Глава 10 Генезис речи-мышления: суггестия и дипластия
Глава 10
Генезис речи-мышления: суггестия и дипластия
I. Начальное отношение и начальное общение людей
Как условлено с первых глав, в тему этой книги входит только старт человеческой истории. Но не начальный этап истории и не начальные формы социума и этноса. Ни древнейшая дуальная организация, ни родовой строй вообще и его ступени, ни экзогамия или другие аспекты семейно-брачных отношений — ничто это не составляет темы данного философско-естественнонаучного трактата, задача которого только в том, чтобы по возможности прочнее, чем делалось до сих пор, поставить ногу на порог.
Этнологи и археологи, углубляющиеся в предысторию, начинают уже с того, что даны люди. Между последними существовали такие-то отношения, пусть тёмные и экзотичные. Мы же сперва отсекли то, что оказывается возможным отодвинуть за пределы этого понятия «люди», и тем самым лишь уточнили хронологический отрезок, охватывающий явление «начала», — и не предлагали ничего большего. Но это «начало» должно быть таким, чтобы оно содержало в себе всё будущее движение. Мало того, что движение будет его отрицать, превращать в противоположность, начало тут — то, что будет кончаться. Вопрос об институтах и структурах первобытного общества касается определённой части истории, тогда как исследуемый предмет — начало всей истории.
В частности, первобытнообщинный способ производства — это не только специфический по отношению к другим способ производства, но и первичный способ производства вообще. Это значит, что в самом субстрате первобытной экономики налицо нечто такое, что будет отрицаться дальнейшей экономической эволюцией человечества.
В основе всей истории производства вообще лежит способность людей производить больше, чем им нужно для восстановления затраченных в этом акте сил. Отсюда возможность и специализации, и обмена, и производства объектов культуры, и присвоения чужого прибавочного продукта (эксплуатации). Много думали и спорили: почему у человеческого труда есть это свойство, дающее человеку возможность прогрессирующего в истории наращивания и накопления сил, свойство производить, т. е. извлекать и перерабатывать из природной материи больше, чем необходимо для репродукции себя в виде своего организма и в виде своего потомства? Но может ли вол выполнить больше работы, чем надо его организму для восстановления затраченных сил? Да. Совершенно очевидно, что закон сохранения энергии тут ни при чём. «Свойство» человека состояло не в том, что он вообще мог производить некоторый избыток сверх своих затрат, т. е. минимальных потребностей, а в том, что он вынужден был это делать. Притом вынужден был производить этого избытка всё больше, вследствие чего должен был прибегать к орудиям, к технике, ко всему тому, что мы называем производительными силами.
Теория докапиталистических способов производства никогда не может быть достаточно полно разработана (и особенно первобытнообщинного способа производства), если не преодолеть распространения на все времена представления о «homo oeconomicus», извлечённого из капиталистической эпохи. Согласно этому ходячему представлению, хозяйственная психология всякого человека может быть сведена к постулату стремления к максимально возможному присвоению. Нижним пределом отчуждения (благ или труда), психологически в этом случае приемлемым, является отчуждение за равноценную компенсацию. «Экономический человек» отождествляется с «экономящим» — «не расточающим». Действительно, поведение, обратное указанному постулату, при капитализме не может быть ничем иным, как привеском. Но даже при феодализме, как видно из источников, хозяйственная психология содержала гораздо больше этого обратного начала: значительное число средневековых юридических и законодательных актов запрещает или ограничивает безвозмездное дарение, подношение, пожертвование недвижимого и движимого имущества. Чем дальше в глубь веков и тысячелетий, тем отчётливее этот импульс.
И в самом деле, весьма наивно перекидывать прямой мост между человеческой алчностью и инстинктами питания у животных. Алчность — не константа истории. Она созревала как противоположность и отрицание специфичного для первобытных людей «противоестественного» поведения: стремления к максимальному безвозмездному отчуждению благ. В таком утверждении нет ничего идеалистического, как нет ничего материалистического в уподоблении психики всех людей «материализму» лавочника. Материальный, коммерческий расчёт — это совсем иное понятие, чем материальная детерминированность общественной жизни. На заре истории лишь препоны родового, племенного, этнокультурного характера останавливали в локальных рамках «расточительство» и тем самым не допускали разорения данной первобытной общины или группы людей. Это значит, что раздробленность первобытного человечества на огромное число общностей или общин (причём разного уровня и пересекающихся), стоящих друг к другу так или иначе в отношении «мы — они», было объективной хозяйственной необходимостью. Но норма экономического поведения каждого индивида внутри этих рамок состояла как раз во всемерном «расточении» плодов труда: коллективизм первобытной экономики состоял не в расстановке охотников при облаве, не в правилах раздела охотничьей добычи и т. п., а в максимальном угощении и одарении каждым другого, хотя и только по сформировавшимся обычным каналам. Дарение, угощение, отдавание — основная форма движения продуктов в архаических обществах[694].
Такая экономика подразумевает соответствующую психику. Это поведение явно противоположно «зоологическому индивидуализму», да и не может быть приравнено к действию у животных, скажем, родительского инстинкта кормления детёнышей или призыву петухом куриц к найденному корму. Взаимное отчуждение добываемых из природной среды жизненных благ было императивом жизни первобытных людей, который нам даже трудно вообразить, ибо он не соответствует ни нормам поведения животных, ни господствующим в новой и новейшей истории принципам материальной заинтересованности индивида, принципам присвоения. «Отдать» было нормой отношений. Не будем углубляться дальше в предпосылки и следствия такого устройства первобытного хозяйствования в аспекте теоретической экономии. Но рассмотрим, какова стимуляция этого отказа от прямого потребления благ. Для этого продолжим изучение работы мозга.
У человека работу центральной нервной системы можно разделить на три блока: 1) сенсорно-афферентный, т. е. осуществляющий приём, анализирование, ассоциирование разнообразнейших раздражений; 2) эффекторный, т. е. осуществляющий двигательные и вегетативные реакции, в том числе большие системы действий с их поэтапной корректировкой; 3) суггестивный, т. е. осуществляющий замену указаний, поступающих с первого блока, или ответов, свойственных второму блоку, другими, вызываемыми по второй сигнальной системе. Функцию этого третьего блока называют также «регулирующей» как восприятие, так и поведение, но надо помнить, что тут речь идёт о регулировании по происхождению своему межиндивидуальном — исходящем от другого индивида или других индивидов; лишь в своём развитии впоследствии (по Выготскому — Лурия) функция, которая была раньше разделена между двумя людьми, становится способом самоорганизации деятельности одного индивида, интерпсихическое действие превращается в интрапсихическую саморегулирующуюся систему[695]; это связано с преобразованием суггестии в контрсуггестию[696].
Как уже нами было выяснено, образование третьего блока имеет свою эволюционную базу в высшей нервной деятельности у животных и подходит к своему непосредственному кануну у палеоантропов (троглодитов). Но у неоантропов происходит преобразование кардинальной важности — переход интердикции в суггестию. В морфологии головного мозга этому соответствует появление у Homo sapiens весьма развитого префронтального отдела лобной доли коры, в особенности верхней его части, за счёт крутого уменьшения объёма затылочной доли, которая в филогении троглодитид неуклонно и интенсивно разрасталась[697]. У высших животных префронтальный отдел представлен весьма незначительно по сравнению с человеческим и, по-видимому, соответствует (гомологичен) лишь тому, что находится у человека в нижней (базальной) части этого отдела, но не в верхней его части; полагают, что у них он играет роль органа, в известной мере обеспечивающего принцип доминанты в работе центральной нервной системы. На эндокранах ископаемых прямоходящих высших приматов, т. е. представителей семейства троглодитид, включая палеоантропов, он тоже выражен слабо в соответствии с покатым, убегающим лбом и низким сводом экзокрана. В количественных показателях эволюции головного мозга высших приматов, согласно В. И. Кочетковой, бурный скачок роста префронтального отдела вверх, а тем самым и всей верхней лобной доли, обнаруживается только при переходе от палеоантропов к неоантропам. Только на этом филогенетическом рубеже на смену относительно низкому черепному своду появляется наш высоко поднятый. Он и свидетельствует о появлении слова как фактора управления поведением.
Именно тут, в префронтальном отделе, осуществляется подчинение действий человека словесной задаче (идущей от другого или от самого себя) — оттормаживание остальных реакций и избирательная активизация нужных нейрофизиологических систем[698]. Соответственно мы и должны считать, что из всех зон коры головного мозга человека, причастных к речевой функции, т. е. ко второй сигнальной системе, эволюционно древнее прочих, первичнее прочих — лобная доля, в частности префронтальный отдел. Этот вывод будет отвечать тезису, что у истоков второй сигнальной системы лежит не обмен информацией, т. е. не сообщение чего-либо от одного к другому, а особый род влияния одного индивида на действия другого — особое общение ещё до прибавки к нему функции сообщения[699].
Само разграничение этих двух сторон в человеческой речи уже не новость в советской психологической науке — новой является лишь задача определённо расположить во времени последовательность их возникновения. Вот что пишет А. Р. Лурия в работе «Регулирующая функция речи в её развитии и распаде»: наряду с «важнейшей» функцией речи — передачей информации — «существует и ещё одна её (речи) сторона, играющая столь же значительную роль в формировании сложных психических процессов. Речь не только служит средством общения и орудием кодирования полученного опыта. Она является одним из (?) наиболее существенных средств регуляции человеческого поведения…»[700]. Пусть не будут нижеследующие критические замечания поняты как недооценка огромной важности внесённого тут разграничения (тем более вообще вклада А. Р. Лурия в науку о работе мозга). Но во-первых, вопросительным знаком мною отмечена характерная и для нескольких других авторов предосторожность: как бы не оказаться в противоречии с какими-то философско-социологическими истинами, если сказать, что речь — единственное (а не «одно из наиболее существенных») средство регуляции человеческого поведения. Но в самом деле, вдумаемся: какие же ещё средства могут быть поставлены в тот же ряд? Если назовут «экономические отношения», «юридические нормы» и т. п. — здесь просто слово «регулятор» употребляется в другом смысле, как и если бы назвали «обмен веществ» и т. п. Если же укажут на другие бессловесные механизмы межиндивидуальных воздействий, а именно эмотивные, то они не более чем сопутствующие компоненты или дериваты речевого регулирования поведения. Наконец, уж и вовсе не убедительны указания на межиндивидуальные воздействия посредством безмолвного предоставления индивиду средств его деятельности, изготовленных другими и якобы передающих ему их опыт, или шире — исторический, социальный опыт: младенец якобы приобщается к обществу через пелёнки, соску, взрослый через инструменты, хотя бы никто не объяснял ему способа их употребления. Конечно же, ничего этого на деле не бывает: никто не суёт другому в руки новый инструмент без пояснений и показа (в последний тоже вовлечены слова «вот», «потом этак» и т. п.), что же до младенца, он в доречевом возрасте «социализируется» от употребления фабричных изделий ничуть не больше, чем пчёлы от пользования самыми модернизированными ульями с электрическими лампочками. Во-вторых, и это гораздо важнее, в приведённой формулировке А. Р. Лурия обе различаемые им функции речи рассматриваются только как одновременно сосуществующие в речевой деятельности современного человека, без попытки представить себе, что вторая, выделенная здесь, можно сказать открытая Выготским и им, регулирующая функция существовала некогда сама по себе в чистом виде, до того, как в эволюции человека к ней присоединилась или над ней надстроилась функция информации, обмена опытом[701]. Но дальнейшие успехи в изучении нейропсихологии речевой деятельности возможны только посредством генетического расчленения её на разные ступени. Только когда мы выделим не просто «играющую значительную роль» функцию, но регулятивную или инфлюативную фазу в теории возникновения второй сигнальной системы, мы выйдем на дальнейшую широкую научную дорогу.
Слабой стороной столь сильной школы А. Р. Лурия, школы нейропсихологии, слабой стороной всего быстро развивающегося учения о локализации корковых функций, в частности и в особенности о функциях лобных долей, по моему убеждению, является «неисторичность»[702]. Не следовало бы располагать в одной плоскости характеристику работы лобных долей животных и человека, ибо, повторяю, у животных даже и чисто морфологически лобные доли не имеют того самого существенного — верхних переднелобных формаций, что характеризует их у человека. Не следовало бы и употреблять одни и те же понятия «задача», «намерение» для определения воздействия лобных долей на поведение и животных, и человека, ибо «задача» существует в голове экспериментатора, а не животного, у последнего же, как говорилось выше, никаких «моделей потребного будущего», или «целей», «задач», «намерений» и в помине нет: это продукты второй сигнальной системы. Животные, даже в стаде, являются прежде всего индивидуальными организмами. Человек, даже в одиночестве, является прежде всего носителем второсигнальных воздействий и тем самым социального опыта и социальной истории.
В наибольшей степени «неисторичность» нейропсихологии проявляется в исследовании именно мозговых зон и механизмов человеческой речевой деятельности. Словно она и возникла, и отродясь существовала в её нынешнем виде, как сложенное из многих взаимосвязанных компонентов целое. Это оправдано тем, что главным помощником и посредником в этой работе является клиника — изучение и лечение патологии речи центрального, коркового происхождения: какое дело врачу до того, какие элементы речевой функции современного человека вышли на сцену раньше или позже в эволюционном формировании людей? Для него важно лишь то, что нарушение того или иного компонента речевой деятельности может служить диагностическим признаком поражения (опухоль, инсульт, травма и т. д.) того или иного определённого участка коры левого полушария (у правшей) головного мозга. Соответствующая отрасль невропатологии, афазиология, в последнее время достигла чрезвычайных успехов. Мозговая локализация (корковая топика или топография) различных форм патологии речи (афазий), а тем самым и выявление зон коры, управляющих в норме разными элементами, из которых слагается импрессивная и экспрессивная речь, — всё это продвинуто довольно далеко[703].
Но в какой исторической последовательности эти зоны появлялись как опоры разных фаз развития второй сигнальной системы? Для ответа на этот вопрос может служить не только эволюционная морфология мозга по ископаемым черепам (ибо «речевые зоны» достаточно анатомически выражены и обозримы, чтобы их наличие, отсутствие, степень выраженности отразились на эндокранах этих ископаемых черепов), а и данные самой афазиологии.
Для этого надо взглянуть на клинические наблюдения под пока ещё необычным для афазиологов углом зрения. Больному с афазией врач или дефектолог задаёт вопрос, даёт задачу, инструкцию и по ответам или ответным действиям больного определяет характер, форму, как и степень, глубину нарушения восприятия речи или высказывания. Произошла такая-то поломка в механизмах слушания и усвоения чужой речи, в механизмах говорения. При этом афазиолог в своих выводах элиминирует самого себя, соучастие второго лица в данном речевом общении; но закономерна гипотеза, что на самом деле поломка относится как раз к реакциям на вопросы, обращения, сообщения, задания. Дело не в том, что больной не может обращаться как надо со слышимыми или произносимыми им словами, а в том, что он не может обращаться как надо с партнёром — реагирует при поражениях соответствующих участков коры по эволюционно архаическим, т. е. отменённым и заторможенным с развитием коры, схемам. Иными словами, афазии не просто поломки, т. е. не просто уничтожение чего-то, но они возбуждение чего-то, а именно возбуждение некоторых мозговых механизмов, которые в норме (вне поломки) подавлены, не возбуждаются, «загнаны вглубь», перекрыты более молодыми образованиями. Тот факт, что больной с афазией обычно ясно сознаёт свой дефект, нимало не противоречит этой гипотезе. Больной сохраняет сознание и самоконтроль современного человека, он вовсе не «хочет» реагировать на современную речь, скажем, как кроманьонец, но замечает, что болен, что не может реагировать правильно. А афазиолог недостаточно учитывает, что поражение того или иного участка коры головного мозга вызвало нарушение отношений и взаимодействий больного с людьми, регресс вниз на ту или иную древнюю ступеньку второй сигнальной системы, когда она ещё не была или не вполне была номинативно-информативной, а выполняла другие задачи.
В норме генетически позднейшие слои и образования коры подавляют или преобразовывают эти ранние функциональные системы, обнаруживающиеся ясно и изолированно только в патологии. С развитием вида Homo sapiens происходило морфологическое и функциональное подтягивание более начальных второсигнальных зон и систем к уровню позднейших. Ведь современный ребёнок должен включаться сразу в нынешнюю систему речевой коммуникации. Развитие его мозга в этом смысле лишь в малой мере повторяет филогенез, в большей же мере разрабатывает врождённое устройство, обеспечивающее усвоение языка современного человеческого типа[704]. А филогенетически исходные типы второсигнальных реакций у современного человека подавлены, на ребёнке они наблюдаемы лишь отчасти, в общем же обнажаются лишь в патологии.
Если это так (а клинические наблюдения автора, как и критический анализ литературных данных, заставляют так думать), афазиологии суждено стать бесценным источником научных знаний о пути, генетически пройденном второй сигнальной системой. Первую часть этого пути составляла фаза, когда она, собственно, не была средством «отражения» чего-либо из предметной среды, а была реагированием лишь на специфические воздействия людей, причём автоматическим (роковым), как, скажем, взаимосвязь органов внутри организма, т. е. принадлежала к бытию, а не к сознанию или познанию. Главы, посвящённые тормозной доминанте, имитации и интердикции, уже подготовили читателя к пониманию того, о чём мы говорим. Вторая сигнальная система родилась на фундаменте интердикции. Это был объективный механизм межиндивидуального воздействия на поведение. Дело не меняется от того, что теперь перед нами явление «интердикция интердикции» — свойственные Homo sapiens механизмы парирования интердикции.
И. П. Павлов не успел познать весь скрытый потенциал своей великой научной идеи о двух сигнальных системах у человека. Он шёл от того, что открыл в высшей нервной деятельности животных. Он писал: «Слово для человека есть такой же реальный условный раздражитель, как и все остальные, общие у него с животными, но вместе с тем и такой многообъемлющий, как никакие другие», ибо «слово благодаря всей предшествующей жизни взрослого человека связано со всеми внешними и внутренними раздражителями, приходящими в большие полушария (всех их сигнализирует, всех их заменяет), и может вызвать все те действия, реакции организма, которые обусловливают те раздражения»[705]. Ныне наука вправе пойти дальше в суть дела. Слово только ли заменяет и сигнализирует «все» раздражители, т. е. только ли изоморфно им, или оно делает ещё что-то, чего они не делают и чего в них нет? Простейшая иллюстрация: разве комбинации слов не производят постоянно и объекты, которых нет в мире реальных раздражителей, но которые становятся образами, а часть которых позже воплощается в реальность средствами искусства и техники? А что значит при ближайшем рассмотрении выражение, что слово «заменяет» все внешние и внутренние раздражители: не ясно ли, что, прежде чем «заменять», слово должно было освобождать место для замены, т. е. «отменять» те реакции, те действия организма, которые прежде вызывались этими раздражителями, т. е. тормозить их? Между тем физиологи, исследовавшие вторую сигнальную систему человека, увязли в простых параллелях между словами и первосигнальными раздражителями. Их справедливо критикует Н. И. Чуприкова: «Интересы даже тех исследователей, которые занимаются изучением работы мозга человека, до сих пор в значительной степени сосредоточены не на тех закономерностях, которые отличают высшую нервную деятельность человека от высшей нервной деятельности животных, а на нервных явлениях, в равной мере свойственных животным и человеку»[706].
Но когда речь идёт о генезисе этих человеческих закономерностей, даже говорить об их «отличии» от высшей нервной деятельности животных было бы недостаточно и неопределённо. На деле то было появлением врага, противника у первой сигнальной системы. Организм стал производить действия, не диктуемые его собственной сенсорной сферой. Следовательно, он не стал производить действий, диктуемых этой его собственной сенсорной сферой. Последние в этот момент подавлены, поражены.
Долго, очень долго вторая сигнальная система была всего лишь таким фактором, управляющим некоторыми действиями, целыми цепями действий, вторгаясь там и тут в поведение ранних людей. Она отвоевывала всё более обширные поля у первосигнальной детерминации поведения. А неизмеримо позже она приобрела знаковую функцию, слова и системы слов стали нечто означать и значить, в том числе «заменять» первосигнальные раздражители.
Но мало расчленить историю второй сигнальной системы на эти два столь глубоко различных этапа. Хоть такое расчленение и важно и трудно, всё же самое трудное исследовать и объяснить, как же именно одно явление превратилось в другое. Этим мы и должны заняться ниже.
Сначала подведём итог сказанному. У порога истории мы находим не «надбавку» к первой сигнальной системе, а средство парирования и торможения её импульсов. Только позже это станет «надбавкой», т. е. отрицанием отрицания. Важным шагом к такому преобразованию служит превращение интердикции в суггестию, хотя последняя и лежит ещё в рамках инфлюативного этапа второй сигнальной системы. Суггестия становится фундаментальным средством воздействия людей на поступки и поведение других, т. е. особой системой сигнальной регуляции поведения.
Эта нейрофизиологическая система взаимного оттормаживания и побуждения тех или иных действий предшествует возможности возникновения общественных отношений и общества, но в то же время может рассматриваться как первичная завязь общественных отношений.
Как мы выше констатировали, суть этой системы в том, что она побуждает индивида делать что-либо, что не диктуется собственными сенсорными импульсами его организма. Причём она явно и далеко выходит за пределы имитативного побуждения, присущего и животным. В этом смысле она уже антибиологична.
Вот каков корень у закономерности экономического поведения первобытных людей. Как видим, он уходит в наидревнейшую глубину истории. Мы находим там не деятельность одиночек, оббивающих камни, которой так злоупотребляют многие авторы для объяснения начала истории, а прежде всего отношения людей — отношения столь непохожие на те, какие кажутся единственно нормальными этим авторам. История первобытного общества в его нормах, обычаях несла ещё долгие последствия указанной начальной системы человеческих взаимодействий — даже тогда, когда появились и развились следующие, более поздние формы речевого общения. То были антибиологические отношения и нормы — отдавать, расточать блага, которые инстинкты и первосигнальные раздражители требовали бы потребить самому, максимум — отдать своим детёнышам либо самкам.
Остаётся повторить, что такой порядок вещей требовал необходимого корректива — распадения человечества на великое множество общностей с разными искусственными признаками их обособления и культурами, которые ставили предел отчуждению благ «вовне». Эти признаки одновременно и отмежёвывали общность и сплачивали её[707]. Для нашей проблемы особенно важно, что тем самым мы должны предполагать на границах первобытных общностей какое-то нарушение механизма суггестии, возникновение чего-то ей препятствующего, что можно уже отнести к явлениям «непонимания» в широком смысле.
II. Суггестия
В главах 4 и 5 мы рассмотрели физиологические явления тормозной доминанты и имитативного рефлекса и показали, что их встреча в высшей нервной деятельности семейства троглодитид должна была породить высоко развитое явление интердикции. Наибольшую ступень интердикции мы назвали генерализованной интердикцией: имитационное провоцирование некоторого одного действия парализует возможность каких бы то ни было других действий (очевидно, за исключением функционирования автономной нервной системы) и это состояние парализованности, вероятно, может продолжаться долго — даже после прекращения действия данного имитатогенного агента.
Тем самым высшую форму интердикции можно было бы считать низшей формой суггестии: это уже не торможение лишь того или иного отдельного действия, но навязывание некоего состояния, допустим, типа каталепсии. Однако таков лишь зачаток суггестии, ибо под ней (под «внушением») понимается возможность навязывать многообразные и в пределе даже любые действия. Последнее предполагает возможность их обозначать. Между этими предельными рубежами умещается (развивается) явление суггестии. Нас прежде всего интересуют его первые шаги.
Надо напомнить ещё раз коренной тезис, что вначале, в истоке, вторая сигнальная система находилась к первой сигнальной системе в полном функциональном и биологическом антагонизме. Перед нашим мысленным взором отнюдь не «добрые дикари», которые добровольно подавляют в себе вожделения и потребности для блага другого: они обращаются друг к другу средствами инфлюации, к каковым принадлежит и суггестия, для того чтобы подавлять у другого биологически полезную тому информацию, идущую по первой сигнальной системе, и заменять её побуждениями, полезными себе. Это явление инфлюации, в том числе суггестии, не имеет никакого отношения к гносеологии. Сколько обвинений в идеализме и мистицизме было обрушено на вывод Леви-Брюля о «прелогическом мышлении», тогда как это явление действительно существовало на заре истории и проявлялось впоследствии, но только оно не было «мышлением»: оно было подавлением первосигнального (ещё единственно верного тогда) способа отражения окружающей среды и системой принудительного воздействия на поведение друг друга. Да, оно принадлежит не к гносеологии, а к онтологии. Это только взаимодействие особей. Тут нет отношения субъекта к объекту, а есть лишь отношение организма к организму. «Прелогическая стадия» ничуть не угрожает логике: тут логике ещё решительно нечего было делать. Мы убедимся, что и антилогика в самом деле гораздо раньше объявилась в этой сфере отношений между индивидами, чем логика в сфере человеческого познания. Но о генезисе логики мы будем говорить только в конце этой главы (и, тем самым, этой книги).
Развитие второй сигнальной системы у людей ни в коем случае не было следствием разрастания общего объёма головного мозга по сравнению с объёмом (весом) тела и прямо никак не связано с этим процессом энцефализации (церебрализации) в филогении троглодитид. Во-первых, в биологической эволюции вообще налицо тенденция увеличения мозга (независимо от размеров тела), и по этому показателю прямоходящие высшие приматы, включая человека, вовсе не оказываются специфичными: степень энцефализации увеличивается у всех ископаемых млекопитающих. По степени развития неокортекса («новой коры»), т. е. по степени «неоэнцефализации», можно выделить весь отряд приматов из общей родословной млекопитающих, но невозможно отдифференцировать собственно человеческую линию[708]. Во-вторых, разрастание объёма головного мозга в эволюции семейства троглодитид было прямым морфологическим следствием прямохождения и прямым биохимическим следствием плотоядения, т. е. повышенного усвоения протеина[709]; эти два фактора влекли за собой широкий размах индивидуальных вариаций объёма мозга, иначе говоря, делали структурно и гистологически возможными колебания в сторону повышенной массы мозга, а естественный отбор закреплял эти отклонения, несомненно, потому, что они были биологически выгодны. По Боне, это разрастание мозга происходило в том же темпе, как и другие морфологические трансформации млекопитающих в плейстоцене. Но у Homo sapiens средний размер и вес головного мозга не возрастал и не возрастает сравнительно с поздними палеоантропами. При этом размах индивидуальных вариаций данного признака у Homo sapiens весьма увеличился сравнительно с палеоантропами, нередко встречается объём мозга, значительно превышающий среднюю величину, но ничто не закрепляет этих отклонений: они биологически нейтральны и средняя величина остаётся неподвижной для черепов любого времени с верхнего палеолита до наших дней. Между тем речевая функция мозга в корне отличает неоантропа от палеоантропа.
Как видно, социальность и разум человека никак прямо не коррелированы с тотальной величиной его головного мозга.
Зато корреляция начинает проступать, когда измерению подвергаются по отдельности длиннотный, широтный и высотный диаметры роста головного мозга в филогенетической цепи: шимпанзе — австралопитек — археоантроп — палеоантроп — неоантроп. До неоантропов наиболее интенсивно увеличивался тотальный длиннотный размер (за счёт роста задних областей); напротив, только мозг неоантропов даёт интенсивный сдвиг высотного диаметра: в лобной и теменной долях эпицентры роста перемещаются из нижних отделов в верхние, происходит, как мы уже отмечали, усиленный рост префронтальной области, а также поднятие и выравнивание поверхности мозга в его своде[710].
Интересно, что как раз в самой высотной, и тем самым весьма молодой, точке свода нашего мозга предположительно локализуется самый исходный, «инициальный» очажок речевой функции человека (см. схему коры мозга). Правда, вопрос пока является дискуссионным и открытым. Это так называемая «речевая зона Пенфильда», или «верхняя речевая кора», — якобы зона начальной речевой активизации[711]. Эту зону Пенфильд и Робертс якобы обнаружили при оперативных исследованиях эпилепсии в так называемом дополнительном моторном поле на медиальной (внутренней, т. е. обращённой к другому полушарию) поверхности в задних отделах верхней лобной извилины. Некоторые авторы считают это прочно установленным фактом[712], другие при применении иных нейрохирургических методов не обнаруживают нарушений речевых функций при поражениях этой зоны (А. Р. Лурия) или замечают воздействие всего лишь на оттенки интонации и модуляции. Впрочем, последнее не служило бы решающим возражением: кто знает, может быть, так и рождалась первичная дифференциация звуков-знаков. Не будем пока ни принимать, ни отбрасывать гипотезу Пенфильда — Робертса. Заметим лишь, что, если бы указанная зона в самом деле всё-таки оказалась начальным очажком второй сигнальной системы, было бы поразительно, что он расположен в самой высотной части, т. е. на самом молодом верхнем крае коры мозга Homo sapiens. В пользу второсигнальной принадлежности этого образования (если принять описание его Пенфильдом и Робертсом) говорит его односторонняя локализованность в доминантном полушарии (в левом у правшей), тогда как близко расположенные центры локализации находятся в обоих полушариях.
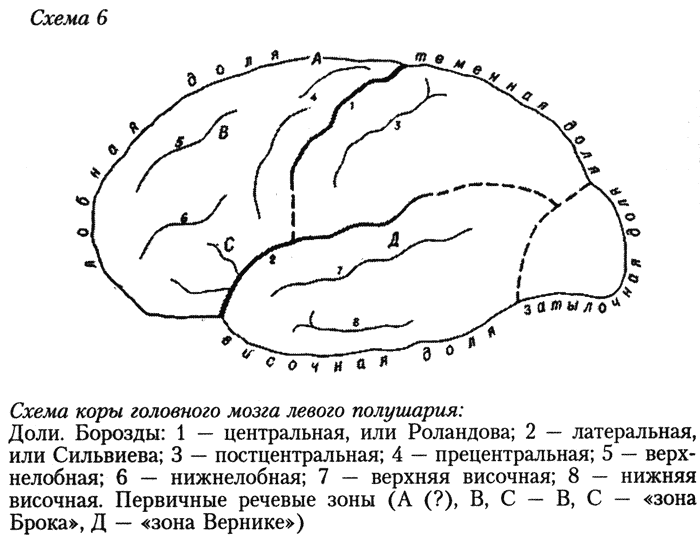
Интересно, что довольно высоко в структуре коры головного мозга Homo sapiens, впереди от средней части прецентральной извилины, лежит и участок, управляющий элементарными графическими действиями. Это свидетельствует, что они возникают на весьма ранних стадиях генезиса второй сигнальной системы. Люди начальной поры верхнего палеолита уже «рисовали», ещё не вполне умея «разговаривать», если применять эти термины в современном смысле. Туда, в эту древность, уходят корни письменной речи. Зато когда иные археологи фантазируют, будто они обнаружили «рисунки» мустьерцев (палеоантропов, неандертальцев), мы уверенно можем считать это исключенным в той же мере, как заявку на изобретение перпетуум-мобиле: в мозге палеоантропов ещё не было той высотной части, где находится центр, управляющий графическими действиями; они не могли носить и неуправляемого двигательно-подражательного характера (какие осуществляют шимпанзе), так как подражать в «рисовании» ещё было тогда некому.
Ещё далее вперёд от самой высотной части свода головного мозга Homo sapiens (где сходятся лобная и теменная доли) лежит тот префронтальный отдел, о роли которого, в особенности его верхней формации, мы уже говорили в связи с постановкой проблемы происхождения второй сигнальной системы. Это образование присуще только Homo sapiens.
Можно сказать, что это главное морфологическое звено второй сигнальной системы. Отсюда по исключительно богатым нервным путям и контактам происходит возбуждение, активация (с помощью сетевидной, или ретикулярной, формации) отвечающих «задаче» или «намерению» двигательных центров мозга и одновременное торможение всех других двигательных центров, не идущих к делу[713].
Эволюционная морфология мозга не может ничего прямо сказать о развитии функциональной асимметрии полушарий (доминантность — субдоминантность, правшество — левшество). Вообще говоря, асимметрия как один из аспектов парности работы полушарий головного мозга обнаружена у животных: симметричные центры двух полушарий могут в данный момент находиться во взаимном антагонизме, т. е. в одном полушарии выполнять функцию возбуждения, во втором — торможения[714]. Но у человека асимметрия закреплена: лишь кора одного полушария, обычно левого (у правшей), управляет всей второсигнальной функцией. Афазиологией доказана левосторонность управления динамикой как речевой, так и рече-мыслительной деятельности в лобных долях. Однако пока недостаточно подчёркивается, что вся эта односторонность управления второй сигнальной системой, т. е. локализация всех прямо причастных к ней зон и центров в «доминантном» полушарии, означает сопряжённое торможение центров, управляющих неречевыми движениями — преимущественно с противоположного, «субдоминантного» полушария, но отчасти с того же[715]. Если так, можно существенно переакцентировать обычное восприятие понятий «правшество», «левшество»: дело не в том, что у «правши» правая рука (и другие органы) обладает некими повышенными возможностями, а в том, что у него, напротив, приторможена, снижена левосторонняя моторика (в первую очередь руки), у «левши» — наоборот. Есть немало оснований ограничить недавнее слишком абсолютное представление о двигательном «правшестве» и «левшестве»: в большинстве случаев то и другое как-то смешано в индивиде. Но по некоторым данным онтогенеза можно предполагать, что в филогенезе асимметрия прошла три фазы: правшество — левшество — снова правшество; поэтому их следы у нас могут наслаиваться друг на друга.
Что же мы обнаружили, выясняя древнейшие уровни, «исходные рубежи» второй сигнальной системы с помощью эволюционной морфологии мозга, афазиологии, генетической психологии? Мы обнаружили, что древнейшие зоны речевой деятельности возникают в моторной (двигательной), а не сенсорной (чувствующей) области коры. Это и отвечает выдвинутому выше тезису, что вторая сигнальная система родилась как система принуждения между индивидами: чего не делать; что делать. Мы получим дальнейшее подкрепление этого тезиса, перейдя от тотальных параметров, характеризующих особенности головного мозга Homo sapiens, к эволюционному сравнению состояния отдельных долей, извилин и борозд, поскольку они отразились на эндокранах ископаемых предковых форм, и к частным видам афазий.
Подчас антропологи принимают намеченное на эндокране питекантропа образование на месте так называемой «зоны Брока» в нижней лобной извилине за свидетельство присутствия у него речи, исходя из того, что у современного человека повреждения этой зоны вызывают нарушение двигательной (произносительной) речевой функции — моторную афазию. Со своей стороны, повреждения «зоны Вернике» в первой височной извилине вызывают сенсорную афазию — нарушение восприятия чужой слышимой речи. Но речь не просто наличие двух устройств — передающего сигналы и принимающего их, тем менее — одного из двух; недостаточно и двусторонней связи между ними, т. е. единства слухо-двигательного анализатора. Допустим, питекантропы могли издавать и принимать разные крики; это не членораздельность, не речевые знаки в смысле современной науки. В. В. Бунак на основе анатомии периферических органов речи — гортани, челюстного скелета — установил, что у форм, предшествовавших Homo sapiens, не могло быть членораздельной синтагмической речи[716] (дедукция какой-то якобы предшествовавшей, «нечленораздельной речи» не удалась — она противоречит всей аксиоматике науки о речи); в 1971 г. появилось сообщение, что Ф. Либерман (Ун-т Коннектикут) изготовил искусственный макет гортани неандертальца (шапелльца) и получил экспериментальное физическое подтверждение того, что она не могла бы производить членораздельных звуков[717].
Однако суть дела вовсе не в этой физико-акустической стороне проблемы. Она попала в центр внимания только тех авторов, которые не в курсе современной лингвистики, как и афазиологии. Поражение «зоны Вернике» в височной доле вызывает не потерю слуха или возможности различать звуки, нарушает не акустическую основу восприятия речи, словом, порождает не «фонетическую глухоту», но «фонологическую глухоту» — невозможность правильно узнавать фонемы как смыслоразличающие элементы речи. Наука фонология, отделившаяся от фонетики (за которой осталась физиология звукообразования и звуковосприятия), изучает наличные во всяком языке звуковые «букеты», которые психика человека расценивает как один и тот же элемент, сколь бы ни отличались друг от друга эти звуки в плане акустическом, и которые имеют и сохраняют свою определённость только по противопоставлению другому такого же рода «букету» в той же артикуляционно-фонетической группе[718]. Именно в этом проявляется тут «чрезвычайная прибавка» человека — в возможности отождествлять звуки, высоко различные акустически (ср., например, весьма искажённое «повторение» маленьким ребёнком слов взрослого), как и, напротив, в возможности делать эти звуки принципиально нетождественными даже при их немалой акустической близости — посредством оппозиции, по природе столь же абсолютной, как и нейрофизиологические явления возбуждения и торможения.
Что касается не восприятия речи, а говорения, то оказалось, что и двигательная (моторная) сторона речевой деятельности раздвоена точно так же. Поражения управляющих ею зон в коре мозга, в том числе нижней лобной извилины, нижнего отдела прецентральной извилины, нижнего отдела теменной доли, приводят либо к эфферентной моторной афазии — нарушению функции противоположной фонематической, а именно функции не разделения фонем, а их связности, плавности, слияния хотя бы и различных звуковых единиц в одну единицу, например в слог; либо к афферентной моторной афазии — смешению, неразличению фонем, в частности, близких по артикуляции. В последнем случае перед нами патология не самой артикуляции звуков органами речи, а психо-физиологического механизма контроля этих движений по фонологическим меркам, иначе говоря, психо-физиологического механизма обратной коррекции речевых движений[719].
Выходит, что «сенсорная афазия» и «моторная афазия» представляют собою два свидетельства одного и того же факта — появления в мозге Homo sapiens на определённом этапе его формирования принципиально нового уровня реагирования. Важно, что оба они и в анатомо-физиологическом смысле тесно соединены. Нижние отделы лобной и теменной долей (очаги моторных афазий) близко примыкают к переднему отделу височной доли (очагу сенсорной афазии). Только будучи по существу единым аппаратом, они могут осуществлять эхолалическую (речеподражательную) операцию, лежащую глубоко в основе всей нашей речевой, а тем самым и речемыслительной деятельности: непроизвольное повторение слышимого, причём не на акустико-фонетическом (не как у попугаев или скворцов), а именно на фонологическом уровне. В норме эта операция у нас редуцирована, так что зарегистрировать её могут только тончайшие электрофизиологические приборы[720], но при поражениях или функциональных расстройствах в коре, т. е. когда подавляющие её позднейшие нервные образования вышли из строя, она выступает с полной наглядностью и назойливостью. Врач говорит больному «встаньте», тот повторяет «встаньте», но не встаёт. Эхолалическая реакция на речь не несёт никакой смысловой нагрузки. Показано, что это явление характерно для ранней стадии освоения речи ребёнком младшего возраста, а также для различных нервных расстройств (неврозов), в том числе истерии[721]. Тот факт, что этот фундаментальный механизм речевой деятельности протекает у нас необычайно быстро, следовательно, по простейшим нейронным путям, свидетельствует о его особой древности, в некотором смысле даже первичности в эволюционном становлении второй сигнальной системы у неоантропов.
Этому соответствует указанная анатомическая связь органов, или зон фонологического анализа и контроля, в коре, в частности близость и взаимосвязь нижнелобной и височной долей. Если у австралопитеков лобная и височная доли плотно примыкали друг к другу, то у синантропов и палеоантропов они были резко разделены довольно широкой, глубокой и узкой сильвиевой ямкой, а у неоантропов края её снова соединились, однако переместившись вперёд и при новых очертаниях всей височной доли[722]. Современные знания о работе, выполняемой у нас U-образным изгибом коры в глубине височной ямки, недостаточны для интерпретации этого крутого морфологического преобразования. Однако можно с известной долей уверенности предполагать, что указанный изгиб осуществляет прямую и кратчайшую нейронную связь между «зоной Брока» и «зоной Вернике» и тем самым обеспечивает эхолалическую (речеподражательную) подоснову второй сигнальной системы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Инерция мышления
Инерция мышления «Размышляя о возможном, люди пользуются примерами прошлого и предвосхищают будущее с воображением, занятым прошедшим. Этот путь рассуждений часто является ошибочным, так как реки, вытекающие из истоков природы, не всегда укладываются в старые
Глава 14 Homo erectus. Развитие мозга. Зарождение речи. Интонации. Речевые центры. Глупость и ум. Смех—плач, их происхождение. Обмен информацией в группе.
Глава 14 Homo erectus. Развитие мозга. Зарождение речи. Интонации. Речевые центры. Глупость и ум. Смех—плач, их происхождение. Обмен информацией в группе. Homo erectus оказался весьма пластичным «прачеловеком»: за миллион с лишним лет своего существования он все время
Глава 17 Гибкое тело человека. Особые свойства человеческого мозга. Значение образного мышления. Пещерные рисунки. «Сверхтучная» женщина. Был ли кроманьонец обнаженным? Появление религии. Шаманы и служители божьи.
Глава 17 Гибкое тело человека. Особые свойства человеческого мозга. Значение образного мышления. Пещерные рисунки. «Сверхтучная» женщина. Был ли кроманьонец обнаженным? Появление религии. Шаманы и служители божьи. Ясно, что наш вид Homo sapiens sapiens неизмеримо превосходил все
§ 17. Признаки мышления
§ 17. Признаки мышления 1. Любые генетически детерминированные формы поведения не могут считаться мышлением. Такая детерминация может иметь как прямой генетический характер, так и косвенные формы. Генетически детерминированные формы поведения могут реализовываться
§ 18. Биологические проблемы мышления
§ 18. Биологические проблемы мышления Обладатели крупного мозга становятся своеобразными заложниками его размеров. Нравится или не нравится животному, но в большом количестве нейрональных связей постоянно фиксируется повседневная информация о внешнем мире и
Глава 3 Феномен человеческой речи
Глава 3 Феномен человеческой речи I. О речевых знакахВозникновение человеческой речи — одновременно и «вечная проблема» науки, и проблема всегда остававшаяся в стороне от науки: на базе лингвистики, психологии, археологии, этнографии, тем более спекулятивных рассуждений
9.6. Функциональная асимметрия мозга и нейрофизиология речи
9.6. Функциональная асимметрия мозга и нейрофизиология речи После фундаментальных работ выдающегося нейрофизиолога Р. Сперри (1913–1994) понятие функциональных различий полушарий человеческого мозга стало общепринятым (Sperry R., 1969). Функциональная асимметрия между двумя
Феномен человеческой речи
Феномен человеческой речи "Язык выделил человека из всего животного мира" — пишет Дж. Бернал, и это бесспорно. Но что же является главной характеристикой языка, речи? Специфическое и самое существенное свойство человеческой речи — наличие для всякого обозначаемого
3.2. Механизмы мышления
3.2. Механизмы мышления Ответу на этот вопрос были посвящены наши исследования последних десяти лет. В работе ставилась задача изучить структуру корковых связей при решении различных мыслительных операций. При этом мы исходили из фундаментальных идей российской
Создание орудий мышления
Создание орудий мышления Как нельзя голыми руками заниматься плотницким делом, так и мышление редко может осуществляться с помощью голого мозга Бо Дальбом, Ларс-Эрик Джанлерт. Компьютерное будущее (в печати). Перед каждым агентом стоит задача наилучшего использования
Формы мышления
Формы мышления На основе многолетних экспериментальных исследований Ладыгина-Котс пришла к выводу, что мышление животных имеет всегда конкретный чувственно-двигательный характер, что это мышление в действии, причем эти действия являются всегда предметно-отнесенными.
Запечатление речи
Запечатление речи Многих всегда поражал парадокс: речь, самое сложное и совершенное из всего, на что способен человек, ребенок осваивает в столь раннем возрасте, в котором во всем другом мало на что способен, несмышленыш.Сравнительно недавно этологи нашли разгадку этого
Глава 7 ГОВОРЯЩИЕ ГОЛОВЫ[94] Как мы понимаем язык и используем его в речи
Глава 7 ГОВОРЯЩИЕ ГОЛОВЫ[94] Как мы понимаем язык и используем его в речи На протяжении столетий людей ужасала возможность того, что созданные ими вычислительные машины могут перехитрить их, в чем-то превзойти или лишить рабочих мест. Этот страх часто обыгрывался в