЭМП, и специальных органов чувств (электрорецепторы), которые их воспринимают.
Предполагается, что из 20 тыс. видов рыб, плавающих сейчас в водоемах Земли, не менее 300 видов способны создавать ЭМП. По напряженности создаваемых ими ЭМП рыб можно разделить на три группы: электрических, создающих ЭМП свыше 20 В в целях нападения и обороны; слабоэлектрических, создающих ЭМП до 17 В в целях локации и связи, и неэлектрических (до нескольких милливольт). Генерируемые рыбами ЭМП располагаются в полосе частот до 2 кГц. Они создаются или специальными электрическими органами или неспециализированными нервно-мышечными структурами.
Биологические ЭМП ничем не отличаются по своим свойствам от ЭМП, созданных техническими генераторами, т. е. имеют электрическую и магнитную составляющие, ослабляются с расстоянием и т. п.
Наверняка у биологических ЭМП имеются те же биотропные параметры, как и у искусственных ЭМП. Во всяком случае уже известно, что рыбы более чувствительны к переменным полям в сравнении с постоянными, что реакция зависит от частоты, формы и длительности импульсов.
Чувствительность рыб одной группы располагается в пределах 0,01—0,1 мВ/см, а другой — в пределах 10—100 мВ/см, т. е. на несколько порядков ниже. В первом случае речь может идти о своеобразном электрическом восприятии внешней среды. Предполагают, что центры, регулирующие электрорецепторы, располагаются в продолговатом мозге и мозжечке. Следовательно, для некоторых животных электромагнитный океан становится зримым.
Главные направления в использовании биологических ЭМП рыбами заключаются в обороне и нападении, в пространственной ориентации и в сигнализации, связанной с общением между биообъектами. Интересно, что группа рыб на ЭМП реагировала лучше, чем одиночная особь. Возникают интересные вопросы об эволюционной исключительности рыб при использовании ЭМП. Можно надеяться, что некоторые стороны использования ЭМП рыбами со временем будут обнаружены и у других водных или наземных животных.
Впервые в «сухопутных» условиях электрические поля удалось зарегистрировать у земноводных животных. Но это случилось не потому, что у земноводных ЭМП сильнее, чем у растений, птиц или млекопитающих, а по той причине, что лягушка, явилась излюбленным и удобным экспериментальным объектом для нейрофизиологов. Как раз изолированный седалищный нерв гигантской лягушки был объектом исследований американцев Барра и Маура, которым удалось на расстоянии до 12 мм отмечать возникновение ЭМП в момент возбуждения нерва.
Более детальные работы по регистрации электрических ауральных (от греческого слова «аура» — воздух) полей у различных сухопутных животных и у человека проводятся с 1966 г. в Ленинградском государственном университете под руководством профессора П. П. Гуляева. Были подтверждены данные о существовании ЭП у возбужденного изолированного нерва лягушки. Впервые обнаружили ЭМП у возбужденного изолированного рецептора растяжения речного рака.
Нужно отметить, что сам факт синхронизации в активности большого числа нервных клеток привел к предположению об их объединении не только синаптическим (контактным), но и эфаптическим (через ЭП) путем.
Как ни интересны ЭП, сегодня наиболее интригующими являются магнитные поля мозга. В этой области, как в прошлом в области эмоций, другом — соперником мозга выступает сердце.
Известно, что биотоки нашего организма, возникающие за счет биохимических реакций, играют большую роль в физиологических процессах (особенно в деятельности сердца и мозга!), объединяя функционирование отдельных элементов указанных органов. Уже теоретически можно было предсказать, что биотоки должны порождать биомагнитные поля, но практически зарегистрировать такие поля было очень трудно.
Цену усилий можно оценить хотя бы по тому факту, что, как мы теперь знаем, магнитное поле сердца человека составляет примерно миллионную часть земного магнитного поля, а магнитное поле мозга еще в 100 раз слабее.
Перед исследователями биомагнитных полей, как и перед героями древних былин, вставали три главных препятствия, которые казались неодолимыми. Во-первых, нужно было создать датчик для обнаружения очень слабых магнитных полей. Во-вторых, нужна была уникальная усилительная аппаратура, необходимая для регистрации воспринятых слабых сигналов. И, в-третьих, нужна была экранировка от земного магнитного поля и технических электромагнитных помех.
Начальное решение первой проблемы можно назвать лобовой атакой. Из школьного курса физики известно, что магнитное поле наводит электрический ток в витке провода, а величина этого тока зависит от интенсивности поля и от числа витков, используемых для регистрации магнитного поля. Поскольку интенсивность биомагнитного поля очень мала, чувствительность датчика можно было повысить за счет увеличения числа витков. Исходя из такого простого рассуждения, советские и американские исследователи независимо друг от друга сумели в начале 60-х годов нашего века впервые зарегистрировать магнитное поле сердца человека, используя катушки, число витков в которых достигало миллиона. Но это был предел возможностей датчиков такого рода: воспринимать более слабые, а также стационарные магнитные поля, возникающие, например, при повреждении сердца — эти датчики не могли. Нужны были другие идеи.
Помощь пришла с неожиданной стороны. Работая в области криогенной техники со сверхпроводящими материалами, английский ученый Б. Д. Джозефсон (он получил Нобелевскую премию за свое открытие) обнаружил, что между двумя сверхпроводниками, разделенными тонким слоем диэлектрика, возникает в этих условиях электрический ток, параметры которого в значительной степени зависят от окружающего магнитного поля. Этот принцип позволял регистрировать очень слабые как переменные, так и постоянные магнитные поля, что выгодно отличало его от датчиков в виде катушек.
На основе эффекта Джозефсона были созданы СКВИДы (сверхпроводниковые квантовомеханические интерференционные датчики), чувствительность которых может достигать величины 10-14 Тл. Магнитометры, работающие на базе СКВИДа, получили широкое распространение в геофизических и космических исследованиях, в экспериментальной физике и, что нас в наибольшей степени интересует, в медико-биологических исследованиях.
Регистрация магнитных полей человека с помощью СКВИДов была начата в середине 60-х годов в США, а позже в орбиту этих исследований включались и другие страны (Канада, Франция, Япония, Финляндия, Италия и др.). В августе 1978 г. в Гренобле (Франция) состоялось Международное рабочее совещание по биомагнетизму, на котором в основном обсуждались проблемы, связанные с регистрацией магнитных полей здорового и больного сердца и мозга человека.
Нужно заметить, что магнитография человека на основе СКВИДов остается еще относительно дорогой процедурой, а к недостаткам этих датчиков следует отнести их сложность и необходимость регулярной заливки дефицитным жидким гелием.
Более перспективными по отмеченным параметрам могут быть квантовые магнитометры с оптической накачкой (МОН), практическое применение которых успешно развивается в нашей стране. Ленинградским исследователям во главе с Е. Б. Александровым в 1978 г. была присуждена Государственная премия за цикл исследований МОН.
В Институте земного магнетизма, ионосферы и распространения радиоволн АН СССР (ИЗМИРАН) А. Н. Козловым и С. Е. Синельниковой несколько лет назад был создан МОН, в датчике которого использовались пары щелочного металла цезия. Созданный геофизиками прибор оказался пригодным и для медицинских целей. В этом случае уже не требуется дорогая и сложная криогенная техника, на основе которой работают СКВИДы, а чувствительность прибора достаточно высока [Ливанов и др., 1978].
Таким образом, имеются три пути преодоления трудностей при создании датчиков для восприятия биомагнитных полей: катушки с миллионами витков, СКВИДы и МОНы. Для преодоления второго препятствия — создания надежной усилительной техники — за прошедшие годы наметилось много путей, и мы их перечислять не будем, заметив только, что препятствие это преодолимо.
Когда я летом 1978 года посещал Хельсинкский технологический университет, финские коллеги показали мне запись МКГ эмбриона, находящегося еще в утробе матери. Таким образом, магнитография позволяет «разглядеть» отдельные внутренние органы, создающие биомагнитные поля, и оценить их работу.
Я уже перечислял органы, от которых записаны биомагнитные поля, но меня как нейрофизиолога прежде всего привлекает возможность регистрации магнитных полей мозга магнитоэнцефалограммы (МЭГ).
Впервые сообщение о регистрации МЭГ опубликовал американский исследователь Д. Коен в 1968 г., но лишь в последние годы научились надежно регистрировать не только фоновую МЭГ, но и вызванные магнитные поля мозга, возникающие при действии световой вспышки, звукового щелчка или электрического раздражения пальца руки испытуемого.
Детальное сравнение фоновых МЭГ и ЭЭГ при их одновременной регистрации у одного и того же испытуемого показало, что в состоянии бодрствования имеется строгое соответствие МЭГ и ЭЭГ в диапазоне альфа-ритма (8—14 Гц). Но при засыпании человека, что сопровождается обычно преобладанием медленной активности в электрической деятельности мозга, медленные волны могли появляться одновременно в ЭЭГ и МЭГ или только в ЭЭГ, или только в МЭГ. Подобные эпизодические рассогласования в показателях МЭГ и ЭЭГ были обнаружены у больных эпилепсией во время их бодрствования. Эти факты свидетельствуют о возможности получения новой информации о деятельности мозга с помощью метода МЭГ.
Еще большие различия между ЭЭГ и МЭГ выявились при сравнении вызванных ответов мозга. Магнитные ответы четко локализовались над проекционными областями соответствующих анализаторов в коре больших полушарий головного мозга человека, тогда как электрические ответы были более расплывчатыми. Магнитный ответ в большей степени зависел от интенсивности применяемого раздражителя, чем электрический ответ мозга.
Следовательно, некоторые тайны целостного мозга можно узнать с помощью чувствительного магнитометра, а совершенствование этого прибора позволит больше узнать и о деятельности отдельной нервной клетки. На пути совершенствования методов магнитометрии можно, судя по публикациям, встретить такие альянсы, как, например, специалиста невролога со специалистом по магнетизму геологических объектов. Задача у этих специалистов одинаковая — определить слабые магнитные поля.

Рис. 13. Запись магнитных полей сердца, мышц, глаза и мозга с помощью магнитометра оптической накачки
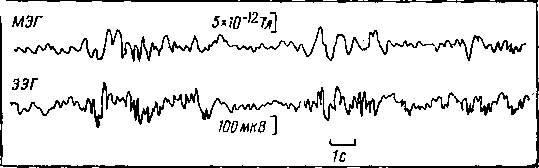
Рис. 14. Одновременная запись ЭЭГ и МЭГ человека с помощью СКВИДа [Коен, 1975]
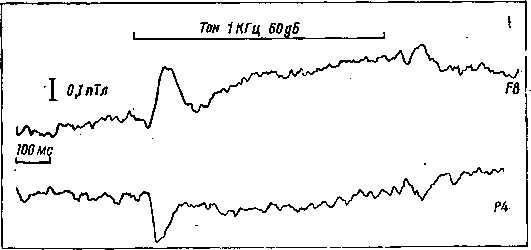
Рис. 15. Вызванный магнитный ответ на звуковое раздражение, записанный с разных участков (F8P4) мозга автора книги финскими исследователями А. Пенттииеном и К. Рейникайненом в магнитоэкранированной комнате в Финляндии
В наших условиях прохождение по коридору человека со вставными металлическими зубами могло сказаться на работе чувствительного магнитометра. А если сотрудник в соседней комнате брал в руки ножницы для вскрытия почтового конверта, то прибор «зашкаливал» на несколько минут.
По внешнему виду основная часть градиентометра напоминает бинокль или скорее микроскоп. Два датчика представляют собой параллельно расположенные два вакуумных стеклянных цилиндра, заполненных парами цезия. Два гибких световода доносят свет специальной лампы до цилиндров, а два экранированных провода относят от цилиндров электрические сигналы, порожденные в фотодетекторе на других концах цилиндров. Эти сигналы меняются при действии биомагнитных полей, а усилительная и регистрирующая аппаратура фиксирует такие изменения [Ливанов и др., 1978].
По своему назначению градиентометр также можно сравнить с микроскопом, поскольку он позволяет разглядеть малые магнитные поля, генерируемые различными органами человека.
На рис. 13 видно, что, располагая датчик возле сердца, мы можем записать магнитокардиограмму (МКГ). Помещая датчик около плеча, мы уже не видим магнитных полей сердца, так как они быстро уменьшаются с расстоянием, но можем зарегистрировать магнитные поля мышц (магнитомиограмму — ММГ), возникающие при сокращении бицепсов. Моргание глаз тоже не ускользнет от чуткого датчика прибора и отразится в магнитоокулограмме (МОГ). Наконец, когда датчик располагали около затылка, регистрировали магнитные поля мозга (магнитоэнцефалограмму — МЭГ). Можно видеть, что наибольшие магнитные поля создает сердце, а наименьшие — мозг. Поэтому МКГ исследована сегодня наиболее полно.
На рис. 14 можно видеть одновременную запись МЭГ и ЭЭГ у одного и того же испытуемого. Бросается в глаза похожесть двух процессов, если не обращать внимания на изменение фаз и выраженность отдельных волн. Отсюда следует, что источниками магнитных и электрических сигналов чаще всего являются одни и те же физиологические процессы. Только МЭГ могли регистрировать бесконтактным методом на расстоянии до 2 см от поверхности черепа испытуемого, а для регистрации ЭЭГ всегда необходимы контакты с кожной поверхностью пациента.
Неэмоциональные слова «бесконтактный метод» свидетельствуют о значительном преимуществе магнитографии в медицине в сравнении с электрографией. От контактов электродов с кожной поверхностью человека в значительной степени зависит качество получаемых записей, по которым ставят диагноз. Обеспечить одинаково хороший контакт часто бывает трудно (он зависит от индивидуальных свойств кожной поверхности), а иногда и вовсе невозможно (при кожных заболеваниях, при обширных ожогах и т. п.).
Кроме того, контакты с электроаппаратурой требуют повышенной заботы об электробезопасности процедуры, тогда как магнитография избавляет персонал от таких забот, упрощая процедуру записи до минимума.
Наконец, запись магнитограмм человека можно осуществлять намного быстрее, чем запись электрограмм, значительно увеличивая тем самым пропускную способность соответствующих кабинетов.
Во время моего недавнего пребывания в Лаборатории низких температур Хельсинкского технологического университета в магнитоэкранированной камере с помощью СКВИДа финские исследователи зарегистрировали вызванное магнитное поле на звук в двух областях мозга (рис. 15). Интересно, что вектор МП во фронтальной области был другого направления, чем вектор в височной области, чего нельзя было обнаружить при регистрации вызванного электрического потенциала на тот же звук.
Сегодняшнее прозаическое перечисление преимуществ бесконтактной регистрации магнитных полей мозга уместно сравнить с недавними сообщениями о «мозговом радио» и других фактах, относящихся к разряду таинственных явлений человеческой психики.
Всего лишь двадцать лет назад известный ленинградский физиолог профессор Л. Л. Васильев в своей книге «Таинственные явления человеческой психики» выделил главу «Существует ли „мозговое радио“?». Кроме случаев так называемой спонтанной телепатии, в этой главе описываются попытки зарегистрировать ЭМП вокруг головы человека.
Те же самые двадцатые годы нашего столетия, которые породили метод ЭЭГ человека и предположения об электрических разговорах нейрона с нейроном, дали основания для гипотезы о «мозговом радио». Это направление исследований связано с именем итальянского невролога Ф. Кацамалли. Он сообщал, что когда человек, находящийся в электрически экранированной камере, испытывал эмоциональное возбуждение, экспериментатор мог прослушивать звуки, усиленные радиоприемником. Иногда эти радиоволны мозга Ф. Кацамалли регистрировал графически, хотя относил их к метровому, дециметровому или сантиметровому диапазонам. Заметим, если волны мозга можно было слышать, значит, по крайней мере они модулировались звуковой частотой.
В Ленинградском институте мозга им. В. М. Бехтерева попытались повторить опыты Кацамалли, но не получили положительных результатов.
Немецкие физики Ф. Зауербрух и В. Шуман сообщили в 1928 г., что сумели зарегистрировать ЭМП мышц человека и животных с частотой 50 Гц. Академик П. П. Лазарев писал: «Всякое ощущение, всякий акт движения должны образовать волны, и голова человека должна излучать волны большой длины (до 30 000 км) в окружающую среду»[3]. Следовательно, одни исследователи говорили о низкочастотных ЭМП, излучаемых мозгом человека, а другие — о высокочастотном; одни регистрировали электрическое поле вокруг возбужденного нерва лягушки [Гуляев и др., 1977], другие — магнитное [Краюхин, 1945], а третьи ничего не могли зарегистрировать [Хведелидзе и др., 1965]. Последние авторы приводят расчеты, согласно которым магнитное поле возбужденного нерва должно иметь величину порядка 10-13—10-15 Тл, что находится за пределами чувствительности имеющейся аппаратуры.
Некоторые исследователи в качестве датчика «мозгового радио» предлагают использовать мозг другого человека, предполагая при этом, что передача идет в сантиметровом диапазоне или с помощью низкочастотных ЭМП [Путхофф, Тарг, 1976].
Даже краткое изложение тех фактов, которые посвящены «мозговому радио», показывает, что это таинственное явление психики может изучаться с помощью современных технических средств путем регистрации магнитоэнцефалограммы. МЭГ имеет только диагностическое значение и нам неизвестны сообщения о том, чтобы магнитные поля мозга одного человека как-то влияли на деятельность мозга другого человека.
Другое дело — влияние искусственных ЭМП на мозг. Здесь имеется немало «таинственных» аспектов, связанных со старением, с оживлением и с явлениями сна.
Возможно, впервые A. Л. Чижевский заметил еще в 1940 г., что ослабленное МП укорачивает жизнь крыс. К подобным выводам пришли американцы Ван Дейк и Халперн, но уже в опытах на мышах, результаты которых опубликованы в 1965 и 1966 гг. Ослабленные МП укорачивали также жизнь дрозофил.
Когда на сессии Американской ассоциации содействия науке в 1978 г. перечисляли 37 способов продления жизни, которые, скорее всего, будут использоваться к 2000 г., то забыли упомянуть о благоприятном воздействии МП на процессы старения. Между тем имеются сообщения, что МП определенной индукции могут увеличивать как среднюю, так и, что особенно привлекательно, видовую продолжительность жизни позвоночных животных и насекомых при общем воздействии. Особенно ярко в экспериментальных условиях выявляется увеличение срока жизни под влиянием МП в условиях некоторых неблагоприятных факторов (инфекция, радиация, гипоксия и т. д.).
Ростовские исследователи Л. X. Гаркави, Е. Б. Квакина и М. А. Уколова [1979] сообщают, что после воздействия в течение 2—3 месяцев ПеМП на голову пожилых крыс животные превращались в подвижных и бодрых. Редкая грубая шерсть сменялась мягкой и густой, глаза становились ярко розовыми, а кожа мягкой и эластичной. Омоложенные животные отличались от молодых только по размеру.
Объяснение этим поразительным результатам авторы находят в гипотезе преодоления хронического стресса, характеризующего старость, путем развития свойственной молодости реакции активации (предстрессовое состояние, сопровождающееся увеличением тимуса, определенным соотношением нейтрофилов и лимфоцитов в крови, умеренным повышением активности щитовидной железы и половых желез и преобладанием секреции минералкортикоидных гормонов коры надпочечников). Длительное поддержание реакции активации приводило у самок к восстановлению нормального полового цикла.
Существует элевационная теория старения (В. М. Дильман), которая исходит из того факта, что в старости повышается активность гипоталамуса. На протяжении всей книги мы не раз обращали внимание на выраженную реакцию гипоталамуса при воздействии ЭМП. Отмечался также корригирующий характер действия ЭМП на функции мозга. Эти же факты пригодились для объяснения омолаживающего эффекта МП.
В опытах на крысах было показано, что для поддержания молодости следует через определенный период повторять сеансы воздействия МП.
ЭМП могут влиять на мозг и сразу после смерти. По крайней мере такой вывод можно было сделать уже из названия доклада «Электрическая активность мозга в постлетальном периоде, вызванная облучением электромагнитным полем», представленного группой исследователей, возглавляемой В. В. Тяжеловым из Института биологической физики АН СССР. Доклад зачитан на Всесоюзной конференции «Действие физических факторов на живой организм», проходившей в Одессе в апреле 1978 г.
Напомню читателю, что латинский термин «постлетальный» переводится на русский язык как «после смерти». С другой стороны, известно, что сам факт смерти устанавливают по прекращению электрической активности мозга. Следовательно, влияя на мертвую (даже отрезанную) голову кролика, у которой, конечно, нет ЭЭГ, импульсно-модулированным полем СВЧ с плотностью потока мощностью около 5,0 мВт/см2, можно получить кратковременное (на 30—60 с) возникновение ЭЭГ. Такое «оживление» может возникнуть после минутного воздействия полем СВЧ и тем надежнее, чем короче был интервал между моментом прекращения ЭЭГ и моментом начала воздействия. Самый большой интервал, установленный экспериментаторами, достигал 3 ч. В дальнейшем речь должна идти и о других таинственных явлениях человеческой психики — о сне и сновидениях.
Сноподобное действие ЭМП отмечали многие исследователи. Сюда можно отнести повышение порогов на сенсорные и болевые раздражители, торможение условных рефлексов и, главное, преобладание в ЭЭГ тех картин, которые характеризуют стадию медленноволнового сна. Можно еще добавить, что центр сна обычно локализуют в гипоталамусе, который, по многочисленным сообщениям, наиболее чувствителен к ЭМП.
Химия медленноволнового сна также похожа на изменения в мозгу при действии ЭМП. Например, введение гамма-аминомасляной кислоты вызывает появление веретен и преобладание в ЭЭГ волн дельта-диапазона, а также сон по поведенческим показателям.
Подобные изменения вызывало введение в кровяное русло животного серотонина или его предшественников. Магнитобиологические работы показывают, что содержание в организме указанных выше веществ меняется при воздействии МП.
Как показал наш краткий обзор, проблемы, связанные с влиянием ЭМП на мозг, нередко относят к разряду «таинственных», констатируя наше незнание каких-то существенных сторон мозговой деятельности. Часто встречаются замечания о том, что магнитные поля обладают сомнительным биологическим действием, и пора разобраться в этом деле. Привкус таинственности, противоречивости, зыбкости все еще сопровождает исследования по электромагнитной биологии. Неспроста, считают некоторые, слово «магия» идет в предметном указателе сразу после слова «магнетизм», а понятия «гипнотизер» и «магнитезер» являются синонимами.
Таинственность пасует там, где все становится понятным и где ясен механизм явлений и возможно их повторение. Следовательно, настала пора поговорить о механизмах.
Начать эту главу следует с того, что понятие «механизм действия» является очень многоликим понятием, включающим как биологические, так и физико-химические и даже квантово-механические подходы. Например, для нейрофизиолога важным является обсуждение рефлекторного и непосредственного механизмов действия ЭМП на мозг. Автору этих строк много сил и времени понадобилось, чтобы доказать (вначале себе самому, а потом и другим) существование непосредственного действия ЭМП на ЦНС. Теперь можно утверждать, что с этой пространственной точки зрения в механизм биологического действия ЭМП на мозг включаются как рефлекторные, так и непосредственные процессы.
Таблица 4. Время появления различных реакций нервной системы на магнитные поля свыше 20 мТл
Время Виды реакций Секунды (1 — 3) Нет Десятки секунд Сенсорная, ЭЭГ, условно-рефлекторная Минуты Двигательная активность, изменения сенсорных процессов, электрическая активность, УР и морфологии Часы Дистрофические изменения Сутки Деструктивные измененияХарактеризуя механизм влияния ЭМП на мозг с временных позиций, следует отличать по крайней мере быструю систему реагирования (в случае радиозвука, вызванного потенциала на импульс поля СВЧ, магнитофосфена и других явлений) и медленную систему, проявляющуюся в случае изучения сенсорной индикации МП, ЭЭГ-реакций на ЭМП и т. д. Распределение временных параметров появления отдельных реакций ЦНС на МП отображено в табл. 4.
В этот анализ временной последовательности включения различных механизмов мозга и организма в реакции на ЭМП следует включить ответы на выключение ЭМП, последействие, репарационные изменения и т. д. Сейчас еще мало фактов, чтобы нарисовать правдоподобную картину развития перечисленных процессов. Остается пока констатировать сложность этой задачи и дожидаться накопления экспериментального материала и появления новых свежих идей для его обобщения.
Нервную систему часто разделяют на двигательную и вегетативную. Хотя обе они подвержены влиянию ЭМП, вегетативная нервная система выступает как более чувствительное образование. Об этом подробно пишет М. И. Яковлева в своей книге, которая называется «Физиологические механизмы действия электромагнитных полей» [Л., Медицина, 1973].
В вегетативную нервную систему входят парасимпатический (как полагают, более чувствительный к ЭМП) и симпатический отделы, которые совместно обеспечивают регуляцию деятельности внутренней системы организма. На эти регуляторные процессы и действуют ЭМП, изменяя чаще всего деятельность органа, вызванную какими-то другими причинами.
Конечно, физиологические механизмы действия ЭМП не ограничиваются вегетативной нервной системой. Можно раздельно рассматривать чувствительность к ЭМП отдельных участков рефлекторной дуги: рецептор — центральное звено — эффектор. И здесь центральное звено оказалось более уязвимым.
При электрическом раздражении седалищного нерва крысы (находящейся под наркозом) можно регистрировать как биоэлектрические процессы в иннервируемой мышце, так и биопотенциалы различных отделов головного мозга. Если крысу поместить на 10—20 мин в постоянное магнитное поле с индукцией 400 мТ, то в икроножной мышце нельзя было заметить изменений потенциала действия (рис. 16), в то же время вызванный потенциал на электрическую стимуляцию указанного нерва увеличивался по амплитуде и приобретал дополнительные колебания. Сходную картину наблюдали как в сенсомоторной области коры больших полушарий головного мозга, так и в гипоталамусе и в коре мозжечка. Подобные изменения вызванного потенциала наблюдали при повышении уровня бодрствования животного или при увеличении силы раздражения. Иными словами, МП указанных параметров как-то усиливало ответ мозга на электрическую стимуляцию нерва, тогда как на ответ мышцы то же поле влияния не оказывало.
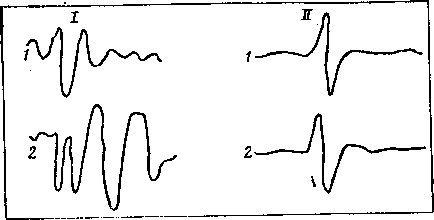
Рис. 16. Вызванный потенциал сенсомоторной коры больших полушарий мозга (I) крысы и потенциал действия икроножной мышцы (II) лягушки на электрическую стимуляцию седалищного нерва в фоне (1) и после 10—20-минутного воздействия постоянным магнитным нолем с индукцией 400 мТ (2)
Продолжая перечисление возможных механизмов действия ЭМП на мозг, следует обратить внимание на межцентральные отношения. Наиболее чувствительным к ЭМП многие исследователи считают гипоталамус, где сосредоточены высшие вегетативные центры (рис. 17). Но имеется мнение, что гиппокамп может брать на себя ведущую роль в организации реакции мозга на ЭМП. Конечно, важную роль в осуществлении этой реакции играет кора больших полушарий мозга и другие его ведущие отделы.
Иерархия межцентральных отношений в мозге при действии ЭМП меняется в зависимости от исходного функционального соотношения отдельных центров, а для детального анализа этого соотношения необходимо знать, что происходит на клеточном уровне.
Не разбирая подробно роль нейрона и глии в реакции мозга на ЭМП (чему посвящена отдельная глава), мы должны основное внимание уделить мембранным процессам, функция которых окончательно не выяснена.
Выдвигаемые гипотезы о молекулярных механизмах функционирования биологических мембран должны удовлетворять принципу реципрокности протекания в мембране сопряженных молекулярных процессов, что обусловливает совершенно необычные с позиций классических представлений конкурентные взаимоотношения поглощения Са и фосфорилирования АДФ, переноса электронов и активации АТФазы в митохондриях, а в плазматических мембранах приводит к реципрокным соотношениям электрогенного натриевого насоса и аккумуляции К+, активности холино- и адренорецепторов.
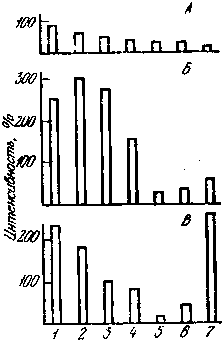
Рис. 17. Реакции разных отделов мозга кролика на ЭМП в норме (А), а также после введения кофеина (Б) или адреналина (В)
Ордината — интенсивность реакции (%); 1 — гипоталамус; 2 — сензомоторная кора; 3 — зрительная кора; 4 — специфические ядра таламуса; 5 — неспецифические ядра таламуса; 6 — гиппокамп; 7 — ретикулярная формация
Этому требованию удовлетворяет гипотеза М. А. Шишло, согласно которой в мембране митохондрий происходит переход разности электрических потенциалов в скачок концентрации ионов водорода на границе раздела двух сред, который используется для синтеза АТФ (рис. 18). Система включает два основных элемента, работающих на основе электронно-колебательных взаимодействий, которые в соответствии с пространственно-зарядовой асимметрией мембраны, определяемой разностью pH по обе стороны и гидрофобными взаимодействиями белков и липидов, должны иметь положительную кооперативную связь ?рН с виброном, участвующим в синтезе АТФ, и отрицательную кооперативную связь с вибронным возбуждением, осуществляющим выброс Н+ за пределы митохондрий и участвующим в транспорте кальция.
Из анализа модели вытекает важнейший вывод о том, что система, управляемая скачком концентрации протонов, не может быть системой с дипольными искажениями и должна обладать свойствами кристаллов, в которых параметром порядка служит деформация. Только в этом случае сброс протона по его электрохимическому градиенту может сопровождаться эффективной утилизацией энергии этого перехода в синтезе АТФ.
Если управление посредством изменения разности потенциалов вибронной неустойчивостью системы с дипольным искажением является общепризнанным и физически ясным процессом, то управление посредством ?рН требует пояснения и доказательств.
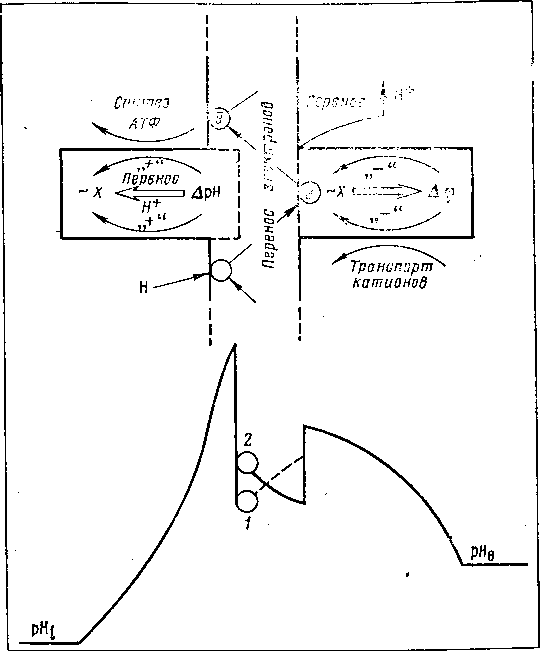
Рис. 18. Схема механизма биологического действия ЭМП
Оно становится возможным лишь в том случае, если деформационная система стационарно находится в искаженном состоянии в результате химических гидрофобных взаимодействий. Быстрое появление скачка протонов на границе липид—белок нейтрализует гидрофобный градиент системы с деформационным искажением, что приводит к ее вибронному возбуждению, и протоны стекают по градиенту, участвуя в синтезе АТФ. Такой механизм обеспечивает суммацию в акте синтеза АТФ энергии деформационного искажения с энергией, выделяемой в ходе сбрасывания Н+ по их электрохимическому градиенту. Ни одна равновесная искаженная ян-теллеровская система без механизма поддержания искаженной конфигурации не может обеспечить такой суммации двух вкладов при активации эндергонической химической реакции синтеза АТФ. Из этого вытекает фундаментальное значение гидрофобных взаимодействий в мембранах.
Исходя из изложенного, ограничение процессов переокисления липидов становится особо необходимым для нормального функционирования АТФ-синтетазы и любого другого механизма, построенного на использовании энергии деформационного искажения кооперативных электронно-колебательных систем.
Становится очевидным, что гипероксия может избирательно повреждать функционирование этих деформационных кооперативных систем ослаблением гидрофобных взаимодействий при усилении перекисного окисления липидов.
В рассматриваемой системе изменение пространственно-зарядовой асимметрии мембраны изменяет вклад деформационной и электрической составляющих в элементарный акт синтеза АТФ, но не изменяет стехиометрию: один квант вибронного возбуждения системы с деформационным искажением — один элементарный акт синтеза АТФ. Изменяется при этом лишь форма и высота потенциальных барьеров для туннелирования протона и время протекания элементарного акта синтеза АТФ.
Интересно, что изменение трансмембранной ?рН принципиально обращает пространственно-зарядово-временную симметрию системы с дипольными искажениями. Повышение pH закономерно приводит в такой системе к переключению от механизма поглощения К+ в обмен на Н+ к механизму электронного переноса К+, активируемого туннельным переносом электрона в данной структуре.
На этом мы закончим описание модели и покажем далее ее адекватность в описании эффектов ЭМП в митохондриях.
В рассматриваемой системе при воздействии ЭМП можно ожидать взаимодействия электрической составляющей с дипольной кооперативной системой мембран митохондрий. При этом ЭМП может инициировать ток деполяризации в сегнетоэлектрике, что должно активировать дыхательную цепь митохондрий и приводить в конечном итоге к диссипации поглощенной энергии в тепло. Система поддержания асимметрии мембраны за счет окислительно-восстановительного потенциала должна быть при этом активирована, и после выключения поля должен остаться «след» от компенсации деэнергизующего действия ЭМП.
Исследования показали, что в момент действия высокочастотного ЭМП происходит вибронное возбуждение дипольной кооперативной системы, осуществляющей сброс протона за пределы митохондрий. Поэтому после прекращения действия ЭМП можно ожидать появления неравновесного распределения зарядов на мембране (либо протонов, либо катионов), которое должно приходить к равновесию в ближайшем периоде последействия.
Как мы описывали ранее, ЭМП при действии на нервную систему может вызвать появление и развитие комплекса функциональных сдвигов. В основе этих сдвигов лежат разнообразные биофизические процессы, суть которых остается еще неизученной. Таких процессов одновременно происходит несколько, и мы не беремся здесь перечислять их все. Упомянем только, что МП могут индуцировать электрический ток (некоторые исследователи только этот механизм и признают), а поля СВЧ большой интенсивности могут вызывать нагревание (на чем основаны многие исследования в США). Воздействие ЭМП на возбудимые структуры можно связать с изменением калий-натриевого градиента в клетке за счет колебаний молекул воды, гидратирующих ионы и белковые молекулы, поверхностного слоя мембраны клетки.
Возможность влияния ЭМП на свойства водных растворов показана при техническом использовании так называемой магнитной обработки воды, в результате чего уменьшается осаждение на стенках котлов растворенных в ней солей, понижается смачиваемость поверхностей твердых тел, повышается скорость процессов сгущения суспензий, фильтрации, твердения цемента и т. д., а также изменяются ее биологические свойства.
ЭМП могут поляризовать боковые цепи белковой молекулы, вызывая разрывы водородных связей и изменяя зону гидратации молекул. Существует предположение, что сильные ПМП могут влиять на ориентацию макромолекул, в частности РНК и ДНК, и тем самым изменять биологические процессы. Увеличение активности ферментов трипсина и карбоксидисмутазы под влиянием ПМП позволяет предполагать, что химические изменения играют важную роль в первичных механизмах действия ЭМП. Поглощение переменных полей может происходить за счет ионной проводимости среды, релаксационных колебаний дипольных молекул и резонансных процессов. ЭМП, таким образом, нарушает структуру водных систем, изменяет активность ферментов и влияет на проницаемость биологических мембран.
Первичная реакция биологической системы на разные ЭМП заключается, вероятно, в возникновении поляризации. Это представление согласуется с теорией П. П. Лазарева [1935], по которой ЭМП низкой частоты вызывает в тканях периферические движения ионов. Накопление заряженных ионов на границах фаз может вызвать обратимое выпадение коллоидов. Возможно, этот механизм связан с неспецифическим действием переменных ЭМП, тогда как тепловое действие сопровождается необратимыми изменениями в клетке.
Независимо от характера взаимоотношений между биологическими тканями и энергией переменных ЭМП эффект обусловлен явлениями поглощения. При высоких частотах (3*1010 — 3*107 Гц) основным механизмом поглощения являются изменения ионной проводимости. При понижении частоты до 3*105 — 3*107 Гц наблюдается максимум поглощения за счет релаксационных колебаний белковых молекул. При дальнейшем понижении частоты может возникнуть максимум поглощения за счет релаксационных колебаний полярных макромолекулярных комплексов.
Можно предполагать, что уже на клеточном уровне действие разных в физическом отношении ЭМП выражается в неспецифической реакции поляризации мембраны. При обсуждении механизма влияния ПМП на нервную клетку выдвигалось предположение о действии этого фактора на переменно возбудимые зоны аксона и считалось, что конечный эффект можно объяснить умножением и суммацией очень малых биофизических явлений. Многочисленные данные показывают участие глиальных клеток в реакциях ЦНС на ЭМП. Скорее всего, на сегодняшнем этапе наших знаний следует говорить о влиянии ЭМП на глионейрональный комплекс, а это влияние реализуется в основном через изменение окислительных процессов.
Влияние ЭМП на возбудимые структуры сходно с действием анода постоянного тока. Оно не вызывало сокращения нервно-мышечного препарата, но изменяло хронаксию и снимало парабиоз. Сходным образом действовало поле УВЧ и ПМП.
Многие исследователи показывают, что ЭМП санти- и миллиметрового диапазонов волн способны вызвать как гипер-, так и деполяризацию наружной мембраны клеток. Вероятно, конечный эффект взаимодействия ЭМП с клеткой должен определяться типом клеток, их энергетическим обменом и его сопряженными связями с процессами активного транспорта и электрогенеза.
Важно, что как ПМП, так и ЭМП высоких частот способны вызывать в нервных и мышечных клетках однотипные генерализованные ответы в виде гипер- или деполяризации. Большинство исследователей относят эти изменения к влиянию ЭМП на процессы активного и пассивного транспорта в наружной мембране в связи с нарушением кооперативного связывания кальция на поверхности самой мембраны или в саркоплазматическом ретикулуме клетки. Важность кальциевого механизма действия ЭМП на нервную систему не вызывает сомнений.
Но несомненно перспективным является и привлечение представлений о едином механизме транспортных процессов на наружной клеточной мембране за счет сбрасывания протона по полю, формируемому за счет катионобменных свойств цитосола клетки и буферных свойств внешней среды. Тогда в трансмембранной разности потенциалов можно выделить две компоненты: стационарную, обусловленную свойствами внутриклеточного ионообменника (в том числе его емкостью) и значением pH внеклеточной среды, и метаболическую, обусловленную скоростью продукции протонов через АТФазные реакции и гликолитический путь окисления углеводов.
Исходя из этого при всех способах увеличения скорости продукции протонов в цитосоле клеток можно ожидать активацию электрогенного натриевого насоса, если последний не повреждается самим внешним возмущением. Тогда связь кальцевого обмена клетки с его процессами активного транспорта должна опосредоваться метаболическими реакциями клетки. Активация выброса протонов из митохондрий и усиление гликогенолиза под действием кальция являются известными феноменами. Тогда активацию электрогенного натриевого насоса можно рассматривать как закономерный процесс реакции клетки на ЭМП возмущения, сопровождающееся нарушением кооперативного связывания кальция и активацией гликолитических процессов.
Получается, что ответ в виде гиперполяризации нервной клетки может быть получен за счет магнитной составляющей ЭМП, имеющей высокое сродство к митохондриям. Подавление функции последних закономерно приводит к активации гликолитических процессов и создает предпосылки для активации электрогенного натриевого насоса. Реакция нейрона на электрическую составляющую ЭМП, скорее всего, начинается с поверхности клетки в гликокаликсе. В реальных условиях две компоненты взаимодействия ЭМП могут суммироваться.
Модель взаимодействия процессов активного транспорта в наружной мембране с метаболическими реакциями клетки позволяет подойти к интерпретации возникновения колебательного процесса в системе наружная мембрана—цитосол клетки в виде периодических изменений скорости активного транспорта натрия и калия в наружной мембране и сопряженных реакций метаболического котла клетки.
Описываемый механизм колебательного процесса может обусловливать медленные колебания мембранного потенциала глиальных клеток и приводить в ряде нейронов к возникновению периодической спайковой активности, сопряженной с медленными колебаниями потенциала покоя наружной мембраны. Кажется вероятным, что подобные связи электрогенеза с метаболическими реакциями могут лежать в основе наблюдаемого после действия ЭМП преобладания в нервной системе медленных биоэлектрических потенциалов.
Изложенные представления не противоречат возможности первичного деполяризующего действия внешнего ЭМП на клетку, когда фаза гиперполяризации будет уменьшена или даже полностью отсутствовать. Конечный ответ клетки может зависеть от прямого действия ЭМП соответствующих интенсивностей на механизм натриевого насоса. Последнее наблюдается при действии ЭМП миллиметрового диапазона и не имеет места при действии слабых ПМП. С другой стороны, характер ответа должен зависеть от присущих той или иной клетке метаболических характеристик, определяемых как генетически, так и их текущим состоянием. Среди генетически обусловленных характеристик можно выделить максимальную скорость продукции АТФ, максимальную скорость генерации протонов и восстановленных никотиновых коферментов, принимающих участие в регуляции активного транспорта потенциалобразующих ионов в наружной мембране.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ГЛАВА 11 Электрический ток и магнитные поля
ГЛАВА 11 Электрический ток и магнитные поля О том, что некоторые рыбы могут генерировать электрический ток, было известно еще древним грекам, хотя они и не знали, что то шоковое оцепенение, которое вызывали у рыбаков электрические скаты, было связано с действием
Глава 9. Физиология мозга
Глава 9. Физиология мозга Нет области науки более важной для человека, чем исследование его собственного мозга. Ф. Крик (1916–2004), английский генетик, лауреат Нобелевской премии 1962 г. Несмотря на значительный объем данных, полученных в исследованиях по эволюционной
ПРОБЛЕМЫ ПРИСПОСОБЛЕНИЯ: ЕСЛИ УМЕЛЫЕ ТРЮКИ ВЫЖИВАНИЯ БЬЮТ В СОБСТВЕННЫЕ ВОРОТА
ПРОБЛЕМЫ ПРИСПОСОБЛЕНИЯ: ЕСЛИ УМЕЛЫЕ ТРЮКИ ВЫЖИВАНИЯ БЬЮТ В СОБСТВЕННЫЕ ВОРОТА Питие доводит до врача: как тропические муравьи справляются с наводнениями Представьте себе: у вас есть дом на берегу реки, и вдруг река выходит из берегов. Что вы будете делать? Наверное,
Глава 1. Строение мозга
Глава 1. Строение мозга Мозг человека – самый сложный и загадочный орган тела. Для того чтобы его понять, человечеству пришлось потратить несколько столетий, но чем дальше мы продвигаемся в изучении, тем больше возникает новых вопросов. По большому счету, мозг
Сколько звезд имеют собственные названия?
Сколько звезд имеют собственные названия? Собственные названия имеют всего 275 ярких звезд, 80 процентов из них даны арабами. Часто это названия частей тела тех фигур, которые давали название (у арабов) всему созвездию. Например, Бетельгейзе – «плечо гиганта», Денебола –
МОИ СОБСТВЕННЫЕ ИССЛЕДОВАНИЯ
МОИ СОБСТВЕННЫЕ ИССЛЕДОВАНИЯ Я сам провел эксперименты двух типов. В экспериментах первого типа, которые были поставлены с несколькими группами в Европе и США, четыре человека выступали в качестве добровольцев-испытуемых и сидели в одном конце комнаты спиной к
Глава 4. Морфогенетические поля
Глава 4. Морфогенетические поля 4.1. Морфогенетические зародыши Морфогенез не происходит в вакууме. Он может начаться только с уже организованной системы, которая служит морфогенетическим зародышем. В процессе морфогенеза новая морфическая единица более высокого уровня
Глава 9. Движения и моторные поля
Глава 9. Движения и моторные поля 9.1. Введение В предыдущих главах обсуждалась роль формативной причинности в морфогенезе. В этой и двух последующих главах речь пойдет о роли формативной причинности в управлении движением.Некоторые движения растений и животных являются
9.6. Морфогенетические поля и моторные поля
9.6. Морфогенетические поля и моторные поля Несмотря на то что поля, контролирующие изменения формы специализированных двигательных структур животных, — это фактически морфогенетические поля, они вызывают, скорее, движения, нежели изменения формы. По этой причине
ГЛАВА IV. ОВОЩИ ЛЕСА И ПОЛЯ
ГЛАВА IV. ОВОЩИ ЛЕСА И ПОЛЯ Белый картофель североамериканских индейцев Среди бесчисленного множества растений, которые покрывают поверхность суши и водную поверхность земного шара, нет, быть может, ни одного, которое с большим правом заслуживало бы внимания добрых
Глава 8 Пол мозга
Глава 8 Пол мозга No woman, no cry. Боб Марли У женщин проблема одна, Я повторяю всегда От Камчатки до Аргентины. Эта проблема — мужчины. Огден Нэш/Курт Вайль (пер. О. Волковой) Сосновая полевка (Microtus pinetorum) — моногамный вид грызунов: самцы помогают самкам выращивать потомство.
16. Неспособность нести ответственность за собственные действия
16. Неспособность нести ответственность за собственные действия Самые знаменитые слова Гито: «Врачи убили Гарфилда; я всего лишь выстрелил в него». Иначе говоря, он не взял на себя ответственность за смерть президента. Гито упрекал жену в их бедности, заявляя,
Глава 6. Электромагнитные поля меняют поведение
Глава 6. Электромагнитные поля меняют поведение Отмечено многочисленными исследователями, что разные ЭМП могут менять двигательную активность (ДА) организма, изменять чувствительность к раздражителям, нарушать формирование условных рефлексов и угнетать память. Все эти