Глава 6. Мобилизующий отбор
Глава 6. Мобилизующий отбор
Эволюция подотряда Ammonitina протекала необычайно бурно. Все морфологические особенности раковины — и внешняя форма, и скульптура, и перегородки — проявляли при этом сильнейшую мобильность. <…> Способы усложнения лопастной линии были также необычайно многообразны.
Б.Е. Руженцев
Мутабильность
К проблеме генетической сложности структуры можно подойти еще с одной стороны. Дело в том, что специальные гены, как и всякие другие, обязаны мутировать, порождая внутрипопуляционную изменчивость по мощности соответствующих рабочих структур. Заметим, что мощность морфологической структуры, с точки зрения генетика, следует отнести к мерным (количественным) признакам, для которых типичен непрерывный характер изменчивости. Мутации по этим признакам можно разделить на две четко очерченные категории — с качественным и количественным эффектом. Первые вызывают настолько резкое изменение значения признака, что наблюдатель видит их эффект без всяких измерений. Примерно также проявляет себя и потеря гена, поэтому в основе мутаций с качественным эффектом обычно лежит серьезное повреждение функции гена.
Мутации второй категории ведут к небольшому (на несколько процентов) сдвигу величины признака, и этот эффект, как правило, меньше шума среды. Еще одна особенность данных мутаций заключается в том, что они могут не только снижать, но и повышать значение признака. Поэтому природные популяции нередко хорошо отвечают на отбор по мерным признакам в обоих направлениях. За мутациями этого типа и за возникающими в результате их действия аллелями исторически закрепилось несколько неудачное название полигенных.
В гл.4 мы рассмотрели аддитивную модель наследования количественных признаков. Суть ее сводилась к тому, что величина признака складывается из суммы примерно равных вкладов большого числа генов. Обширный опыт биометристов показывает, что реальная картина наследования количественных признаков весьма часто начинает неплохо соответствовать аддитивной модели, если их измерять логарифмической, а не обычной (арифметической) шкалой. Особенно четко «улучшающий» эффект логарифмирования проступает, когда значение признака у родителей отличается в несколько раз. Столь большая разница обычно бывает у форм, разделенных или длительной эволюцией, или длительной селекцией. Следовательно, разность логарифмов величины признака родительских форм можно трактовать как сумму соизмеримых эффектов аллельных замещений по значительному числу локусов (см. (5.6)). Аддитивность логарифма отражает мультипликативность величины признака, измеренного арифметической шкалой. Иными словами, эта величина представима в виде произведения близких по значению сомножителей, за каждым из которых стоит отдельный ген.
В предыдущем разделе мы показали, что мультипликативный характер генетического контроля размеров и мощности рабочей структуры возникает в результате весьма длительного движущего отбора, направленного на интенсификацию ее функции. При этом идет аккумуляция генов (названных нами специальными), управляющих программой развития структуры.
Наши знания о скоростях полигенных мутаций базируются на данных, полученных при исследовании очень важного количественного признака — жизнеспособности. Мы уже знаем (гл.4), что мутации по этому признаку разделяются также на две категории: 1) рецессивные летали и полулетали, снижающие жизнеспособность не менее чем в 10 раз; 2) слегка вредящие мутации, снижающие в гомозиготном состоянии жизнеспособность всего на 2–4 %. Напрашивается явная аналогия с только что рассмотренным делением на две категории мутаций по обычным мерным признакам. Летали и полулетали подобны мутациям с качественным эффектом, а слегка вредящие мутации можно считать аналогами полигенных. Т. Мукаи так и назвал их «полигенными мутациями по жизнеспособности». Напомним, что этот японский генетик, следя за возникновением мутаций обеих категорий, сделал важное открытие — он установил, что у дрозофилы полигенные мутации по жизнеспособности возникают в 20–30 раз чаще, чем летали и полулетали.
С эволюционной точки зрения наибольший интерес представляют именно полигенные мутации. Во-первых, они относительно безвредны, т. е. незначительно сказываются на основных показателях приспособленности, и, во-вторых, по скорости возникновения, по-видимому, не менее чем на порядок превосходят мутации с качественным эффектом.
Молекулярная природа генов с количественным эффектом
Здесь мы коснемся сложного вопроса, еще далекого от полной ясности, однако весьма немаловажного для последующего изложения, — молекулярной организации наследственного вещества. Хромосома многоклеточных — это очень длинная (до нескольких сантиметров) единая молекула ДНК. Хотя она и является носителем наследственной информации, из этого совсем не следует, что любая нуклеотидная последовательность генома обязательно влияет на приспособленность. Анализ строения ДНК многоклеточных показывает, что генетическая информация (по крайней мере, доступная нашему пониманию) размещена вдоль этой гигантской молекулы далеко не равномерно. Более того, нет сомнений, что гены занимают в ней только небольшую часть. При этом сам ген состоит из совершенно разных по информационному содержанию сегментов — экзонов и интронов. Первые чаще всего кодируют первичную структуру какого-нибудь белка, а вторые обычно ничего не кодируют (рис. 26).
В межгенных промежутках (а иногда и в интронах) разбросаны нуклеотидные последовательности, имеющие отношение к генной регуляции. Активность гена проявляется в производстве им своего слепка — молекулы РНК, из которой затем вырезаются участки, соответствующие интронам. Заметим, что транскрипция РНК является лишь одним из этапов экспрессии гена. Другие этапы включают в себя синтез, сборку и созревание белковых молекул, а также их транспорт и встраивание в соответствующие субклеточные структуры.
В многоклеточном организме большое число генов находится под жестким контролем программ развития всевозможных морфологических структур, и многие из них активны только в клетках определенного типа. Многочисленные данные, полученные в последнее время, свидетельствуют, что такой контроль осуществляется согласованным взаимодействием регуляторных белков (транс-факторов) с довольно короткими и весьма специфичными последовательностями ДНК (цис-элементами). Большинство цис-элементов расположено невдалеке от точки начала транскрипции гена; одни из них ответственны за тканеспецифичность генной экспрессии, другие — за ее уровень.
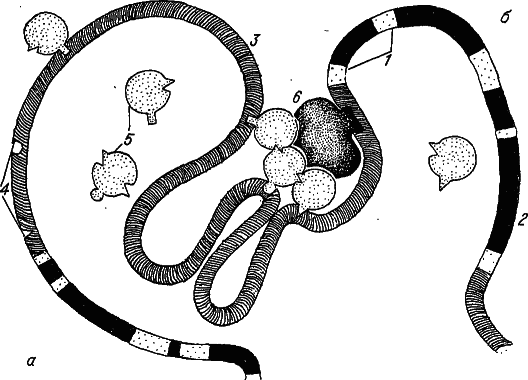
Рис. 26. Строение сегмента хромосомы эукариот. а — неактивный, б — активный ген. 1 — экзоны; 2 — интроны; 3 — межгенные участки; 4 — цис-элементы; 5 — транс-факторы; 6 — РНК-полимераза.
Для включения гена (т. е. для инициации транскрипции РНК) необходимо образование комплекса транс-факторов с белками транскрипционной машины (представленной прежде всего субъединицами РНК-полимеразы). При этом цис-элементы сближаются, а отрезки ДНК между ними сгибаются в петли. Таким образом, большой сегмент хромосомы с геном и протяженными (до нескольких тысяч нуклеотидов) соседними последовательностями выступает как единая матрица для сборки мультимолекулярного ансамбля, инициирующего транскрипцию. Понятно, что изменение числа цис-элементов и даже изменение расстояния между ними может отразиться на скорости сборки такого ансамбля, т. е. в конечном счете на уровне генной экспрессии.
Хромосомы многоклеточных буквально начинены так называемыми мобильными элементами — всевозможными последовательностями ДНК, способными к независимому «размножению» и передвижению внутри генома. По-видимому, именно эти перемещения ответственны за возникновение большей части так называемых спонтанных мутаций, составляющих основу генетической изменчивости природных популяций. Легко представить, что встраивание довольно крупного отрезка чужеродной ДНК в экзонный участок гена приведет к инактивации последнего, а соответствующая мутация пополнит ряды рецессивных леталей, подлежащих удалению очищающим отбором. Однако гораздо чаще мобильные элементы должны попадать в протяженные межгенные промежутки, что может отразиться лишь на экспрессии расположенных по соседству генов. На уровне фенотипа такие изменения наследственного материала будут выглядеть скорее всего как полигенные мутации.
Мукаи проследил за уровнем экспрессии гена алкогольдегидрогеназы (фермента, восстанавливающего спирт) у дрозофилы в течение 300 поколений. Хромосома с этим геном была выведена из-под контроля очищающего отбора, что позволяло проследить за накоплением спонтанных мутаций, затрагивающих активность фермента. За время наблюдения межлинейная дисперсия по ферментативной активности возросла в 27 раз, хотя среднелинейное значение активности осталось прежним. Также не изменились и физико-химические свойства алкогольдегидрогеназы, обусловленные первичной структурой белка. Мукаи предположил, что мутации затронули какие-то районы, расположенные за пределами гена.
Мобильность
По Дарвину, эволюция основана на отборе особей с едва заметными уклонениями фенотипа. Поскольку отбор направлен на изменение величины определенного количественного признака, принципиальное значение имеет не мутабильность генов, а мутабильность признаков. Существует несколько способов измерения последней. Например, каждая особь изогенной линии самоопыляющихся растений может стать основательницей дочерней сублинии. Если все особи-основательницы имеют тождественные генотипы, то средние значения количественных признаков в сублиниях будут совпадать. Если же особь-основательница несет мутацию, затрагивающую развитие одного из количественных признаков, то среднее значение данного признака у потомков мутантною индивида скорее всего будет смещено. Следовательно, определяя генотипические значения признака в большой совокупности дочерних сублиний, можно оценить уровень его мутабильности. Такого рода анализ был проведен у кукурузы и у мыши, в обоих случаях получена удивительно высокая величина мутабильности — около 0,01 мутаций на признак на геном за поколение.
Таким образом, в результате мутирования в любой крупной популяции за одно поколение должна возникать новая генетическая дисперсия по любому количественному признаку. Получается, что даже изогенная линия может отвечать на отбор. Как мы помним (см. уравнение (4.19)), величина этого ответа пропорциональна наследуемости признака, т. е. доле генетической компоненты в его фенотипической дисперсии. В экспериментах по отбору на число щетинок в изогенных линиях дрозофилы скорость появления мутационной дисперсии оценена в несколько тысячных от величины фенотипической дисперсии. Длительный и интенсивный отбор должен вести к постепенному исчерпанию запасов генетической изменчивости в популяции. Поэтому ответ на отбор в конце концов будет определяться лишь скоростью возникновения мутационной дисперсии.
Процесс мутирования по разным генам независим, поэтому в первом приближении можно считать, что мутационная дисперсия по величине признака, возникающая за поколение, равна сумме мутационных дисперсий по каждому локусу. Выходит, что с ростом генетической сложности признака должна расти и мутационная дисперсия его величины. В предыдущей главе мы установили, что увеличение числа специальных генов ведет к росту размеров и мощности рабочей структуры. В связи с этим приобретает особое значение вопрос, как будут соотноситься между собой обе характеристики структуры (мощность и мутационная изменчивость) при увеличении числа специальных генов.
В качестве меры изменчивости структуры, обусловленной мутированием одного локуса, удобно взять сигму мутационного эффекта (?0), которую можно трактовать как наиболее вероятное значение сдвига мощности структуры при мутировании одного из ее специальных генов. Так как естественный отбор «измеряет» эффект мутационных замен логарифмической линейкой, то имеет смысл оценивать величину ?0 в долях от мощности интактной структуры (напомним, что ?lnP ? ?Р/Р).
Если бы мощность органа определялась суммой вкладов его специальных генов, то величина относительного прироста мощности при добавлении к программе развития еще одного гена была бы тем меньше, чем выше генетическая сложность органа. Очевидно, что в данном (аддитивном) случае генетического контроля накопление специальных генов вело бы к созданию рабочих структур, мало чувствительных к мутациям, так как величина сигмы мутационного эффекта уменьшалась бы с увеличением числа их специальных генов.
Однако обычно мощность органа представима в виде произведения сомножителей, олицетворяющих собой вклады отдельных специальных генов, поэтому добавление еще одного такого гена увеличивает мощность органа в некоторое число раз (вне зависимости от его мощности или от числа специальных генов), т. е. величина относительного прироста мощности рабочей структуры не зависит от ее генетической сложности. В данном (мультипликативном) случае генетического контроля увеличение числа специальных генов не снижает чувствительности органа к мутациям, поскольку любой сдвиг (в процентах) вклада отдельного гена представляет собой сдвиг на тот же процент мощности органа. Таким образом, сигма мутационного эффекта на мощность рабочей структуры не должна зависеть от числа ее специальных генов.
Начиная с Дарвина неоднократно используется метафора, что естественный отбор как бы лепит новые формы, поэтому сам собою возник вопрос о пластичности материала, с которым имеет дело эволюция. Часто высказывалось, по-видимому, совершенно правильное мнение, что биологический материал не пластилин, что формирование структур подчиняется своим законам и т. д. Не входя в дискуссию о «законах» образования биологических структур, заметим, что с функциональной точки зрения имеет значение только их мощность. В связи с этим вместо несколько неопределенной пластичности биологического материала лучше говорить об эволюционной изменчивости мощности рабочих структур. Эту изменчивость назовем мобильностью и будем ее оценивать по скорости, с которой под действием длительного движущего отбора изменяется мощность структуры. А учитывая «логарифмические очки» естественного отбора (т. е. его «склонность» оценивать не абсолютный, а относительный сдвиг величины признака), назовем мобильностью скорость, с которой изменяется логарифм мощности рабочей структуры под действием длительного движущего отбора определенного давления. Еще раз напомним, что небольшой относительный сдвиг величины признака (измеренный в долях от его исходного значения) можно трактовать как сдвиг логарифма величины признака.
Как ответ на длительный отбор мобильность (М) должна прямо зависеть от скорости возникновения мутационной дисперсии логарифма мощности структуры, что, в свою очередь, определяется числом специальных генов (N), их средней мутабильностью и средней дисперсией мутационного эффекта <?02>. Поскольку мощность структуры определяется мультипликативным действием ее специальных генов, то сигма мутационного эффекта ?0, измеренная в долях от мощности интактной структуры, не зависит от числа этих генов. Кроме того, число специальных генов любой морфологической структуры довольно велико, поэтому значение <?02> для разных структур, по-видимому, различается незначительно. Наконец, по средней мутабильности специальных генов рабочие структуры также не могут существенно различаться. Таким образом, с достаточной точностью должно выполняться соотношение
М ~ N, (6.1)
т. е. мобильность структуры пропорциональна числу ее специальных генов.
Перевернув назад несколько страниц, можно убедиться в том, что совсем недавно мы установили почти тривиальную связь между мощностью структуры у среднестатистического вида филетической группы, с одной стороны, и числом его специальных генов — с другой (см. (5.10)). Сопоставив это соотношение с (6.1), получаем, что
<lnP> ~ <М>, (6.2)
т. е. в пределах филетической группы средневидовой логарифм мощности рабочей структуры пропорционален ее средневидовой мобильности.
Дифференциальное выживание филетических линий
Представим себе совокупность родственных видов, обитающих в сходных экологических условиях. Пусть все особи этих видов снабжены набором гомологичных рабочих структур. Отличия касаются лишь их размеров и деталей строения. Вообразим, что в среде обитания данной таксономической группы начались однонаправленные изменения какого-то жизненно важного фактора. В таких условиях мощность одной из рабочих структур может стать недостаточной для сохранения прежней численности популяций. Теперь любой вид должен успеть скомпенсировать ухудшение среды усилением мощности лимитирующей структуры, иными словами, судьба вида попадает в зависимость от скорости усиления этой структуры. Если данная скорость не сможет затормозить процесс падения абсолютной приспособленности вида, то он вымрет.
Пусть перед каждым видом поставлена одна и та же задача — увеличить мощность гомологичной структуры на один и тот же процент (скажем, на 30 %). Быстрее всего эту задачу можно решить, усилив экспрессию нескольких специальных генов, но для этого вид должен успеть за отведенный ему судьбою срок зафиксировать несколько благоприятных мутаций. Темп фиксации последних при прочих равных условиях пропорционален скорости их возникновения, а она тем выше, чем больше генов участвует в контроле развития структуры. Таким образом, мы получаем удивительный результат: чем сложнее устроена лимитирующая структура, тем больше шансов у обладающего ею вида на филетическое выживание, и, наоборот, чем проще она устроена, тем больше у вида шансов на вымирание. Напомним, что для равенства всех прочих условий степень благоприятности мутаций не должна зависеть от числа специальных генов, а это выполняется при мультипликативном характере генетического контроля.
Генетически сложная структура может быстро изменить свою мощность и в противоположном направлении, т. е. снизить ее. Для этого достаточно ослабить экспрессию нескольких специальных генов. Поэтому сложно устроенная структура сообщает виду повышенные шансы на выживание даже в ситуации, когда среда как бы благоприятствует более примитивным. Вид — обладатель генетически примитивной структуры не может быстро сделать ее еще примитивнее, так как слишком мало генов контролирует ее развитие. Таким образом, вне зависимости от того, чего требует окружающая среда от рабочей структуры — усиления или ослабления — главное, что определяет судьбу вида, заключается в скорости изменения мощности лимитирующей структуры. Во всех случаях больше шансов на выживание имеют виды, у которых такая структура управляется большим числом генов.
Это обстоятельство можно выразить в терминах более обычных для теории эволюции: на виды, эволюционирующие в одной адаптивной зоне, постоянно действует отбор, благоприятствующий увеличению генетической сложности структур, от которых среда регулярно требует усиления или ослабления функции. Такой отбор имеет смысл назвать мобилизующим, так как главная «цель», которую он преследует, заключается в увеличении мобильности структур, наиболее часто лимитирующих скорость адаптации.
Итак, мобилизующий отбор ведет к постоянному увеличению числа специальных генов лимитирующей структуры. Откуда эти гены берутся? Создать совершенно новый ген за короткое время невозможно. Может быть где-то в геноме хранится запас готовых к употреблению генов? Однако такого запаса нет и быть не может, так как если ген не работает, то он не проверяется на качество очищающим отбором. Аллельный состав такого гена подчиняется только законам нейтральной эволюции, поэтому под действием мутационного давления через несколько миллионов лет все локусы генного резерва были бы замещены бессмысленными аллелями, превратились бы, как говорят, в хлам из ДНК.
Однако для увеличения числа специальных генов, контролирующих развитие лимитирующей структуры, можно воспользоваться вполне исправными генами, которые в данный момент обслуживают другие структуры. Такой ген (из сферы обслуживания других функций) можно было бы удвоить (дуплицировать), а его дополнительную копию, снабдив новыми контролирующими элементами, направить в распоряжение лимитирующей структуры. Хотя этот путь вполне реален, у него есть один недостаток — он ведет к увеличению наследственной информации генома, а следовательно, и к росту мутационного генного груза.
Конечно, добавление одного лишнего гена — пустяк, и очищающий отбор этого даже не заметит. Но надо полагать, объем наследственной информации генома был доведен до предельной черты еще на заре эволюции многоклеточных. Мы уже упоминали о гипотезе А.С. Кондрашова, по которой даже разделение особей на два пола, несмотря на все связанные с этим энергетические и информационные издержки, служит главным образом цели снижения мутационного груза. Таким образом, генные дупликации как стратегическое направление едва ли перспективны. Куда изящнее выглядит просто смена геном своей специализации (его перепрофилирование), когда ген, управлявший ранее развитием одной структуры, после соответствующей мутации начнет повышать мощность другой, лимитирующей, структуры.
Кроме того, вполне вероятно, что в измененной среде мощность некоторых структур может оказаться избыточной. На первый взгляд, это может показаться странным, но достаточно вспомнить о видах, заброшенных эволюционной судьбой в подземные пещеры. Многочисленные структуры зрительного анализатора, покровительственная окраска, формы поведения, рассчитанные на зрительное восприятие, — все это в полной темноте становится избыточным, излишним. Временная избыточность структур открывает путь к перепрофилированию их специальных генов. Развивая эту мысль, можно допустить, что изменение среды ведет к «перекачке» генов из системы обслуживания структур с избыточной мощностью в состав специальных генов лимитирующей структуры.
Генная специализация
Организм представляется нам совокупностью взаимодействующих друг с другом морфологических рабочих структур, при этом на клетки мы смотрим как на элементарные единицы нескольких типов, которые могут размножаться и производить молекулярные продукты, нужные не им самим, а организму в целом. Такими продуктами могут быть и пищеварительные ферменты, и гормоны, и эмбриональные индукторы. Поскольку функция генов, управляющих синтезом этих продуктов, заключается в создании над клеточного уровня организации, мы можем назвать их генами межклеточного взаимодействия.
В то же время нам хорошо известно, что клетки далеко не атомы. Для реализации основных клеточных процессов — репликации ДНК, синтеза и созревания РНК, синтеза белков, производства и запасания энергии, работы митотического аппарата и многого другого — задействованы тысячи генов. Их принято называть генами домашнего хозяйства. Таким образом, все гены многоклеточного организма можно разделить на два обширных класса — гены межклеточного взаимодействия, ответственные за создание надклеточных структур, и гены домашнего хозяйства, нужные для работы любой клетки.
Гены с новой молекулярной функцией появляются в процессе эволюции очень редко, поэтому рядовой эволюционный сдвиг межвидового ранга связан не с появлением качественно новых генов, а с изменением времени, места и, главное, уровня экспрессии уже имеющихся. Для увеличения числа специальных генов необходимо выполнение двух условий. Во-первых, какой-нибудь «посторонний» ген нужно снабдить новым цис-элементом, чтобы он мог работать в клетках, формирующих рабочую структуру, во-вторых, перепрофилированный ген должен включиться в программу развития лимитирующей структуры, т. е. он увеличит ее мощность. Правда, события подобного рода не могут происходить часто, так как в окрестности специального гена должна появиться вполне конкретная, хотя и короткая, последовательность ДНК.
Вместе с тем уже простого изменения взаимного расположения цис-элементов вблизи кодирующих последовательностей может хватить для ощутимого сдвига уровня генной экспрессии. Очевидно, что такого рода мутации могут происходить не так уж редко. Однако какой бы малой ни казалась нам вероятность перепрофилирования локусов, прямой анализ строения генов, обслуживающих морфологические структуры, часто указывает на их происхождение от генов домашнего хозяйства. Особенно впечатляют в этом отношении белки хрусталика глаза — кристаллины. ?-кристаллины явно родственны белкам теплового шока, а ?-кристаллин птиц и рептилий фактически является одной из форм фермента метаболизма глюкозы — лактатдегидрогеназы.
Основные функции многоклеточного организма в значительной мере обеспечиваются его надмолекулярными структурами. Каждая из них возникает в ходе индивидуального развития под контролем своих специальных генов, работа которых так тонко увязана между собой (в программу развития), что изменение экспрессии любого из них немедленно отражается на функции взрослой структуры. Фактически это означает, что возникающие с повышенной частотой мутации по генной экспрессии начинают выступать в роли мутаций, ослабляющих или усиливающих мощность рабочих структур. Отметим, что изменения наследственного материала, ведущие к увеличению числа специальных генов надклеточных рабочих структур, можно с полным основанием отнести к мутациям, повышающим «выход» благоприятных мутаций.
Математическая модель мобилизующего отбора
Попробуем более формально проанализировать механизм действия мобилизующего отбора. Представим себе, что в какой-то адаптивной зоне обитает большое число родственных видов, обладающих одним и тем же признаком — гомологичной и изофункциональной рабочей структурой. Пусть величина этого признака (под которым в данном случае мы понимаем логарифм мощности структуры) определяется суммарным вкладом его специальных генов. Каждый из них может находиться в двух аллельных состояниях 1 и 0. Аллель 1 соответствует активному состоянию гена (когда он экспрессируется), аллель 0 — отсутствию экспрессии. Таким образом, величина признака (Z) определяется числом специальных генов, находящихся в активном состоянии. Полное число генов (G) в геноме разных видов постоянно, тогда как число специальных генов (N), определяющих степень развития признака, различно. Гены, не вошедшие в состав специальных (их число равно G — N), будем называть «сторонними». Они определяют развитие каких-то других (сторонних) признаков.
В среде обитания время от времени происходят медленные однонаправленные изменения, когда над каждым видом нависает реальная угроза гибели. Избежать вымирания вид может за счет компенсаторного изменения величины признака на какое-то число единиц. (Что соответствует увеличению мощности рабочей структуры в какое-то число раз.) Увеличение признака на одну единицу может произойти за счет фиксации мутаций двух типов. Во-первых, это могут быть мутации, активирующие специальные гены, и, во-вторых, мутации, переводящие ген из разряда «сторонних» в разряд специальных (с аллельным состоянием 1). Точно так же сдвиг признака в сторону его уменьшения может быть достигнут либо инактивацией одного специального гена, либо переводом активного специального гена в состав «сторонних».
Для своего филетического выживания каждый вид должен за единицу макроэволюционного времени осуществить фиксацию нескольких благоприятных мутаций (любого типа), изменяющих признак на требуемую величину ?Z. Донорами этих мутаций являются чаще всего специальные гены признака, так как изменение экспрессии гена — более вероятное событие, чем его перепрофилирование. Соответственно вероятность филетического выживания вида (его адаптируемость) будет расти с увеличением числа специальных генов (N). Это создает предпосылки роста генетической сложности признака.
Такая простая модель позволяет осуществить машинный эксперимент. На каждом шаге модели в окружающей среде каждого вида происходят случайные изменения, которые никак не связаны с изменением среды обитания остальных видов. От одних видов среда требует увеличения признака, а от других — уменьшения. Всю эту картину можно передать типом распределения величины ?Z. В данном эксперименте мы остановились на биномиальном симметричном распределении. С подробностями вычисления вероятностей выживания видов в новых условиях среды можно ознакомиться в книге «Основные факторы макроэволюции».

Рис. 27. Моделирование динамики видовых распределений по величине признака (а) и числу его специальных генов (б). Приведенные распределения (слева направо) соответствуют 20, 50, 100, 200, 800-му шагу модели. Последнее распределение финальное.
Проследим за изменением видовых распределений по величине признака (рис. 27, а). В начальный момент все виды обладают минимальным значением признака, поскольку его развитие обеспечивает всего один специальный ген. Однако завершается машинная эволюция широким финальным распределением со средним значением, равным 18,4. Таким образом, несмотря на то, что среда с равной вероятностью благоприятствует как увеличению, так и уменьшению значения признака, его средневидовая величина (<Z>) неуклонно растет. Правда, параллельно с этим нарастает и уровень межвидовых различий по степени развития признака (?z) (рис. 28). В итоге даже в финальном распределении после долгой эволюции сохраняется довольно много видов с «архаичной», «примитивной», величиной признака.
По-иному ведет себя распределение видов по числу специальных генов (рис. 27, б). По скорости роста среднего значения оно значительно опережает рассмотренное выше распределение по величине признака. Напомним, что рост числа специальных генов отражает усложнение генетической программы развития признака. Своеобразие динамики распределения видов по числу специальных генов состоит в том, что его размах стабилизируется гораздо раньше, чем среднее значение; поэтому в течение длительного отрезка времени мы наблюдаем практически параллельный снос вправо всего распределения. Следовательно, аккумуляция специальных генов идет со сходной скоростью во всех филетических линиях нашей эволюционирующей группы. Финальное распределение соответствует крайней специализации видов в отношении данного признака, поскольку большая часть генов генома попадает в разряд специальных (их среднее число равно 36,2). Ясно, что завершение процесса специализации обусловлено исчерпанием возможностей генома, число генов которого в данной модели неизменно (G=50).
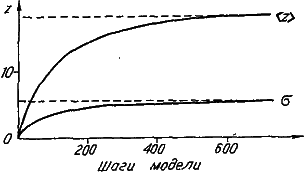
Рис. 28. Изменение среднего значения (<Z>) и сигмы (?) распределений, представленных на рис. 27.
Так как мобильность признака определяется прежде всего числом специальных генов, то рост их среднего числа можно интерпретировать как рост мобильности признака. Мы видим, что модель вполне адекватно передает суть мобилизующего отбора.
Сравнивая оба типа финальных распределений (по N и по Z), следует отметить, что виды «архаичные» и «примитивные» в отношении величины признака оказываются вполне «продвинутыми» по его генетической сложности. Кажущаяся архаичность, очевидно, обусловлена низким уровнем экспрессии большинства специальных генов. Однако мобильность признака у этих «псевдоархаичных» видов довольно высока, поэтому, если начать эволюцию именно с них, финальное распределение достигается гораздо быстрее.
При сравнении данных модельных распределений с реальными распределениями аммоноидей по сложности сутуры нельзя не отметить их разительного сходства, которое распространяется даже на пропорциональность среднего значения и сигмы. Все это позволяет нам предположить, что сущность прогресса лопастной линии состояла в увеличении ее мобильности и, по-видимому, эта цель была достигнута аммоноидеями к началу юры, когда они «освоили» все способы рассечения сутуры.
Принцип максимальной адаптируемости
За долгие 700–800 миллионов лет истории многоклеточных предки каждого современного вида прошли через сотни (а может быть, и тысячи) актов проверки на адаптируемость. В связи с этим на каждую особенность строения современных организмов мы можем взглянуть как на приспособление, повышающее шансы вида на филетическое выживание. Иными словами, организация современных живых существ в какой-то степени должна отвечать принципу максимальной адаптируемости, т. е. максимальной скорости фиксации благоприятных мутаций.
Рассмотрим филетическую группу гипотетических организмов, обладающих всего двумя функционально независимыми рабочими структурами А и Б. Во внешней среде временами случаются два типа изменений: один «требует» усиления (или ослабления) мощности структуры А, а другой предъявляет аналогичные требования к структуре Б. Хотя эти изменения происходят совершенно случайно, их можно охарактеризовать своими частотностями (вероятностями). Например, изменения первого типа могут случаться в 2 раза чаще, чем второго, тоща суммарное время, в течение которого мобилизующий отбор «увеличивает» число специальных генов структуры А, будет в 2 раза больше, чем время, когда он «добавляет» гены к структуре Б. Если число генов в геноме постоянно, то через некоторое время между специальными генами обеих структур должно установиться динамическое равновесие в соответствии с вероятностями требований к ним среды. При этом структура А «перетянет» на себя в 2 раза больше генов, чем структура Б.
Заметим, что именно специальные гены и являются основными поставщиками благоприятных аллелей для движущего отбора, когда среда «требует» изменения той или иной структуры. Очевидно, что по «выходу» благоприятных мутаций структура A в 2 раза превосходит структуру Б, точно отражая соотношение численностей их специальных генов (а стало быть, и мобильностей). Иными словами, при одном и том же давлении отбора на обе структуры благоприятные мутации по структуре А будут возникать и фиксироваться вдвое чаще, чем по структуре Б.
Теперь снабдим наши гипотетические организмы большим числом функционально независимых структур, при этом будем считать, что адаптивная зона предъявляет к каждой из них требования со своей характерной частотой.
Совершенно ясно, что после длительной эволюции между численностями специальных генов отдельных структур установятся равновесные соотношения, которые будут соответствовать вероятностям требований среды к изменению их мощности. Фактически это можно трактовать как приобретение организмами знаний о своей адаптивной зоне. Филетическая линия как бы изучает среду обитания и приобретенные знания воплощает в генетическую сложность морфологических структур. Чем чаще структура лимитировала адаптацию в прошлом, тем она сложнее, тем выше выход благоприятных мутаций при отборе, направленном на ее изменение, и тем легче будет вид преодолевать в грядущем изменения среды, типичные для его адаптивной зоны. В итоге максимальной мобильности достигает орган, чья мощность чаще всего оказывается недостаточной (или избыточной) для проникновения вида в соседнюю экологическую нишу. Это означает, что в филетических линиях, вполне освоивших свое экологическое пространство, выход благоприятных мутаций достигает максимума.
Более того, сам характер генного контроля мощности рабочих структур (мультипликативный) отвечает принципу максимальной адаптируемости. Во-первых, эффект мутации по любому специальному гену одинаково хорошо заметен для отбора и, во-вторых, величина этого эффекта не зависит от числа специальных генов. Такой тип генного контроля, очевидно, является следствием длительного движущего отбора, способного «видеть» лишь, во сколько раз (т. е. на какой процент) рабочая структура у особи с благоприятным аллелем развита сильнее (или слабее), чем у остальных членов популяции (см. гл.5). Повышенная чувствительность структуры к мутациям достигается при подчинении ее специальных генов единой программе развития, когда активность более поздних генов определяется активностью более ранних. Таким образом, привычный для нас характер возникновения структур в ходе онтогенеза соответствует принципу максимальной адаптируемости.
Мобилизующий отбор в простой среде
Мы всегда склонны считать окружающий мир сложным. Выражение «жизнь сложна» воспринимается нами как аксиома. Однако посмотрим на нашу жизнь попристальнее. За выполнение довольно простой и монотонной работы мы получаем деньги, которые затем обмениваем на пищу и кров. Кроме того, существующая плотность населения вполне достаточна для гарантии случайной встречи с особью другого пола. Теперь посмотрим, как решал те же проблемы первобытный человек. Попробуйте-ка выследить оленя и застрелить его из лука или найти в лесу съедобные растения и прокормить ими себя и своих детей. Да и спутника жизни обычно приходилось искать за тридевять земель на территории, занятой другим, нередко агрессивно настроенным родом. Сколько опасностей нужно было избежать при решении всех этих задач. Ясно, что жизнь современного человека намного проще.
Адаптивные зоны филетических групп также различаются по степени сложности. В простых зонах повышенные требования предъявляются к одной из основных функций, поэтому действие мобилизующего отбора направлено на специализацию генов в отношении ограниченного круга рабочих структур. В итоге данные структуры усложняются, а большинство других упрощается (деградирует). Такая эволюционная стратегия традиционно называется специализацией и не считается прогрессом, хотя, с чисто структурной точки зрения, некоторые органы достигают удивительной сложности. Повышенная мобильность небольшого числа структур обеспечивает соответствующие филетические группы высоким темпом ветвления в простых адаптивных зонах. Беда только, что сами эти зоны не отличаются долговечностью, а с их исчезновением гибнет и все многообразие специализированных форм. По-видимому, именно эта стратегическая обреченность специализации не позволяет нам признать ее структурные успехи за прогресс.
В отличие от прогресса проблема деградации привлекает к себе меньше внимания, что, вероятно, связано с оптимистичностью нашей натуры. Под деградацией обычно понимают вторичное упрощение рабочих структур. Так, переход к паразитизму ведет к ликвидации большинства структур, ранее обслуживавших функции доставки, удаления, гомеостаза и охраны. Червь, проникший в наш кишечник, получает доступ к уже переваренной пище. Ему не надо бояться хищников, поскольку это наша забота. От него же требуется только защититься от пищеварительных ферментов с помощью специальной кутикулы, прикрепиться с помощью крючков и присосок к стенке нашего кишечника и все. Единственной неудовлетворенной потребностью остается размножение. Правда, структуры, призванные для разрешения этой проблемы, достигают здесь необыкновенной сложности.
Важнейшим приспособлением к паразитическому образу жизни обычно является колоссальная плодовитость. Так, у трематод мало того, что взрослые черви производят огромное число яиц, но и их личинки, проходя несколько стадий, могут на любой из них размножаться бесполым путем. При этом нередко каждая личиночная стадия протекает только в организме специфичного промежуточного хозяина.
В то же время, если среда обитания вида-хозяина стала изменяться катастрофическим образом, то виды-паразиты должны успеть покинуть его (как крысы тонущий корабль) и подобрать себе другого хозяина. Адаптируемость для них — это прежде всего легкость смены видов-хозяев. Поэтому «деградацию» в данном случае можно рассматривать как колоссальный прогресс функции воспроизведения, «оттянувшей» на себя большую часть генов межклеточного взаимодействия. Эффективность такой стратегии демонстрирует и таксономический успех паразитов. Например, тип плоских червей насчитывает около 12,5 тысячи видов, из которых свободный образ жизни в сложных средах ведет всего около трех тысяч, остальные (9,5 тысячи) — паразиты.
Мобилизущий отбор в сложной среде
Иная судьба складывается у филетических групп, эволюционирующих в сложных адаптивных зонах, где регулярно предъявляются требования к гораздо большему числу рабочих структур. В этих условиях мобилизующий отбор не может обеспечить одну из них большим числом специальных генов, поэтому рабочие структуры чаще всего сохраняют «примитивный», «архаичный» вид. Эволюцию таких форм нельзя назвать бурной, их скромные структурные успехи теряются на фоне ярких достижений специализированных филетических групп. Зато овладение сложной средой обеспечивает архаичные (неспециализированные) формы весьма продолжительным существованием.
Вместе с тем для этих «прозябающих» неспециализированных групп существует одна возможность — под давлением мобилизующего отбора резко поднять уровень адаптируемости. Она состоит в объединении специальных генов двух (или большего числа) рабочих структур для контроля развития одной новой, мобильность которой после такого объединения должна существенно возрасти. Данные сравнительной морфологии свидетельствуют о весьма широком распространении эволюционной тенденции к созданию морфологически сложных (составных) органов, возникших после интеграции нескольких более простых.
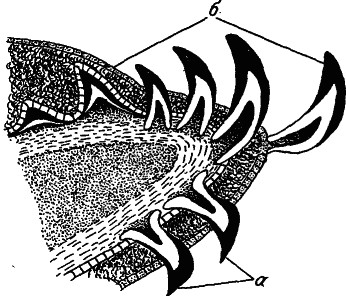
Рис 29. Зубы акулы (по: [Оммани, 1975]).
Чешуи (а), покрывающие тело, заходя на поверхность челюстей, трансформируются в зубы (б).
Взять, к примеру, такое важнейшее для эволюции позвоночных событие, как появление челюстей — принципиально нового пищедобывающего органа. Это приобретение позволило рыбам «оторваться» ото дна и приступить к завоеванию всей водной толщи, поскольку челюсти открыли им доступ к новым источникам пищи.
У первичных позвоночных глотка была прорезана серией парных щелей, на краях которых располагались жабры. Каркасом для этого дыхательного органа служила система висцеральных дуг — опорных элементов, размещенных в промежутках между жаберными щелями. Челюсти возникли при объединении скелета двух передних висцеральных дуг с прочным и относительно неподвижным мозговым черепом. Элементы одной висцеральной дуги преобразовались в верхнюю и нижнюю челюсти, тогда как другая висцеральная дуга взяла на себя роль «подвеска». Третьим структурным компонентом нового органа стали зубы, вначале представлявшие собой не что иное, как слегка измененную чешую, перешедшую на ротовую поверхность челюстей (рис. 29). Итак, мы видим, что данный орган удержания и умерщвления жертвы возник на основе по меньшей мере трех совершенно разных в функциональном отношении структур.
Более всего нашему интуитивному представлению о морфологическом прогрессе отвечают составные полифункциональные органы, определяющие интенсивность сразу нескольких основных функций. Например, ясно, что органы зрения помогают и добыванию пищи, и защите от хищников, и поиску партнера по спариванию. То же верно для многих других структур, включая и такие сложные, как локомоторный аппарат, ответственный за движение животного. По большому счету, в данный аппарат помимо костей, мышц, суставов и сухожилий мы должны включить соматическую нервную систему и все органы чувств. Фактически все это составляет большую часть тела животного.
Объяснение правила Копа
Внешняя среда, несмотря на всю свою сложность, «интересует» вид в отношении очень небольшого числа вопросов: 1) есть ли в среде источники пищи для поддержания достаточно плотных популяций; 2) не слишком ли высок уровень истребления со стороны хищников и прочих биотических факторов; 3) не препятствуют ли физико-химические условия среды успешному завершению индивидуального развития.
Если вид способен существовать в течение многих сотен поколений, значит, он получает удовлетворительные ответы на все эти вопросы. Также ясно, что в условиях медленно ухудшающейся среды один из перечисленных вопросов может остаться без ответа. Какова будет адаптационная реакция вида? Много ли у него путей выхода из кризиса? Вообще для восстановления уровня абсолютной приспособленности существует несколько путей. Эволюционирующий вид их не выбирает, а идет сразу по многим, но к цели (т. е. к спасению) ведет кратчайший. Длина же пути определяется скоростью появления полезных мутаций и степенью их благоприятности.
Ситуация 1a. Поток пищи (энергии) ослабел по вине вида-конкурента. Существуют три варианта выхода. А. Усилить пищевой поток. Это возможно, если вид обладает развитой и мобильной пищедобывающей структурой. Данный вариант можно было бы назвать специальным. Б. Сохранить прежнюю биомассу при меньшей численности, т. е. увеличить размеры особи. В. Сохранить прежнюю численность при меньшей биомассе. Для этого достаточно уменьшить массу средней особи, что сопряжено с усилением удельного метаболизма и укорочением времени генерации. В итоге вид будет быстрее поглощать пищевой ресурс, чем может поставить конкурента на грань катастрофы.
Ситуация 1б. Поток пищи ослабел в связи с уменьшением ресурса ниши. Параллельно в соседних нишах произошло примерно то же самое. Решение в данном случае может быть связано только с измельчанием особей при усилении функции воспроизведения.
Ситуация 2. Повысился уровень гибели особей от хищников. Здесь можно указать на четыре варианта выхода из положения. А. Уйти от конкретного хищника, выработав специфические средства защиты (яд, шипы, панцирь и т. д.), — это специальный ответ. Б. Увеличить размеры особи, став хищнику, что называется, не по зубам. В. Скомпенсировать потери от хищника, снизив энергетические расходы единицы биомассы. Этого можно достичь, увеличив массу особи. Г. Скомпенсировать потери от хищника ускорением развития и усилением функции воспроизведения, что достигается уменьшением массы особи.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 1.4. Отбор животных по признаку уровня рассудочной деятельности
Глава 1.4. Отбор животных по признаку уровня рассудочной деятельности Значение изучения рассудочной деятельности животных трудно переоценить как с научной, так и с этической точки зрения. Большой вклад в изучение этой проблемы внес Л. В. Крушинский и его сотрудники
Естественный отбор
Естественный отбор Итак, Дарвин показал, что основным средством создания всех пород домашних животных является искусственный отбор. Даже в то далёкое время, когда люди вели этот отбор, не ставя перед собой определённой цели, бессознательно, они достигали поразительных
Естественный отбор
Естественный отбор Я не усматриваю предела деятельности этой силы, медленно и прекрасно приспособляющей каждую форму к самым сложным жизненным отношениям. Ч. Дарвин Осы, бабочки и дарвинизмВ предыдущих главах мы неоднократно говорили о естественном отборе. Это и
6. ИСКУССТВЕННЫЙ ОТБОР
6. ИСКУССТВЕННЫЙ ОТБОР В эволюции животных действовали вполне понятные законы, построенные на изменчивости, отборе и наследуемости полезных признаков. Эти незатейливые принципы распространялись на предков человека вплоть до райской эпохи. Райский период эволюции
2. Родственный отбор
2. Родственный отбор «Я бы отдал жизнь за двух братьев или восьмерых кузенов» Дж. Холдейн Создатели теории родственного отбора: Рональд Фишер (1890–1962), Джон Холдейн (1892–1964), Уильям Гамильтон (1936–2000).К этой мысли биологи стали подходить уже в 30-е годы прошлого века.
Родственный отбор
Родственный отбор Кооперация и альтруизм — краеугольные камни социального поведения Homo sapiens. Вряд ли нужно объяснять, что без кооперации (сотрудничества, взаимовыгодного поведения) устойчивое существование социума у приматов невозможно. Очевидно и то, что наши предки
Половой отбор.
Половой отбор. Так как при доместикации часто возникают у одного из полов особенности, наследственно связанные с этим полом, то, без сомнения, то же должно встречаться и в природе. Таким образом, возникает возможность модификации каждого из обоих полов путем
Половой отбор
Половой отбор Для того чтобы мы окончательно не деградировали и не вымерли, как мамонты, Ее Величество Природа придумала половой отбор, который невозможен при партеногенезе и гермафродитизме. Половой отбор заключается в выборе полового партнера по каким-либо внешним
ЕСТЕСТВЕННЫЙ ОТБОР
ЕСТЕСТВЕННЫЙ ОТБОР Естественным отбором считается процесс, который Дарвин назвал «борьба за существование», при котором наиболее приспособленные организмы выживают, а наименее приспособленные погибают. Согласно положениям дарвинизма, естественный отбор в популяции с
ОТБОР
ОТБОР Естественный отбор — это процесс, сокращающий в популяции пропорцию организмов, относительно неприспособленных к выживанию и увеличивающих пропорцию организмов с фенотипом, близким к оптимальному, для определенной среды и уровня конкуренции. Здесь важен высокий
Естественный отбор
Естественный отбор Естественный отбор – важнейший фактор эволюции. Дарвинизм (а именно на базе дарвинизма построена СТЭ), как уже отмечалось выше, называют теорией естественного отбора.Кратким и удачным определением отбора может служить сформулированное И. Лернером:
Групповой отбор
Групповой отбор Подходя в основном нормально к эволюционной психологии, Дарвин уступил искушению, известному как групповой отбор. Рассмотрим его центральное объяснение эволюции морального чувства. В "Происхождении человека" он написал, что "повышение стандартов этики и
Глава 12 Естественный отбор: кто выживет?
Глава 12 Естественный отбор: кто выживет? Есть горстка ученых, пытающихся выявить признаки, общие для всех когда-либо существовавших человеческих культур и цивилизаций. Эти ученые роются в трудах этнографов и в статьях антропологов в поисках различий между племенами и