Глава 16 Не связано ли это с размножением?
Глава 16
Не связано ли это с размножением?
Нельзя постепенно эволюционировать от четвероногого существа к двуногому. Какой должна быть промежуточная стадия — трехногой? Я такой никогда не видел.
Тимоти Уайт
Говоря обобщенно, сексуальность человека постоянна, а сексуальность животного периодична. Человек способен к размножению в любое время, животное — только в определенные периоды… Это отличие станет для нас особенно ясным, если мы попробуем представить себе, что в человеческом обществе лица противоположного пола будут проявлять интерес друг к другу только в летнее время, как, например, певчие птицы, или испытывать половое влечение один раз в несколько месяцев, как собаки, или вообще лишь однажды в течение всей жизни, как муравьи.
Джулиан Хаксли
Людям, возможно, не приходит в голову, что прямохождение связано с полом, но тем не менее это так.
Оуэн Лавджой
«Для любого четвероногого встать на задние лапы и побежать было бы сплошным безумием, — сказал Оуэн Лавджой, специалист по локомоции. — С точки зрения чистой эффективности бегать на двух конечностях абсурдно». Так рассуждал Лавджой, когда в начале 1980 года зашел в мою лабораторию, чтобы побеседовать со студентами о принципах локомоции. Я решил, что мне полезно будет послушать их беседу.
«Я полагаю, что это совершенно нелепо, — говорил Лавджой. — Даже попытки объяснить прямохождение кажутся нелепыми.
Утверждают: человек переселился в саванну и научился вставать на ноги, чтобы смотреть поверх высокой травы. Сущая чепуха. Это могло бы помочь ему, если бы он уже умел это делать. Но научиться этому после того, как он пришел в саванну, было невозможно. Он никогда не смог бы этого сделать.
Говорят: человек пользуется орудиями — он должен был встать на задние конечности, чтобы освободить руки для манипулирования предметами. В конечном итоге — да. Но не с самого начала. Эта идея всегда казалась мне сомнительной, а находки ископаемых существ из Летоли и Хадара полностью опровергли ее. Эти существа ходили на двух ногах, но еще не употребляли орудий. Они передвигались этим способом, может быть, уже миллион лет, прежде чем их потомки начали использовать орудия.
Говорят: человек в течение какого-то времени передвигался, опираясь на согнутые пальцы рук, подобно горилле и шимпанзе, и постепенно встал на ноги. Ерунда. Сама идея о том, что способ локомоции шимпанзе представляет собой переходную стадию на пути к прямохождению, — несусветная глупость. Передвижение с опорой на фаланги пальцев — это сугубо специализированная черта, которая связана с адаптацией к определенному образу жизни и не может привести к прямохождению».
«Чтобы разобраться в способах локомоции, — продолжал Лавджой, — нужно видеть в них не просто элементы причинно-следственных связей, а часть общей стратегии, направленной на выживание вида. Палеонтологи и анатомы склонны выделять в строении животного какую-то одну характерную черту, а затем провозглашать животное ее носителем: вот это — брахиатор, а то — двуногое существо. Это очень упрощенный подход. На самом деле локомоторная система животного, так же как и его репродуктивная система, составляет часть сложного адаптационного комплекса, связанного со специфической экологией. Должна существовать целая группа адаптации, функционирующих совместно. Короче говоря, чтобы разобраться в локомоторной системе шимпанзе, нужно понять, какова общая стратегия этого животного, направленная на выживание. Но еще раньше надо иметь хотя бы некоторое представление о том, что такое руки и ноги и как они работают». По словам Лавджоя, даже преподаватели биомеханики демонстрируют в этом вопросе потрясающее невежество. Поэтому он решил начать с элементарных принципов.
В самом общем виде тело — это кусок плоти, лежащий на земле или в воде. Чтобы добывать себе пропитание, тело должно двигаться. Для перемещения в воде ему нужны плавники и хвост, а на земле — ноги (исключение составляют змеи). Если единственная функция ног состоит в том, чтобы проталкивать тело вперед, тогда каждый раз при движении будет теряться много энергии в результате трения и других помех. Вот почему животному, чтобы ходить или бегать, нужно иметь такие конечности, которые могли бы не только продвигать тело вперед, но и поддерживать его над землей.
Ноги древнейших передвигавшихся по земле позвоночных — амфибий и рептилий, в том числе существующих и поныне тритонов и аллигаторов, — выполняли обе эти функции, но недостаточно хорошо. Они отходили от боков тела, что очень неудобно для бега (это видно, если посмотреть на аллигатора спереди). Чтобы просто поддерживать вес тела с помощью торчащих в стороны конечностей, уже необходимо достаточное усилие. Это все равно что лежащий на полу человек попытался бы отжаться, опираясь на руки, раскинутые в стороны. Вот почему аллигатор не может бегать быстро и на большие расстояния. Он движется, одновременно перенося переднюю левую ногу и заднюю правую. При ходьбе он делает это медленно, при беге — быстрее, но больше он ни на что не способен.
У млекопитающих ноги — гораздо лучшее приспособление для ходьбы и бега, чем у рептилий. Они направлены вниз, а не в стороны, поэтому лошадь может целый день оставаться на ногах, поддерживая свой вес, в то время как у аллигатора при такой попытке силы очень скоро иссякли бы. А лошадь не только стоит и ходит, она еще может бежать рысью или скакать галопом. Бегущий рысцой аллигатор — вещь невозможная.

— Почему же аллигаторы такие неприспособленные? — спросил Лавджоя кто-то из слушателей.
— Это не так.
— Но вы только что сказали…
— Я сказал, что они плохо бегают. А это разные вещи. Аллигатор великолепен, когда бесшумно соскальзывает с илистой отмели. Он прекрасный подводный охотник. Вспомните, что я говорил о группе совместно функционирующих адаптации. Вы должны рассмотреть весь комплекс используемых животным средств эволюционной стратегии, чтобы понять отдельные компоненты. Лошадь в чем-то добивается успеха, но уж, конечно, не в подводном плавании. И это отражено в строении ее ног.
Конечности млекопитающих формировались в процессе развития способности к бегу. Они не только переместились, расположившись прямо под телом, но и развернулись — задняя таким образом, что колено смотрит вперед, а передняя так, что «локоть» повернут назад. Эти перестройки делают более эффективным преобразование энергии при беге, а также ее поглощение.
Лавджой объяснил далее, что весь бег можно представить как процесс создания кинетической энергии и ее поглощения. Кинетическая энергия создается, когда задние ноги толкают животное вперед, и поглощается передними ногами, когда они опираются на землю. Если бы тело животного парило в пространстве и на него подействовал импульс какой-то силы, то оно приобрело бы некоторую скорость в соответствии с величиной этого импульса. В безвоздушном пространстве оно продолжало бы вечно скользить вперед. Однако в реальных условиях животные не парят в вакууме. При беге они должны постоянно вновь касаться земли, каждый раз теряя при этом часть энергии поступательного движения. Вот почему для дальнейшего движения необходим приток очередной порции энергии. Но животное упадет на землю, если его передние ноги не будут выброшены вперед и их мышцы не поддержат тело. И так без конца: энергия поступает, чтобы обеспечить продвижение тела, и поглощается в процессе его поддержания на достаточной высоте.
Чтобы понять, как происходит поглощение кинетической энергии тела передними ногами, нужно представить их в виде своеобразных амортизаторов, смягчающих удар при соприкосновении с землей. Суставы их сгибаются под давлением силы, направленной вниз, а мышцы противодействуют ей. Сопротивление мышц, которые стремятся выпрямить переднюю ногу, согнутую под действием веса тела, можно назвать «отрицательной силой». Она-то и гасит ту кинетическую энергию, которая высвобождается во время поступательного толчка задних ног.
Чтобы действовать как эффективный амортизатор, передние конечности не должны быть жестко соединены со скелетом тела. И их действительно удерживает на месте только мускулатура. Если бы эти конечности были закреплены более жестко, мышцы не могли бы мягко сопротивляться давлению тела, опускающегося вниз. Тогда поглощение кинетической энергии сопровождалось бы сокрушительным толчком. Благодаря тому что передняя конечность отделена от осевого скелета (а не фиксирована в костной впадине, как задняя), поглощение энергии при каждом шаге бегущего животного происходит плавно.
Задняя нога млекопитающего, напротив, не должна играть роль амортизатора. Ее функция состоит в том, чтобы добавлять кинетическую энергию. Поэтому верхний конец бедренной кости непосредственно входит в вертлужную впадину и удерживается там сухожилиями и связками. Прямое сочленение прочнее, и для передних конечностей оно тоже было бы более предпочтительным, если бы перед ними не стояла важная задача поглощения энергии. Спортсмены знают о недостатках непрямого прикрепления. У них часто случается вывих плечевого сустава вследствие разрыва мышц, удерживающих плечевую кость. Но прямое сочленение амортизирующей конечности принесло бы еще большие неудобства.
Чтобы понять это, вам достаточно спрыгнуть на всю ступню, не сгибая ног, с высоты около 30 сантиметров. Прямое причленение бедренной кости к тазу и выпрямленное колено полностью исключают возможность плавного поглощения энергии.
Рассмотрев эту модель типичного четвероногого млекопитающего — с торчащими вниз конечностями, из которых задние повернуты вперед, а передние назад, задние присоединяются прямо к скелету, а передние через посредство мышц, — легко будет понять, почему при переходе к двуногому хождению она становится менее эффективной. Лучше всего это можно продемонстрировать, рассмотрев разложение сил при беге. Если большая часть энергии расходуется на поступательное движение, а меньшая — на поддержание тела над землей, то тело будет перемещаться с большой скоростью. Если же энергия тратится в основном на поддержание тела, то высокая скорость невозможна. В этом и состоит недостаток двуногого хождения — слишком много энергии идет на то, чтобы тело не падало на землю.
В качестве примера рассмотрим идущего человека. В середине шага, когда одна нога опирается на землю, а другая выброшена вперед и висит в воздухе, тело продолжает двигаться вперед по инерции после предыдущего шага. К оставшейся кинетической энергии добавляется новая ее порция, когда человек выпрямляет ногу в колене и лодыжке, тем самым удлиняет ее и толкает тело вперед. В результате тело теряет равновесие, и другая нога, висящая в воздухе, должна быть опущена на землю для опоры. Это движение в значительной части гасит поступательную скорость. Действие сил, направленных вперед и вверх, можно представить в виде векторов, один из которых будет отображать истинное поступательное движение человека или его скорость (см. рис. на с. 232–233).
Чтобы увеличить скорость, чело век может наклониться вперед, усилив одну составляющую движения за счет другой. Правда, при этом возникнет опасность падения, так как сила, поддерживающая тело, уменьшится. Поэтому человек, чтобы не упасть, вынужден будет чаще прикладывать эту силу — быстрее работать ногами. Это и есть бег. В самом начале человек может сильно наклониться вперед (как спринтер во время старта), но когда ноги его начинают работать с предельной для него быстротой, он принимает почти вертикальное положение. Он может увеличить скорость, больше наклонившись вперед, но если его ноги не в состоянии двигаться быстрее, он просто упадет на землю. Этим нередко пользуются спринтеры в самом конце дистанции. Они с силой швыряют себя вперед и часто падают после пересечения финишной черты.
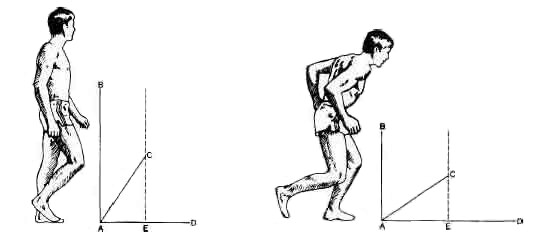
Во время ходьбы человек расходует больше усилий на поддержание тела (АВ), чем на его перемещение вперед (AD). Сумму этих двух сил можно изобразить вектором АС (косая линия). Истинная скорость человека пропорциональна горизонтальной проекции этого вектора (АЕ). Во время бега человек наклоняется вперед и начинает быстрее работать ногами. Теперь на поддержание вертикального положения тела расходуется меньшая доля усилий и соответственно большая доля тратится на поступательное движение. Результирующий вектор сильнее наклоняется в направлении движения, и расстояние АЕ, т. е. скорость, увеличивается.
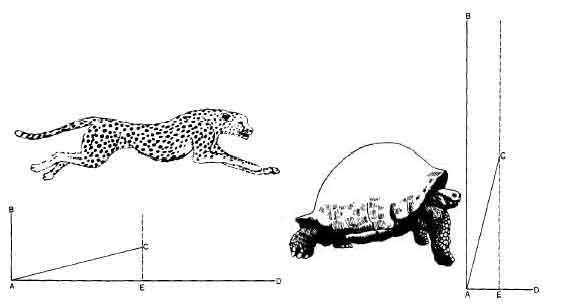
На примере гепарда можно показать, что при малых затратах энергии на движение, направленное вверх, и больших затратах на движение вперед скорость резко возрастает, в данном случае до 110 км/ч. Галапагосская черепаха, напротив, расходует почти всю энергию на поддержание огромного панциря. Ее максимальная скорость не превышает полутора километров в час.
Четвероногое животное не знает этой трудности. Его тело настолько наклонено вперед, что располагается параллельно земле. Большая часть энергии, доставляемой задними ногами, идет на поступательное движение, и лишь незначительную часть ее приходится гасить с помощью передних ног, чтобы поддерживать тело над землей. Передние ноги уже находятся впереди и готовы взять на себя функцию опоры во время кратковременного приземления животного, пока задние конечности поджимаются для очередного толчка. Среди четвероногих наиболее приспособленная «машина для бега» — это гепард. У него длинные ноги и длинный, очень гибкий позвоночник. Когда животное начинает свой прыжок, спина у него согнута. Одновременно с мощным толчком выпрямляющихся задних лап разгибается позвоночник, и гепард летит как стрела, выпущенная из лука, на шесть и более метров, теряя во время приземления на передние лапы лишь незначительную часть своей кинетической энергии.
— Но какое это имеет отношение к прямохождению? — спросили Лавджоя. — Вы объяснили нам, почему четвероногие бегают быстрее, чем двуногие. Но почему тогда некоторые из них перешли к «худшему» способу?
— Чтобы ответить на этот вопрос, — сказал Лавджой, — нужно вспомнить, что наши предки жили на деревьях, а потом спросить себя, чем они там занимались.
Самые древние прародители приматов, продолжал он, не были древесными жителями. Эти небольшие насекомоядные четвероногие, по размерам и форме тела напоминавшие белку, жили и охотились на земле. По мере развития мощного пояса тропических лиственных лесов, где нашли себе приют всевозможные древесные насекомые, не говоря уже о более крупных животных — небольших лягушках, ящерицах, змеях, — некоторые из мелких наземных хищников вслед за потенциальной добычей сами стали взбираться на деревья. С течением времени полог леса был заселен множеством хищных зверьков с когтистыми лапками. Они проводили наверху всю свою жизнь.
Одно дело ловить мелкую добычу на земле и совсем другое — на деревьях. Если некоторые жуки и все гусеницы передвигаются так медленно, что их можно собирать и поедать почти как ягоды, то ящерицы весьма проворны и легко ускользают от преследования. Не схватишь жертву при первом же прыжке — и она скроется на нижней стороне ветки и убежит прочь. Нападающий должен быть способен прыгнуть на добычу с некоторого расстояния, удержать ее и при этом (ведь все происходит в трехмерном пространстве!) не свалиться вниз. Короче говоря, мелкие хищники, чтобы успешно охотиться на деревьях, должны были уметь прыгать, хватать и цепляться. У самых ранних предков приматов, которые, как полагают, напоминали насекомоядных азиатских тупай, в связи с охотой на более крупную добычу вроде древесных лягушек или ящериц на передних конечностях начали, видимо, развиваться более длинные и более цепкие пальцы. Вместо когтей на верхней стороне пальцев появились плоские ногти, и сами пальцы стали походить на человеческие. Постепенно формировалась характерная для приматов «рука».
Ее владелец был уже не столько бегуном, как раньше, сколько прыгуном и гимнастом. В обмен на развившиеся хватательные способности он отчасти утратил ту ловкость, с которой его предки бегали по ветвям деревьев. Ни долгопят, ни галаго, если взять двух самых мелких приматов, и поныне лазающих по деревьям в тропических лесах, не сравнится в скорости передвижения с белкой. Животное, обладающее когтями, в этом отношении имеет преимущество перед тем, у которого развиты кисть и пальцы. Белка может бегать вверх и вниз по стволам любых деревьев, а приматы — нет. Маленькие цепкорукие создания живут наверху, а на землю спускаются лишь тогда, когда нужно перейти с одного дерева на другое. Крупные приматы, такие как шимпанзе, регулярно слезают вниз. Деревья средней величины они осваивают без труда, но могучие гиганты, стволы которых они не могут обхватить, для них недоступны. Чтобы карабкаться по большому дереву, нужно иметь когти.
Все это позволяет понять, почему у белок не развивались кисти и пальцы. Лазание и бег всегда были для них важнее, чем хватание. Эти животные питаются орехами, сосновыми шишками, семенами — неподвижными предметами, которые можно не спеша сорвать и на досуге грызть. Приматы же, напротив, сумели приспособиться к древесному образу жизни и одновременно сохранить свои хищнические наклонности.
Развитие руки приматов сопровождалось другими адаптациями. Для того чтобы успешно прыгать и хватать добычу, нужно хорошо оценивать расстояние. Без этого животное в лучшем случае останется голодным, а в худшем — не попадет на ветку и свалится на землю. Для точной оценки расстояния у приматов развивается бинокулярное зрение, при котором оба глаза центрируются на предмете, обеспечивая восприятие глубины. В этом случае глаза должны располагаться на передней стороне черепа, а не смотреть в стороны, как у белки. Аппарат бинокулярного зрения возникает у предков приматов. Их череп приобретает округлую форму, что соответствует новому положению глаз. В связи с этим емкость черепа возрастает и создается потенциальная возможность для развития более крупного мозга. В то же время челюсти становятся короче. С формированием кисти зубы перестают быть главным органом охоты и сбора пищи, и их становится меньше. Современные низшие и человекообразные обезьяны, так же как и люди, имеют по шестнадцать зубов в каждой челюсти. У их предков это число достигало двадцати двух.
Надежно защищенные от наземных хищников, ранние приматы чрезвычайно успешно освоили древесную среду обитания. Многие из них сделались более крупными и стали все больше специализироваться, приспосабливаясь к той или иной экологической нише. Одни начали питаться главным образом плодами и ягодами, другие — исключительно листьями. У них выработались разные способы передвижения по ветвям деревьев. Некоторые мелкие формы, такие как долгопяты и галаго, совсем отказавшись от бега, продолжали прыгать и цепляться за ветки; для них характерны короткие передние и длинные задние конечности. Одна обширная группа, уже обладавшая типичной для приматов кистью и даже стопой с уплощенной подошвой и цепкими пальцами, сохранила четвероногий способ локомоции. Это были низшие обезьяны. Они предпочитали ходить по ветвям, а не висеть на них или, раскачиваясь, перебрасываться с ветки на ветку. У них был довольно длинный позвоночник, и они сохранили способность к бегу, так как многие из них спускались на землю и должны были быстро передвигаться по ней, чтобы в случае опасности добежать до ближайшего дерева. Четвероногий способ локомоции отличает низших обезьян от человекообразных (антропоидов).
Антропоиды больше приспособлены к брахиации. Вместо того чтобы ходить по веткам, они висят на них, сидят или стоят, передвигаются вниз и вверх, перекидывая с ветки на ветку одну руку за другой. У них более короткий позвоночный столб — три или четыре поясничных позвонка, а не семь, как у низших обезьян. Поэтому они плохие бегуны. Некоторые из них почти не могут бегать.

У приматов встречаются самые разнообразные способы передвижения: Долгопят (на переднем плане) — примитивное ночное животное, ведущее исключительно древесный образ жизни, — прыгает и карабкается по веткам. Гиббон (вверху слева), тоже древесный житель, передвигается путем брахиации. «Четверорукий» орангутан (ниже и левее) с одинаковым успехом пользуется любой из своих конечностей Горилла (внизу слева) передвигается, опираясь на согнутые пальцы рук. У этой обезьяны полувыпрямленное положение тела, она может легко встать на две ноги и часто это делает. Низшая обезьяна (вверху справа) — настоящее четвероногое животное. У нее нет потенциальных возможностей поддерживать выпрямленную позу, которыми в той или иной степени обладают все человекообразные обезьяны. Зато она сохранила способность к быстрому бегу благодаря своим длинным конечностям и позвоночнику, а также кисти, которая при каждом шаге плоско опускается на землю. И только гоминиды осмелились стать полностью двуногими.
В наши дни на земном шаре существуют пять родов человекообразных обезьян. Представители двух родов — гиббоны и сиаманги — истинные брахиаторы. Они передвигаются с помощью брахиации, раскачиваясь подобно маятнику и перебрасываясь с ветки на ветку. У них необычайно длинные руки, удлиненные цепкие кисти и пальцы, небольшое легкое тело и укороченные ноги. Благодаря такой конструкции, сводящей к минимуму вес на конце маятника, эти приматы могут очень быстро передвигаться среди ветвей, уверенно и плавно перебрасывая свои длинные руки и часто покрывая расстояния до трех и более метров. Чтобы оценить красоту этих движений, нужно видеть их в натуре. Если гиббон спускается на землю, что бывает крайне редко, он выпрямляется и идет, переваливаясь на коротких, слабых ногах и раскинув по сторонам для равновесия непомерно длинные руки. Шагающий гиббон напоминает балансирующего канатоходца.
Орангутан, так же как и гиббон, ведет древесный образ жизни, но передвигается совершенно иным способом. Его тазобедренный сустав практически универсален. Обезьяна может вытягивать ногу вниз, назад, вперед, в сторону под прямым углом и почти вертикально вверх. Все ее конечности хорошо приспособлены к хватанию. По сути дела у орангутана четыре ноги и четыре руки. Подобно огромному рыжевато-коричневому пауку, он висит распростершись на дереве, держась за ветви теми руками или ногами, которыми ему удобнее, а свободными добывает себе пищу, главным образом плоды. В редких случаях, когда орангутан спускается на землю, он встает на четвереньки и передвигается медленно и осторожно, как старик, опирающийся одновременно на две трости (по выражению Сейрела Эймерла). Опорой орангутану служат подошвы ног и фаланги пальцев, но нередко он, сжав кисти в кулак, переносит тяжесть тела на них. Можно сказать, что орангутан лишь отчасти использует способ передвижения с опорой на суставы пальцев.
Зато горилла являет собой характернейший пример этого типа локомоции. Это антропоид, почти полностью перешедший от древесного образа жизни к наземному. Здесь горилла находит себе пропитание — грубые стебли растений, корни, побеги бамбука, ягоды. Горилла отказалась от возможности бегать и приобрела взамен крупные размеры и огромную силу (вес взрослого самца достигает 160 килограммов); вместе со свирепым видом это и обеспечивает ее выживание. Молодые гориллы висят и играют на ветках, но взрослые обезьяны слишком велики и малоподвижны для этого. Иногда они сидят, расположившись группой, на очень низких и толстых ветвях, однако обычно предпочитают отдыхать на земле. Эти животные не склонны к миграциям, они подолгу живут на одном и том же месте и почти никогда не бегают.
Шимпанзе тоже передвигаются, опираясь на фаланги согнутых пальцев. Это наименее специализированные из всех антропоидов. Они представляют собой как бы универсальную модель, пригодную для достижения разнообразных целей. Они поедают множество плодов, особенно фиг, и могут подниматься за ними в верхний ярус леса. У шимпанзе довольно длинные руки, которые позволяют им карабкаться вверх и вниз по стволам почти всех деревьев, кроме самых толстых. Ноги, хотя и не очень длинные, все-таки позволяют им на удивление быстро бегать. Человек, преследующий эту обезьяну, имел бы мало шансов догнать ее. Наделенные всеми этими свойствами, шимпанзе проводят большую часть времени на земле, питаясь термитами и другими насекомыми, их личинками, ягодами, почками и корнями, если фиги еще не созрели. Шимпанзе в какой-то небольшой мере способны к согласованным действиям во время охоты. Иногда группе самцов удается загнать молодого павиана или другую небольшую обезьянку на отдельно растущее дерево, поймать добычу и съесть ее.
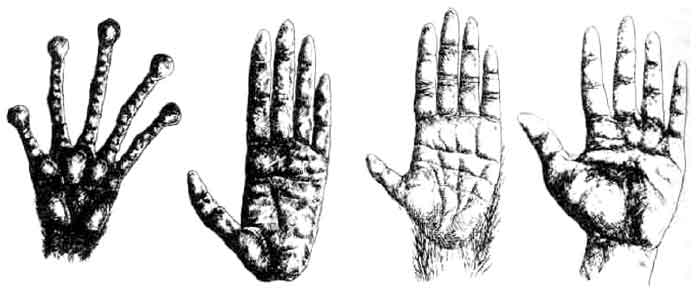
Долгопят, гиббон, шимпанзе, гоминид.
У всех приматов проворные, ловкие руки с гибкими пальцами, на концах которых имеются плоские ногти. Различия в строении кисти связаны со способами локомоции и образом жизни. У прыгающего и цепляющегося за ветки долгопята пальцевые подушечки сильно расширены, что помогает ему удерживаться на деревьях. У гиббона длинные сильные пальцы, которые во время брахиации играют роль крючьев. Шимпанзе — частично древесное, частично наземное животное. У него исключительно ловкие руки, способные даже манипулировать с грубыми орудиями; большой палец руки хорошо развит и может противопоставляться остальным, но при этом из-за своей малой длины доходит лишь до середины указательного пальца, а не до его кончика. У гоминид большой палец гораздо длиннее и развернут в сторону указательного. Этот признак сопутствует двуногому хождению, так как позволяет значительно увеличить манипуляторные способности. Вероятно, все гоминиды обладают кистью такого типа, даже A. afarensis, древнейший из известных представителей этой группы. Его кисть мало отличается от кисти современного человека.
Несмотря на столь различный образ жизни и связанные с ним особенности строения тела, все антропоморфные обезьяны — потенциально прямоходящие животные. Все они обитают в лесах, и их образ жизни не требует способности к бегу. Около 20 миллионов лет назад, в начале миоцена, землю опоясывала широкая полоса тропических и субтропических лесов. Это был период процветания антропоидов, которые по численности намного превосходили низших обезьян. В наше время наблюдается обратное соотношение между этими группами. Конечно, сокращение зоны тропических лесов, начавшееся в конце миоцена и связанное с изменениями климата, отразилось на состоянии человекообразных обезьян; но были и другие, менее очевидные факторы. Лавджой попытался проанализировать их и, как он считает, нашел причину перехода к прямо-хождению, связанную с взаимодействием этих факторов.
— Наконец-то мы вернулись к двуногому хождению, — пробурчал кто-то из слушателей. — Признаться, мне порядком надоели эти обезьяны.
— Надоели они вам или нет, это уже другой вопрос. Вы должны понять их происхождение: хищники, которые поднялись на деревья и сумели стать древесными жителями, сохранив в то же время свои хищнические наклонности, добились этого благодаря полувыпрямленному положению тела, способности к брахиации, развитию кистей рук, короткому позвоночному столбу, бинокулярному зрению и большим размерам мозга. Ничего подобного приматам природа раньше не знала. Когда некоторые из крупных обезьян, такие как горилла и шимпанзе, вторично спустились на землю, они с помощью вновь обретенных способностей смогли создать для себя новые экологические ниши.
— Но они не перешли к двуногому хождению.
— Некоторые сделали это: гоминиды, наши предки.
— Да, но почему?
— Не поговорить ли нам о размножении?
— Я бы предпочел беседовать о бипедии.
— Но поговорим все-таки о размножении.
Разговор о размножении — это в полном смысле слова возвращение к основам бытия. Здесь можно вспомнить остроумное замечание, сделанное почти сто лет назад и приписываемое Герберту Спенсеру: «курица — это средство, с помощью которого яйцо производит другое яйцо». Иными словами, взгляните на выживание вида не с точки зрения индивидуума, а с точки зрения незаметных, но все определяющих элементов — генов. Учитывайте, что в конечном счете самое важное — это сохранить набор генов, а тем самым и получить новую подобную себе особь.
Лавджой согласен в этом со Спенсером. Каждое живое существо, относящееся к любому виду, — не что иное, как защитная оболочка, в которой содержится семя для размножения, тщательно упакованное, чтобы обеспечить максимальную вероятность его выживания и воспроизводства потомков. Для достижения этой цели существо должно есть. А чтобы есть, если это млекопитающее, оно должно передвигаться. Поэтому для понимания локомоторной адаптации животного необходимо рассмотреть его стратегию в области размножения.
Существуют два фундаментально различных подхода к воспроизводству вида. Организм может продуцировать огромное количество яиц, затрачивая на каждое из них очень мало энергии; или же он может производить совсем мало яиц, но вкладывать в каждое много энергии. Эти два подхода известны в науке как «R-стратегия» и «К-стратегия» соответственно.
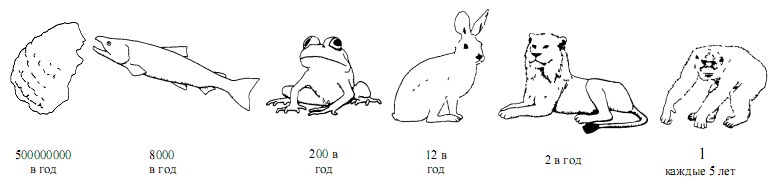
Репродуктивные стратегии в мире животных охватывают весь диапозон от крайних вариантов «R», когда образуется максимальное количество яиц, но нет никакой заботы о потомстве, до крайних вариантов «К», когда главную роль играет забота о потомстве, а частота рождений сведена до минимума.
Крайний пример R-стратегии — устрица, производящая до 500 миллионов яиц в год. Крайний пример К-стратегии — крупные антропоиды (горилла, шимпанзе и орангутан), рождающие детеныша один раз в пять или шесть лет. Между ними располагаются все остальные живые существа, у которых можно найти все переходные градации между R и К. Оба способа эффективны, пока не достигают предела своих возможностей. Предположим, устрица смогла бы удвоить число яиц — довести его до миллиарда. Но если общее количество расходуемой энергии должно быть постоянным, то на одно яйцо ее будет приходиться вдвое меньше. Это может настолько снизить жизнеспособность яиц, что число особей, достигающих зрелости, еще более сократится, хотя оно и так было очень невелико. Ясно, что R-стратегия в чистом виде невыгодна и что возможности этого способа уже используются до предела у таких низкоорганизованных животных, как устрицы. В самом деле, R-стратегию, доведенную до крайности, можно считать неэффективным способом размножения. Какой смысл производить полмиллиарда зародышей, если из них доживут до зрелости меньше десятка?
Однако у устриц нет другого выбора. У них отсутствует мозг, органы чувств находятся в зачаточном состоянии. Устрицы не способны передвигаться, не могут заботиться об отложенных яйцах или хотя бы поместить их в безопасное место. Все, на что они способны, — это выбрасывать зародышей в окружающую среду, увеличивая до разумного предела их число и повышая таким образом их суммарные шансы на выживание.
Хотя R-стратегия и приемлема для устриц (ибо они до сих пор существуют), она явно чересчур расточительна. По мере формирования более высокоорганизованных животных начинают использоваться другие, более экономные способы. Когда появились существа с позвоночником и головным мозгом, стала возможной примитивная забота о потомстве. Некоторые рыбы сооружают гнезда для своей икры; другие держат ее во рту до тех пор, пока из нее не разовьются мальки, способные плавать. У некоторых рептилий дела обстоят еще лучше. Аллигатор насыпает над отложенными яйцами большой холм и стережет их на протяжении всего периода инкубации. Когда детеныши начинают вылупляться и пищать, родитель, услышав их возню, разрывает землю и помогает им выбраться наружу. Однако лишь немногие из рептилий так заботливы, и нет ни одной, которая охраняла бы и воспитывала своих отпрысков в период их роста. Это прерогатива млекопитающих — интеллектуально более развитых и еще более «К-ориентированных» преемников рептилий.
Никто не может сказать, насколько заботливыми родителями были динозавры. Судя по группам окаменевших яиц, найденным в пустыне Гоби, по крайней мере некоторые из них откладывали сравнительно немного яиц: в отдельных гнездах их было всего лишь одна или две дюжины. Таким образом, если динозавры были такими же плохими родителями, как и большинство других рептилий, то у них сочетались минусы обеих стратегий — они не баловали своих отпрысков родительской заботой (R-стратегия) и в то же время откладывали мало яиц (К-стратегия). Это позволяет объяснить их вымирание в эпоху, когда появились К-ориентированные млекопитающие с более развитой заботой о потомстве.
— Чтобы понять, почему вымерли динозавры, нет надобности искать такие необычные причины, как пятна на солнце и климатические катастрофы, — заявил Лавджой. — Они процветали до той поры, пока мир принадлежал им одним и вокруг не было конкурентов с более эффективной стратегией размножения. Они просуществовали сто с лишним миллионов лет — было бы неплохо, если бы люди смогли продержаться столько же. Но как только был достигнут новый уровень адаптации и динозавры столкнулись с существами, способными размножаться в три-четыре раза быстрее, их час пробил.
К-стратегия, несомненно, гораздо эффективнее, чем R-стратегия. Однако и она имеет свои границы. Тяжелый урон наносят несчастные случаи, нападение хищников, сезонная нехватка пищи, болезни. Потерять детеныша вследствие одной из этих причин, после того как пять или шесть лет было затрачено на его выращивание, — очень дорогая цена по сравнению с гибелью зародыша устрицы. Два или три несчастных случая подряд могут привести к исчезновению определенной комбинации генов, носителем которой была конкретная самка, поскольку не исключено, что период ее плодовитости к тому времени уже закончится. Если взять целую популяцию антропоморфных обезьян со всем ее генным фондом, то для нее низкая скорость воспроизводства не менее опасна, чем для одной особи. Серия несчастий — и популяция навеки исчезнет.
В самом деле, уровень К-стратегии, которого достигли современные антропоиды, угрожающе высок. Все они сохранились ныне только в самых благоприятных условиях, и численность их уменьшилась до опасного предела. Эта чрезвычайная уязвимость человекообразных обезьян не всегда заметна на фоне того вклада, который вносит в их исчезновение современный человек. Мы привыкли слышать, что мы уничтожаем человекообразных обезьян. Несомненно, это так, и все же деятельность человека — это лишь капля, переполнившая чашу. Уже несколько миллионов лет дела антропоидов идут под уклон. Даже без вмешательства человека эти животные, вероятно, все равно исчезли бы. В отличие от этого низшие обезьяны все время процветали и размножались. Разница в конечных результатах между двумя группами настолько поразительна, что имеет смысл сравнить их и посмотреть, не объясняется ли она каким-то фундаментальным биологическим различием.
Оказывается, есть даже два таких различия. Низшие обезьяны — это, как уже говорилось, четвероногие существа. Антропоиды — потенциально двуногие. Низшие обезьяны по сравнению с антропоидами менее ориентированы в направлении К-стратегии. Детеныши рождаются у них раз в два года, а не в пять-шесть лет, как у антропоидов. Низшие обезьяны уступают человекообразным в отношении умственных способностей и несколько хуже заботятся о потомстве. Однако эти недостатки с лихвой восполняет более быстрое размножение. Современное благополучие низших обезьян, в большей степени ориентированных в сторону R-стратегии, позволяет думать, что К-ориентированные антропоиды зашли в этой стратегии слишком далеко.
Почему же предпочтение было отдано худшему варианту? Этот вопрос звучит уже во второй раз. Мы уже спрашивали: для чего было переходить к двуногому хождению, если это менее эффективный способ? А теперь спрашиваем: зачем понадобилось доводить К-стратегию до опасной крайности? Обе эти проблемы взаимосвязаны, и их нужно рассматривать вместе.
Прежде всего из нашего обсуждения эволюции приматов должно быть ясно, что локомоцию человекообразных обезьян нельзя считать неэффективной: они живут в тропических лесах и проводят время на деревьях, лазая по ветвям и перебрасываясь с ветки на ветку, или же под деревьями, где могут ходить и сидеть, им нет необходимости быть хорошими бегунами. Такие признаки, как короткий позвоночник и полувыпрямленное положение тела, приобретенные ранее во время жизни на деревьях, не создавали для некоторых из них особого неудобства и на земле, пока леса были обширными и снабжали их необходимой пищей.
Таким образом, четвероногий способ передвижения, несмотря на свои очевидные преимущества, может уступить место — и действительно уступает место — другому способу, если это связано с новыми, более важными адаптациями. Повторим то, что было сказано в начале этой главы: бег — настолько эффективный способ передвижения, что отказ от него выглядит нелепой причудой. Но при рассмотрении общей стратегии выживания преимущества этого типа локомоции могут исчезнуть. Вот почему в миоценовых лесах стало возможным появление потенциально прямоходящих обезьян, ведущих наземный образ жизни. Чтобы понять, почему некоторые прошли этот путь до конца, нужно рассмотреть, каковы были у них другие жизненные потребности и способы их реализации.
Необходимо также учесть, что с самого начала кайнозойской эры (около 70 млн. лет назад) до середины миоцена в ряду приматов все усиливалась тенденция к К-стратегии. Это происходило потому, что такая стратегия оправдывала себя. Самки человекообразных обезьян в результате прогрессивной эволюции могли проявлять все большую заботу о потомстве, и их детеныши выживали лучше, чем у менее заботливых матерей.
На этом уровне вступает в действие принцип положительной обратной связи, когда несколько элементов, взаимодействуя, усиливают друг друга. Если забота о потомстве — полезный признак, то отбор будет благоприятствовать его сохранению, так как самки с более развитыми материнскими качествами будут иметь больше шансов на успех в выращивании детенышей. Таким образом, существующая в популяции генетическая тенденция к развитию материнских инстинктов будет возрастать. Однако усиленная забота о потомстве требует также ряда других качеств. Матери нужен более высокий уровень умственных способностей, так как иначе она едва ли сможет обеспечить лучший уход за детенышами. А это означает развитие мозга, причем мозга не только самой матери, но и ее дочери, которая когда-то ведь тоже станет матерью. Вынашивание детенышей с крупным мозгом требует интенсивного потребления кислорода и переноса через плаценту значительного количества энергии, т. е. больших энергозатрат со стороны матери. Это связано с тем, что к моменту рождения детеныш с крупным мозгом должен быть достаточно развит, нужно, в частности, чтобы в основном закончилось созревание нервных механизмов. Детеныш ничему не сможет научиться, если с самого начала не будет обладать развитым мозгом. При врожденном недоразвитии мозга компенсации в дальнейшем уже не происходит. Поэтому за полноценное формирование детеныша ответствен организм матери. А поскольку ее возможные энергетические затраты всегда ограниченны (так же как и затраты на производство яиц у устриц), при более крупном мозге приходится иметь меньше детенышей.
И наоборот: при меньшем числе детенышей приходится иметь более крупный мозг, чтобы лучше заботиться о них. Таков принцип обратной связи. Все тенденции взаимосвязаны, зависят друг от друга и усиливают друг друга.
В случае эволюции приматов мы имеем дело не с простой обратной связью, когда два фактора воздействуют друг на друга, а с многофакторной круговой системой, которая включает несколько взаимоусиливающих компонентов. Например, если у детеныша большой мозг, то ему необходимо время, чтобы научиться использовать этот мозг, прежде чем он лицом к лицу столкнется с окружающим миром. Значит, у него должно быть продолжительное детство. А в детстве лучший способ обучения — игра, предполагающая наличие партнеров и, следовательно, групповую социальную организацию. Но, чтобы стать членом группы, нужно усвоить основы социального поведения. А достичь этого можно лишь при определенном уровне умственных способностей. Таким образом, социальное поведение в конечном итоге связано с развитием умственных способностей (петля обратной связи), с продолжительным детством (еще одна петля) и, наконец, с энергозатратами матери и системой заботы о потомстве, обеспечивающей определенный уровень развития мозга.
Так выглядит замкнутая система обратных связей, все части которой перекрестно соединены. Например, если какое-то животное входит в состав группы, то время, необходимое ему для того, чтобы найти пищу, обнаружить близость хищника или подыскать полового партнера, может уменьшиться именно благодаря его принадлежности к группе. Соответственно больше времени останется для заботы о потомстве (одна петля), игры (другая) и социальной активности (еще одна), что в свою очередь стимулирует развитие умственных способностей (еще одна петля) и в конечном счете ведет к уменьшению числа детенышей (еще одна). Это можно представить в виде замкнутого контура, все полюса которого соединены друг с другом (см. схему на с. 242).
— Подойдем теперь с другой стороны, — сказал Лавджой, — представим себе систему обратных связей без одного из ее существенных компонентов и посмотрим, что получится. Вообразите, например, детеныша шимпанзе, который вырос в лаборатории, лишенный возможности общения со своими сверстниками. Представьте себе, что он провел там детские годы, а затем, будучи подростком, попал в естественное сообщество шимпанзе. Он не сможет стать его членом, так как не будет знать, что он должен делать и как себя вести. Он не будет уметь прыгать и карабкаться на деревья, не будет знать, как нужно драться и как избегать драк, как следует вести себя по отношению к старшим. Молодой самец гориллы, выросший в изоляции, даже не умеет спариваться. Чтобы научить такую обезьяну нормальному поведению, приходится показывать ей соответствующие фильмы.
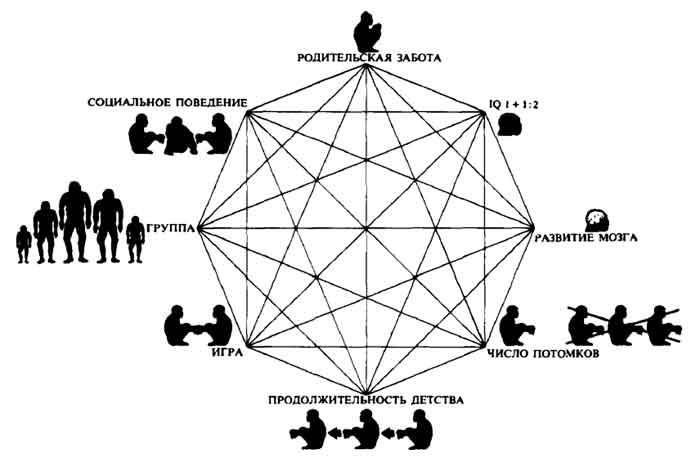
— Итак, — продолжал Лавджой, — существует целый ряд причин, объясняющих усиление К-стратегии. У позвоночных эта тенденция сопровождалась прогрессирующей специализацией. Лягушки более развиты по сравнению с устрицами, аллигаторы — по сравнению с лягушками, кролики — по сравнению с аллигаторами, мартышки — по сравнению с кроликами, антропоиды — по сравнению с мартышками. Более развитый мозг — меньшее число яиц — большая ориентация на К-стратегию.
— Разумеется, я не хочу сказать, что К-стратегия повышает умственные способности, но эти факторы, несомненно, связаны между собой. Поскольку умственные способности — явно адаптивная черта, понятно, что при подходящих условиях она может усиливаться. А эти условия для некоторых приматов, судя по всему, были вполне подходящими. Хотя мы располагаем только косвенными данными в виде ископаемых остатков, можно предполагать, что в миоцене человекообразные обезьяны достигли уже достаточно высокого умственного уровня. По-видимому, эти животные напоминали современных шимпанзе. Они ходили в полувыпрямленном положении, ловко манипулировали руками, имели более развитый по сравнению с низшими обезьянами мозг. Можно думать также, что у антропоидов миоцена была сильно выражена К-стратегия. Наконец-то мы достигли той точки в наших рассуждениях, когда можно вплотную заняться вопросом: почему в одной из линий человекообразных обезьян выработалось истинно двуногое передвижение? Или, другими словами, какая связь между К-стратегией и прямохождением?
Вопрос о прямохождении мучит антропологов уже целое столетие. Вскоре после первой находки яванского обезьяночеловека, сделанной в 1891 году, Дюбуа обнаружил бедренную кость, строение которой показывало, что ее владелец был прямоходящим существом. Однако ученые не могли поверить в сочетание столь примитивного черепа с прямохождением. Все думали, что эти две части скелета не связаны между собой: бедренная кость принадлежала сравнительно позднему человекоподобному существу и случайно оказалась рядом с черепом, ибо такое примитивное создание, как яванский обезьяночеловек, не могло передвигаться на двух ногах.
Это ошибочное представление пустило глубокие корни. Ему вновь отдали дань, когда Реймонд Дарт заявил, что детеныш из Таунга был прямоходящим существом. Даже в 60-х годах, много лет спустя после того, как мир был вынужден признать в австралопитеках прямоходящих приматов, антропологи все еще утверждали, что те передвигались шаркающей, неумелой походкой.
Большинство ученых пришли к согласию в том, что двуногое хождение, крупный мозг и использование орудий развивались параллельно. Для объяснения этого процесса был привлечен принцип обратной связи: каждый из признаков воздействовал на другие и усиливал их. Так, животное, начавшее использовать орудия, а с увеличением размеров мозга и создавать их, должно было часто переносить их с места на место, и это могло послужить стимулом для развития прямой походки. Освобождение верхних конечностей от локомоторной функции в свою очередь способствовало применению орудий и развитию мозга. Около двух миллионов лет назад, как утверждали сторонники этой идеи, была достигнута некая промежуточная стадия: мозг увеличился, но был все еще мал, орудия (судя по находкам из Олдувая) уже существовали, но были крайне примитивны — на нижней границе того, что вообще могло называться «орудиями», прямохождение (если считать, что этот признак формировался параллельно с двумя другими) тоже, — вероятно, находилось в процессе становления и не достигло полного развития.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ГЛАВА IX.
ГЛАВА IX. Углекислота в почве.Многие ученые объясняют богатую растительность древних времен тем, что тогда атмосфера содержала больше кислоты, чем теперь. Поэтому-то Либих был того мнения, что и в настоящее время, если желаем получить самый обильный урожай наших
ГЛАВА XI.
ГЛАВА XI. Атмосферное орошение (ирригация).В 1876 г. в нашей литературе появилась книга, заслуживающая самого серьезного внимания. Но так как книга осмелилась быть оригинальной, а мы привыкли верить, что только книги, написанные на основании трудов заграничных "авторитетов",
ГЛАВА XV.
ГЛАВА XV. Обработка земли под яровые хлеба.Обработку земли под яровые хлеба я начинаю тотчас после уборки озими . Только при соблюдении этого условия можно рассчитывать на самый обильный урожай.Поля, поросшие сорными травами и покрытые густым жнивьем, я вспахиваю
ГЛАВА XVI.
ГЛАВА XVI. Посев.Как нужно сеять, чтобы получить самый обильный урожай? Чтобы ответить на этот вопрос, нам нужно будет припомнить те условия, при которых растения развиваются в желательном для земледельца направлении. Эти условия, о которых мы говорили во второй главе,
Глава I
Глава I Восемь лет назад я написал небольшую книгу «Аксиомы биологии»[1], в конце которой высказал предположение, что возможно создание общей теории эволюции последовательно реплицирующихся систем. Завершил я книгу словами: «Под эту категорию попадают не только объекты
Глава 3
Глава 3 Описание метода лечебной кинологии и результатов исследования В предыдущих главах описывались теоретические аспекты метода лечебной кинологии. Постараемся схематически описать практический аспект методологии. Как уже отмечалось, данный метод находится на
Глава 3
Глава 3 Описание метода лечебной кинологии и результатов исследования В предыдущих главах описывались теоретические аспекты метода лечебной кинологии. Постараемся схематически описать практический аспект методологии. Как уже отмечалось, данный метод находится на
ГЛАВА 7 ПОВЕДЕНИЕ, СВЯЗАННОЕ С РАЗМНОЖЕНИЕМ[5]
ГЛАВА 7 ПОВЕДЕНИЕ, СВЯЗАННОЕ С РАЗМНОЖЕНИЕМ[5] Ритуализация полового поведения Поведение играет колоссальную, если не сказать ведущую, роль, как для самого процесса размножения, так и для полового отбора.Отбор идет по пути жесткой выбраковки партнеров, которые ведут себя
Глава 2. ДНК
Глава 2. ДНК На стене паба “Орел” в Кембридже висит синяя мемориальная доска, установленная в 2003 году в честь пятидесятилетия одного случая, когда разговоры в этом пабе приняли не совсем обычный оборот. Во время обеда 28 февраля 1953 года два завсегдатая “Орла”, Джеймс
ГЛАВА 5
ГЛАВА 5 С помощью Пам Смарт мне удалось разработать более простую и эффективную методику работы с людьми, испытывающими фантомные ощущения в отсутствующих конечностях, чем та, которая была описана в пятой главе.Мы провели серию опытов с людьми, у которых были
Глава III. Мир РНК-ДНК
Глава III. Мир РНК-ДНК 3.1. Миры до РНК и мир РНК Многие исследователи полагают, что первым клеточным миром был мир РНК (Ferris, 1999; Hoenigsberg, 2003). Однако по причинам, рассмотренным выше, более правдоподобна версия, согласно которой в ранних клетках функционировали информационные
Глава 10. Уши{10}
Глава 10. Уши{10} Того, кто заглянет поглубже в ухо, чтобы увидеть, как устроен наш орган слуха, ждет разочарование. Самые интересные структуры этого аппарата скрыты глубоко внутри черепа, за костяной стенкой. Добраться до этих структур можно только вскрыв череп, удалив мозг,