Сравнительная геномика и ранняя эволюция животных[61]
Сравнительная геномика и ранняя эволюция животных[61]
Если не учитывать редкую и плохо изученную мелочь вроде трихоплакса, то самыми примитивными животными можно назвать губок, у которых еще нет настоящих тканей, нервной системы и кишечника. Губки противопоставляются всем прочим животным — так называемым «настоящим многоклеточным животным» (Eumetazoa). Среди этих последних самыми примитивными считаются кишечнополостные, или книдарии (к ним относятся медузы, актинии, гидроидные и коралловые полипы), и гребневики. В пределах Eumetazoa книдарии и гребневики («радиально-симметричные животные») противопоставляются двусторонне-симметричным (Bilateria). К двусторонне-симметричным относятся все прочие животные от жуков и червей до морских звезд и человека.
Генетические исследования последних лет показали, что традиционные представления о примитивности книдарий, скорее всего, сильно преувеличены. В частности, оказалось, что у них, как и у высших животных, есть Hox-гены, которые в ходе индивидуального развития задают полярность зародыша и определяют план строения, в котором явно проступают черты билатеральной симметрии. Это подтвердило старую гипотезу, согласно которой общий предок Eumetazoa был двусторонне-симметричным животным. Для того чтобы составить более полное представление об этом предке, было необходимо прочесть геном представителя книдарий и сравнить его с известными геномами билатерий.
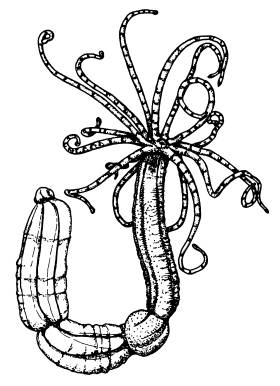
Актиния Nematostella.
И вот в 2007 году американские ученые сообщили о «черновом» прочтении генома актинии Nematostella[62]. Геном состоит из 15 пар хромосом, имеет размер около 450 млн пар нуклеотидов (в 100 раз больше, чем у кишечной палочки, и в 6 раз меньше, чем у человека) и содержит примерно 18 000 белок-кодирующих генов, что вполне сопоставимо с другими животными.
Для каждого гена актинии исследователи пытались найти аналоги в геномах билатерий: человека, дрозофилы, круглого червя, рыбы и лягушки. Если аналог (то есть похожий ген) находился, исследователи делали вывод, что соответствующий ген имелся у общего предка Eumetazoa. Таким образом удалось составить довольно полное представление о генном репертуаре этого загадочного предка.
Оказалось, что репертуар этот был весьма широк и включал не менее 7766 генных семейств, сохранившихся и у книдарий, и у билатерий. Человек унаследовал не менее 2/3 своих генов от общего с актинией предка; сама актиния — примерно столько же. Муха и круглый червь унаследовали от общего предка с актинией лишь 50% и 40% генов соответственно.
Выяснилось, что в эволюционных линиях позвоночных и книдарий было потеряно меньше исходных генов и меньше приобретено новых, чем в линии, ведущей к круглым червям и насекомым. Однако нужно иметь в виду, что применяемые методики не позволяли отличить действительную потерю гена от его изменения «до неузнаваемости». Поэтому в целом можно лишь заключить, что в линии позвоночных геном изменился меньше, чем в линии первичноротых, куда относятся муха и червь. Из этого следует неожиданный вывод: геном человека оказался в целом гораздо более похожим на геном актинии, чем геномы мухи и червя. Сходство затрагивает не только набор генов, но и порядок их расположения в хромосомах.
Около 80% генов общего предка Eumetazoa имеют явные аналоги за пределами животного царства. Это значит, что они были унаследованы животными от одноклеточных предков (как мы помним, ими были воротничковые жгутиконосцы — хоанофлагелляты). Получается, что геном на удивление мало изменился при становлении животного царства. Среди оставшихся 20% генов, аналогов которых нет у одноклеточных, имеется большое число ключевых регуляторов развития. Примерно четверть этих новых генов (то есть 5% от общего числа) содержат участки или функциональные блоки, встречающиеся у одноклеточных, но в других комбинациях. Это указывает на один из основных путей создания новых генов: они формируются из старых путем перекомбинирования фрагментов.
Как и следовало ожидать, значительная часть «новых» генов Eumetazoa выполняет функции, непосредственно связанные с теми новшествами, которые появились у животных на организменном уровне. Это прежде всего гены, отвечающие за межклеточные взаимодействия и передачу сигналов, за перемещения клеток, регуляцию их деления и другие процессы, играющие ключевую роль в ходе индивидуального развития животных.
—————
Hox-гены — регуляторы развития животных[63]. Ключевую роль в эволюции животных играли изменения сравнительно небольшого числа генов — регуляторов индивидуального развития, от которых зависит план строения организма. Главными из этих генов-регуляторов являются так называемые Hox-гены.
Прошло уже больше 50 лет с тех пор, как американский генетик Эдвард Льюис с удивлением рассматривал мутантную плодовую мушку, у которой на голове вместо антенн выросли ноги. Эта странная мутация получилась, когда ген, ответственный за формирование грудных конечностей, включился не в то время и не в том месте. А Эдвард Льюис (вместе с Кристианой Нюссляйн-Волхард и Эриком Вишаусом) получил в 1995 году за исследование этих механизмов эмбриогенеза Нобелевскую премию по физиологии и медицине.
Так было открыто семейство Hox-генов, отвечающих за правильное формирование частей тела у животных. Работа этих генов казалась чудом: вот оно, решение великой загадки, как из сборища одинаковых эмбриональных клеток в правильном порядке дифференцируются ткани и органы и в результате получается сложный организм. Необходимо только в нужный момент включать правильный Hox-ген.
Регуляторные Hox-гены у дрозофилы располагаются в хромосоме в довольно строгом порядке, приблизительно в том самом, в котором происходит дифференцировка основных частей тела двусторонне-симметричного (билатерального) животного. Сначала у раннего эмбриона начинают работать гены, отвечающие за строение органов на голове, затем на груди, затем гены начинают оформлять и хвостовую часть.
Похожие гены были найдены у мыши и у человека. Даже у этих высокоорганизованных существ они выполняют ту же работу: отвечают за порядок эмбрионального развития. Открытие сходных Hox-генов у разных типов животных заставило зоологов и эмбриологов по-новому взглянуть на индивидуальное развитие животных и его преобразования в ходе эволюции. Стало ясно, что, изменив всего лишь один ген или время его включения, можно трансформировать, дублировать, удалить или перенести в другое место сразу целый орган, сохранив при этом общий план строения.
Кроме того, ученые получили новый мощный инструмент для эволюционных построений — семейство гомологичных (происходящих один от другого) генов, присутствующих у всех животных. Все гипотезы о происхождении и ранней эволюции животных теперь включают и этот пласт информации.
Hox-гены располагаются на одной или нескольких (до четырех) хромосомах, обычно тесными группами (кластерами), внутри которых сохраняется более или менее строгий порядок: «головные» гены впереди, «хвостовые» — сзади. У более примитивных представителей многоклеточных, таких как гребневики (Ctenophora) и кишечнополостные (Cnidaria), этих эмбриональных регуляторных генов только четыре, у млекопитающих их уже 48.
Семейство Hox-генов подразделяется на 14 классов. Считается, что эти 14 классов возникали путем дупликации (удвоения) одного или немногих исходных генов. «Дубликаты» затем мутировали и обретали новые функции. У примитивных кишечнополостных и гребневиков имеется всего 4 класса Hox-генов, у предполагаемого общего предка двусторонне-симметричных животных их должно было быть по крайней мере 8, у млекопитающих присутствуют все 14 классов.
Принцип работы всех Hox-генов одинаков. Все они являются транскрипционными факторами (регуляторами транскрипции — «прочтения» генов), то есть их функция состоит во «включении» или «выключении» других генов. В результате работы Hox-факторов запускается каскад реакций, приводящий к появлению в клетке нужных белков.

Обобщенная схема эволюции основных групп многоклеточных животных. Внизу показано предполагаемое строение Hox-кластера у общего предка билатеральных животных. У него предположительно имелось 8 Hox-генов в едином кластере. Гены нарисованы разными цветами и пронумерованы. Цифры дробью указывают, что данные гены совмещены, то есть еще не разошлись в ходе эволюции. Некоторые гены показаны одинаковым цветом, это означает большое сходство в их нуклеотидных последовательностях. Стрелочки указывают направление считывания генов на ДНК.
В последние годы представления о Hox-генах существенно изменились и расширились. Этому способствовала расшифровка ДНК-последовательностей Hox-генов у многих групп животных: кольчатых, плоских и круглых червей, иглокожих, членистоногих, оболочников, ланцетников, не говоря уже о млекопитающих. Под тяжестью новых данных обрушилось представление об упорядоченном расположении Hox-генов в хромосомах. Выяснилось, что, например, у иглокожих первые три Hox-гена располагаются прямо перед последним (14-м), а начинается кластер с пятого гена. У круглых червей и оболочников Hox-гены вообще не образуют кластеров и их порядок в хромосомах не соблюдается вовсе. Это говорит о том, что правильный порядок экспрессии («включения») Hox-генов в различных частях эмбриона не обязательно должен соответствовать порядку расположения этих генов в хромосоме. Последовательность включения Hox-генов зависит помимо «места под солнцем» от ряда дополнительных факторов, которые сейчас активно изучаются (но выяснить пока удалось немногое).
Число классов Hox-генов в разных типах животных оказалось разным. Завораживающее сходство Hox-кластеров у мухи и человека тускнеет, лишь только к ним прибавляется еще какая-нибудь группа двусторонне-симметричных животных. Так, у круглых червей (нематод) найдено всего 5 классов из 14, у оболочников — 9, зато у ланцетника — все 14. У паразитических трематод (класс плоских червей) обнаружено всего 4 Hox-гена на двух хромосомах, и между этими четырьмя разбросаны другие гены, не связанные с Hox-семейством.
И вот вопрос: почему у двусторонне-симметричных плоских червей Hox-генов столько же, сколько и у радиально-симметричных кишечнополостных и гребневиков? У кишечнополостных — вроде бы понятно: недостающих четырех классов генов еще не было, они образовались только после того, как появился гипотетический предок двусторонне-симметричных животных. Но у червей-то куда делась половина предковых регуляторных генов, зачем они избавились от них? Или, может быть, этой недостающей половины не было и у предка? Ответа пока нет. Ясно только, что даже при потере большого числа Hox-генов оставшиеся могут организовать эмбриональное развитие сложного билатерального существа.
Зато стали известны детали регуляции самих регуляторных Hox-генов. Между Hox-генами расположены участки ДНК, прежде считавшиеся бессмысленными. В действительности, как оказалось, с них считываются короткие молекулы регуляторных РНК. Некоторые из них усиливают или ослабляют экспрессию (работу) самих Hox-генов, некоторые косвенно влияют на работу других транскрипционных факторов. В экспериментах показано, что эти микроРНК могут регулировать как соседний, так и отдаленный Hox-ген.
Так что семейство Hox-генов, главное из главных среди генов-регуляторов, само не имеет полной власти в своем хозяйстве. За ним следят мелкие «выскочки» РНК, способные видоизменить экспрессию гена и тем самым замедлить или ускорить формирование органа.
(Источник: Derek Lemons, William McGinnis. Genomic Evolution of Hox Gene Clusters // Science. 2006. V. 313. P. 1912–1922.)
—————
Пожалуй, самый главный вывод, который можно сделать на основе анализа генома актинии, состоит в том, что уже самые первые представители животного царства обладали весьма сложным и совершенным «набором рабочих инструментов», то есть генов, который позволил создать огромное разнообразие сложных многоклеточных организмов, внося лишь небольшие изменения в базовую программу развития. Более того, эти изменения, судя по всему, вносились не в «архитектуру» программы, а только в ее «настройки».
Базовая генетическая программа поведения клетки, имевшаяся уже у первых животных, оказалась столь удачной и гибкой, что дальнейшая эволюция животного царства — в том числе и прогрессивная эволюция, связанная с усложнением организма, — уже почти не требовала радикальных изменений самой программы. Достаточно было лишь небольших изменений в «настройках». Менялись, в частности, регуляторные участки ДНК, от которых зависит тонкая настройка работы генов-регуляторов.
—————
«Главный секрет» животных. На интуитивном уровне мы привыкли относиться к царству животных как к чему-то огромному и чуть ли не бесконечно разнообразному. Но в последнее время все больше появляется фактов, которые показывают, что в действительности животные (Metazoa) представляют собой весьма специфическую, компактную и генетически однородную группу организмов. Прочтенный геном актинии — яркое тому свидетельство.
По-видимому, «самый большой секрет» животных, та генеральная идея, на которой основаны их строение и эволюция, заключается в особой технологии построения сложного организма из множества изначально одинаковых модулей — клеток. Суть технологии в том, что благодаря деятельности ряда ключевых генов — регуляторов развития (в том числе Hox-генов) между делящимися клетками складывается сложная система взаимоотношений, клетки обмениваются сигналами, градиенты концентраций регуляторных веществ задают симметрию и план строения развивающегося организма, и все эти факторы вместе направляют процесс самоорганизации, самосборки сложного многоклеточного существа из генетически идентичных (то есть изначально одинаково «запрограммированных») клеток.
Очень важно помнить, что геном работает не на уровне организма, а на уровне клетки. По сути дела он реально кодирует лишь биохимию и поведение одной клетки. Никакой «программы развития организма» в оплодотворенном яйце нет: там есть программа поведения клетки, не более. Что же касается «программы развития», то она самозарождается из взаимодействия делящихся клеток уже в ходе самого развития (примерно так же, как это происходит у Protozoon).
—————
Развитие червей начинается с хвоста. Нематода (круглый червь) Caenorhabditis elegans — замечательный модельный объект для изучения индивидуального развития животных, равно как и для многих других исследований. Достаточно сказать, что в 2002 году Сиднею Бреннеру, который первым начал использовать этого крошечного червя в качестве модельного объекта в генетике и эмбриологии, была присуждена Нобелевская премия «за открытия в области генетического регулирования развития человеческих органов». Работа, конечно, проводилась на черве, но выводы оказались справедливыми и для человека — вот вам и еще одно доказательство генетического единства животного царства.
Генетики и эмбриологи выявили целый ряд генов, влияющих на развитие червя, причем многие из этих генов имеются также и у других животных, включая человека. Как мы уже знаем, система генетической регуляции развития, включающая Hox-гены и ряд других генов-регуляторов, в общих чертах сходна у всех животных. Однако ученые пока еще очень далеки от полного понимания тех удивительных механизмов, которые заставляют дробящееся яйцо превращаться не в комок клеток, а в сложно и тонко организованное многоклеточное существо.
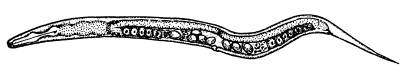
Червь Caenorhabditis elegans — любимец генетиков и эмбриологов. Одно из главных достоинств С. elegans — прозрачность. Под микроскопом хорошо видны на просвет все внутренние органы и даже отдельные клетки. Чтобы увидеть во всех подробностях, как из оплодотворенной яйцеклетки развивается червячок с пищеварительной и нервной системами, мускулатурой, органами размножения, достаточно просто положить яйцо С. elegans на предметный столик микроскопа и наблюдать за ним в течение 14 часов — именно столько продолжается эмбриональное развитие этого животного.
Генетический анализ позволяет лишь выявить гены, необходимые (но вовсе не обязательно достаточные) для тех или иных аспектов дифференцировки клеток. Например, мутации Hox-генов могут приводить к причудливому перемешиванию признаков, характерных для разных отделов тела (на голове дрозофилы могут вырасти ноги, на заднем сегменте груди — дополнительная пара крыльев). Известны гены, мутации в которых приводят к полному нарушению процессов клеточной дифференцировки, так что вместо нормального эмбриона действительно образуется бесструктурный комок клеток. Но чтобы понять, как все эти гены вместе руководят развитием, необходимы эмбриологические эксперименты.
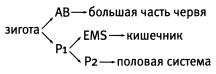
С. elegans — идеальный объект для таких исследований. Развитие этого червя прослежено во всех деталях — от оплодотворенного яйца до взрослого организма. В точности известна судьба каждой зародышевой клетки (бластомера); известно, из какого бластомера образуется каждая из клеток взрослого червя. Надо сказать, что развитие круглых червей, в отличие от многих других животных, очень строго определено с самого начала. Даже на стадии двухклеточного эмбриона его бластомеры отнюдь не одинаковы. Разделив их, мы не получим близнецов, как это бывает у позвоночных. Более крупный передний бластомер АВ дает большую часть тела, мускулатуру, нервную систему; задний бластомер P1— меньшую часть тела, включая половые органы и кишечник. Таким образом, судьба этих бластомеров известна уже на двухклеточной стадии. Она предопределена изначальной полярностью яйцеклетки: одному бластомеру достается больше каких-то регуляторных (сигнальных) веществ, другому меньше.
При этом, однако, бластомеры вовсе не являются замкнутыми системами, изначально запрограммированными на какой-то определенный путь развития. Они не могут правильно развиваться без контакта с другими бластомерами; они обмениваются между собой разнообразными химическими сигналами, корректируя свое поведение в соответствии с переменами в клеточном окружении.
Маркус Бишофф и Ральф Шнабель из Института генетики Технического университета в Брауншвейге задались целью выяснить, какой механизм отвечает за придание эмбриону передне-задней полярности. В экспериментах им чрезвычайно помогло то обстоятельство, что с ранними эмбрионами С. elegans можно обращаться как с миниатюрным живым конструктором. Бластомеры можно разделять и перекомбинировать произвольным образом и даже составлять химерные эмбрионы из клеток разных особей. До взрослого червя такие конструкции, как правило, дорасти не могут, но все-таки умирают не сразу и некоторое время развиваются.
Исследователи отделили у четырехклеточных эмбрионов две передние клетки («дочки» бластомера АВ), из которых в норме должен развиться почти весь червяк. Лишенные контакта с задними бластомерами («дочками» бластомера P1, которые называются EMS и P2), эти клетки делились хаотически и превращались в аморфный комок без всяких признаков передне-задней полярности.
Естественно было предположить, что задние бластомеры выделяют какое-то сигнальное вещество, упорядочивающее деление передних клеток. Так оно и оказалось. «Поляризующим центром» зародыша, как показали эксперименты, служит бластомер P2 (из него впоследствии развивается половая система). Достаточно приложить клетку P2 к потомкам АВ хотя бы на пять минут, чтобы клетки «поляризовались». После этого потомки АВ делятся не случайным образом, а преимущественно в определенной плоскости, так что в результате из них получается не комок, а удлиненный червеобразный зародыш с выраженным передним и задним концом. Задним концом всегда становится та точка, к которой прикоснулась клетка P2.
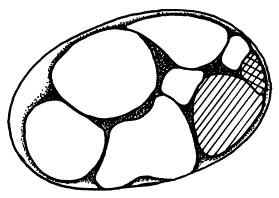
Зародыш С. elegans на стадии 8 клеток.
Очевидно, сигнальное вещество, выделяемое клеткой P2, влияет на ориентацию плоскости деления у дробящихся бластомеров. Сила этого вещества такова, что, прикладывая к клеткам два P2-бластомера в разных местах, ученые получали причудливых монстров с двумя хвостами или L-образно изогнутые зародыши с двумя взаимно перпендикулярными передне-задними осями. Каждая точка контакта с P2 превращалась в хвост!
Теперь надо было выяснить, что это за вещество. Ранее было установлено, что на ориентацию плоскости деления клеток оказывает влияние сигнальный белок Wnt, один из важнейших многофункциональных регуляторов развития у животных. Этот белок в иерархии регуляторов стоит в некотором смысле даже выше Hox-генов. Одни клетки выделяют белок Wnt, другие воспринимают его при помощи специальных белков-рецепторов, и это приводит к активизации других генов-регуляторов, в том числе Hox-генов.
Чтобы проверить, действительно ли бластомер P2 поляризует остальные бластомеры при помощи белка Wnt, исследователи использовали мутантные P2-бластомеры с выключенным геном Wnt. Оказалось, что такие P2-бластомеры поляризующим действием не обладают. Таким образом, природа сигнала была установлена.
Оставалось понять, каким образом сигнал передается от одних клеток к другим. Исследователи обнаружили, что клетки, «поляризованные» P2-бластомером, сами приобретают способность поляризовать другие клетки. Однако если у них выключен ген Wnt, этого не происходит. Из этого авторы сделали вывод, что передача сигнала происходит не путем диффузии сигнального вещества, производимого P2-бластомером, из клетки в клетку (как у модельного существа Protozoon из прошлой главы), а по принципу эстафеты. Поляризованные клетки сами начинают производить белок Wnt и таким образом поляризуют своих соседей.
Дополнительные эксперименты показали, что оставшаяся часть зародыша (та, что получается из бластомера EMS) тоже поляризуется благодаря контакту с P2. Таким образом, бластомер P2 оказался главным организующим центром развивающегося зародыша, определяющим его передне-заднюю полярность. Это важное открытие германских эмбриологов, а также разработанные ими методики сложных манипуляций с бластомерами открывают новые захватывающие перспективы перед учеными, стремящимися проникнуть в тайну развития животных.
Данная работа очень наглядно показывает, каким образом довольно простые системы химического «общения» между клетками зародыша позволяют им «самоорганизоваться» в сложный многоклеточный организм.
(Источник: Marcus Bischoff, Ralf Schnabel. A Posterior Centre Establishes and Maintains Polarity of the Caenorhabditis elegans Embryo by a Wnt-Dependent Relay Mechanism // PLoS Biology. 2006. 4(12): e395.)
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Сравнительная редкость цветов
Сравнительная редкость цветов Среди обитателей умеренного пояса сильно распространено мнение, что пышная растительность тропиков должна порождать также и великолепную флору цветов; такой взгляд подтверждается и многочисленностью крупных красивых цветов, разводимых
"Ранняя Соя" (Soja hispida praecok )
"Ранняя Соя" (Soja hispida praecok ) Ранняя соя, вывезенная нами в 1893г из Азии и акклиматизированная в Европе, была выпущена в продажу в 1899г Гетмановским Имением. Соя наша проникла во все хлеборобные губернии и области России, а также заграницу и в нынешнем году в Каталоге Вильморена
ОТ СТРУКТУРЫ К ФУНКЦИИ (функциональная геномика)
ОТ СТРУКТУРЫ К ФУНКЦИИ (функциональная геномика) Недостаточно знания, необходимо также применение; недостаточно хотеть, надо и делать. И. Гёте Хорошо известно выражение, что от слова «халва» во рту слаще не становится. Так же обстоит дело и с нашим геномом. В нем есть
ВСЕ ПОЗНАЕТСЯ В СРАВНЕНИИ (сравнительная геномика)
ВСЕ ПОЗНАЕТСЯ В СРАВНЕНИИ (сравнительная геномика) Между двумя представителями рода человеческого сходства меньше, чем между двумя различными животными. Мишель де Монтень То, что ново в себе, будет понято только по аналогии со старым. Ф. Бэкон Как уже говорилось,
ВСЕ ПОЗНАЕТСЯ В СРАВНЕНИИ (сравнительная геномика)
ВСЕ ПОЗНАЕТСЯ В СРАВНЕНИИ (сравнительная геномика) Между двумя представителями рода человеческого сходства меньше, чем между двумя различными животными. Мишель де Монтень То, что ново в себе, будет понято только по аналогии со старым. Ф. Бэкон Как уже говорилось,
Глава 11. Генетическая эволюция продуктов поведения животных
Глава 11. Генетическая эволюция продуктов поведения животных Что именно мы понимаем под фенотипическим эффектом гена? Поверхностное знание молекулярной биологии может предложить нам такой ответ: каждый ген кодирует синтез одной полипептидной цепи, и в простейшем смысле
Глава 3 Сравнительная геномика: эволюционирующие геномные ландшафты
Глава 3 Сравнительная геномика: эволюционирующие геномные ландшафты Пер. А. НеизвестногоВажность перехода к геномикеВ догеномную эру были установлены фундаментальные принципы молекулярной эволюции и было сделано немало конкретных наблюдений, имеющих большое значение
Глава 4 Геномика, системная биология и универсалии эволюции: эволюция генома как феномен статистической физики
Глава 4 Геномика, системная биология и универсалии эволюции: эволюция генома как феномен статистической физики Пер. К. МуратовойВ предыдущей главе была подчеркнута относительная стабильность отдельных генов, составляющая яркий контраст динамизму геномной эволюции.
Глава 19 СРАВНИТЕЛЬНАЯ ПСИХОФИЗИОЛОГИЯ
Глава 19 СРАВНИТЕЛЬНАЯ ПСИХОФИЗИОЛОГИЯ Сравнительная психофизиология – наука, нацеленная на установление закономерностей и выявление различий в структурной и функциональной организации мозга, поведении и психике у животных разных видов, в том числе и у человека.
СРАВНИТЕЛЬНАЯ АНАТОМИЯ
СРАВНИТЕЛЬНАЯ АНАТОМИЯ Биологам, занимающимся сравнительной анатомией, давно известно, что виды, образующие более крупные группы (тип, класс или отряд), имеют в своем строении много схожих черт. Развитие техники генетического анализа позволило установить более прочные
РАННЯЯ СЕНСОРНАЯ ДЕПРИВАЦИЯ
РАННЯЯ СЕНСОРНАЯ ДЕПРИВАЦИЯ Невозможность осуществления адекватного виду животного импринтинга, приводит к ранней сенсорной депривации (депривация — лишение, отсутствие чего-либо), вызывающей часто необратимые изменения структурной и функциональной организации
СРАВНИТЕЛЬНАЯ СИМВОЛИКА ГЕНОВ
СРАВНИТЕЛЬНАЯ СИМВОЛИКА ГЕНОВ Читатели, которые интересуются литературой по генетике, рано или поздно сталкиваются с проблемой путаницы в обозначениях генов. Дело в том, что различные авторы пользуются различными символами для обозначения одного и того же гена. Это
Глава II. Ранняя Земля
Глава II. Ранняя Земля 2.1. Образование Солнечной системы Около 4.6 млрд лет тому назад газопылевая туманность, из которой затем сформировалась наша Солнечная система, вступила в фазу сжатия. Участившиеся столкновения пылевых частиц вызывали разогрев материи особенно в
Ранняя диагностика и индивидуальное лечение
Ранняя диагностика и индивидуальное лечение Генетики уже давно заняты сравнением геномов здоровых и злокачественных клеток. Они обнаружили ряд генов, патологическое изменение которых резко повышает риск заболеть раком. Сегодня с помощью генетического анализа можно