ГЛАВА 6 Зрение в ином свете
ГЛАВА 6
Зрение в ином свете
Глаза насекомых и других членистоногих представляют собой сложные органы, состоящие из множества одинаковых элементов. В каждом таком элементе, называемом омматидием (фиг. 20), представлены все составные части глаза; соседние омматидии отделены друг от друга слоем светонепроницаемого пигмента. Роговица представляет собой прозрачный участок твердой кутикулы, покрывающей все тело насекомого. Под роговицей расположен хрусталик, который фокусирует свет на ретинальных клетках. Эти фоторецепторные клетки (обычно их бывает 7 или 8) связаны с мозгом при помощи нервных волокон. В состав каждой такой клетки входит рабдомер — палочкообразная структура, ориентированная параллельно длинной оси клетки и расположенная на той ее стороне, которая обращена внутрь омматидия. Все вместе рабдомеры образуют единую структуру — рабдом. Рабдомеры служат как бы «световодами», по которым свет от хрусталика распространяется по краям клеток ретинулы и вызывает в этих клетках фотохимическую реакцию того же типа, что и в глазе человека.
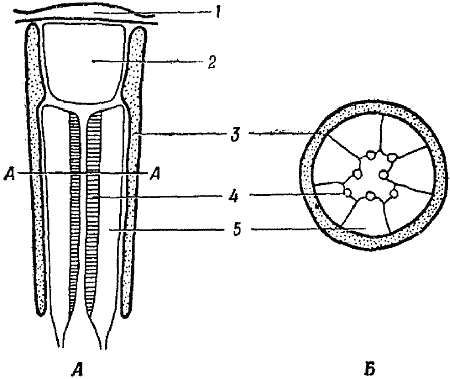
Фиг. 20. Продольный и поперечный разрезы омматидия — элемента сложного глаза насекомых
А. Продольный разрез омматидия. Каждый омматидий отделен от соседних омматидиев слоем пигмента 3. Роговица и хрусталик 2 фокусируют свет на рабдомерах 4, по которым он распространяется к светочувствительным пигментам в клетках ретинулы 5. Б. Поперечное сечение омматидия на уровне А — А, иллюстрирующее расположение клеток ретинулы и рабдомеров.
Всего несколько лет назад принципы функционирования сложного глаза казались совсем простыми, но впоследствии, когда были разработаны более тонкие методы исследования (например, микроэлектродная техника), прежняя теория была опровергнута. Хотя в настоящее время мы располагаем достаточно обширными сведениями о работе отдельных частей глаза насекомого, законченное представление о его функционировании у нас еще не сложилось. По классической теории, сложный глаз насекомого рассматривался как пучок трубочек (омматидиев), каждая из которых заканчивается светочувствительным элементом — ретинулой. Считалось, что омматидий пропускает к ретинуле только те световые лучи, которые более или менее параллельны его оси (фиг. 21, А), и, следовательно, воспринимает лишь небольшую часть окружающего мира — ту часть, которая находится прямо перед ним. Полная картина этого мира, как полагали ученые, создается всем комплексом омматидиев и представляет собой мозаику, состоящую из множества точек различной яркости, которая зависит от количества света, падающего на каждую ретинулу. Такая теория четко объясняла, каким образом насекомые обнаруживают движущиеся предметы. Если какой-то объект перемещается в поле зрения насекомого, он последовательно стимулирует все новые и новые омматидии. Однако мозаичное изображение должно быть очень грубым по сравнению с изображением, которое возникает в глазу человека. Другими словами, глаз насекомого, по-видимому, характеризуется очень слабой остротой зрения, или разрешающей способностью, которая в свою очередь зависит от числа омматидиев в глазу. Число омматидиев в каждом глазу стрекозы, которая летает очень быстро и обычно охотится, руководствуясь зрением, достигает 28 000. Глаз стрекозы может улавливать малейшие движения предметов на расстоянии 12 см, тогда как рабочий муравей с ничтожным числом омматидиев вряд ли способен на большее, чем отличать свет от тьмы.
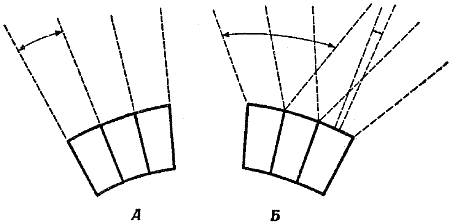
Фиг. 21. Схема, показывающая восприятие омматидиями световых лучей в соответствии с классической теорией зрения насекомых (А) и с результатами электрофизиологичёских экспериментов (Б)
Согласно классической теории, каждый омматидий может улавливать только лучи, параллельные его оси; острота зрения насекомого, по этой теории, определяется величиной угла между осями соседних омматидиев. Результаты современных исследований показывают, что отдельный омматидий способен различать световые лучи, падающие на него под разными углами; следовательно, глаз насекомого обладает большей остротой зрения, чем полагали раньше.
Существование мозаичного изображения впервые убедительно доказал немецкий ученый Экснер, который сфотографировал окно сквозь фасеточный глаз светляка, помещенный на предметное стекло микроскопа. На фотографии было видно расплывчатое изображение оконного переплета, а за ним — неясные очертания собора. Позднее определили остроту зрения сложного глаза насекомых. Полученная величина давала основание считать, что этот глаз способен различать два источника света лишь в том случае, если идущие от них лучи попадают в два соседних омматидия. При этом полагали, что свет проникает в омматидий только тогда, когда распространяется практически параллельно оси омматидия.
К несчастью для классической теории мозаичного зрения, эти эксперименты показали, что ценность метода условных рефлексов при исследовании органов чувств весьма ограниченна. Позднее, когда была разработана методика регистрации с помощью микроэлектродов нервных импульсов, возникающих в отдельных омматидиях, исследователям стало ясно, что омматидий представляет собой нечто более сложное, чем простую трубочку. Электрофизиологические эксперименты показали, что «сектор обзора», в пределах которого каждый омматидий чувствителен к световым лучам, составляет 20…30°, а не 2…3°, как утверждала классическая теория (фиг. 21, Б). В то же время омматидий способен различать два луча, идущих под углом 1/3° друг к другу, тогда как из классической теории следовало, что глаза насекомых способны различать два луча, если угол между ними составляет не менее одного градуса, т. е. если каждый луч возбуждает отдельный омматидий. Итак, омматидий получает свет от большей области окружающего пространства, чем предполагали раньше, а сложный глаз обладает большей остротой зрения. Это означает, что каждый омматидий «видит» какое-то изображение, а не просто отличает свет от темноты.
Более того, поля зрения соседних плотно прилегающих друг к другу омматидиев должны перекрываться. Таким образом, классическая теория оказывается несостоятельной; новая же теория, которой предстоит занять ее место, пока что создается довольно медленно. Отчасти это объясняется тем, что создание новых методов и освоение полученной с их помощью поистине необозримой информации требует много времени. Изучая отдельные аспекты функционирования сложного глаза, исследователи обнаруживают все новые тайны, для исследования которых необходимы новые методы.
Более тщательно изучены теперь и вспомогательные структуры сложного глаза. Каждый омматидий собирает информацию от сравнительно большого участка окружающей среды, и отдельное светящееся пятно воспринимается сразу несколькими омматидиями. Таким образом, информация, получаемая соседними омматидиями, в значительной степени перекрывается. Отсюда следует, что клетки ретинулы получают невероятно сложную и запутанную информацию, которая должна быть рассортирована и закодирована в густой сети нервных волокон, расположенных позади ретинальных клеток. Именно эту нервную сеть нужно исследовать, чтобы разгадать секреты зрения насекомых. Весьма возможно, что в результате таких исследований будет обнаружен некий механизм, сортирующий и кодирующий информацию, подобный механизму, который имеется в улитке уха человека, где поступающая в кортиев орган сложная совокупность колебаний преобразуется в закодированное сообщение, состоящее из нервных импульсов.
Между тем остается открытым вопрос: почему в опытах с условными рефлексами, подтвердивших классическую теорию, были получены столь заниженные данные об остроте зрения насекомых? Считалось, что насекомое может различать два источника света лишь в том случае, если угловое расстояние между ними не меньше 1…2°, тогда как позднее с помощью микроэлектродов было показано, что сложный глаз способен различить источники света, расположенные в 10 раз ближе друг к другу. Таким образом, оптическая система и нервные волокна сложного глаза имеют достаточно хорошую разрешающую способность, и вполне вероятно, что возможности зрения насекомых ограничивает центральная нервная система. Быть может, насекомые не могли различать два близко расположенных источника света по той же самой причине, по которой лягушки не могли увидеть мертвых мух: изображение этих мух возникало на сетчатке, но лягушки не «приспособлены» к тому, чтобы на него реагировать (см. гл. 5). И действительно, поведение некоторых насекомых свидетельствует о том, что они обладают исключительно высокой остротой зрения и могут различать мельчайшие детали.
Существует несколько видов ос, которые в отличие от своих общественных собратьев ведут одиночный образ жизни. Эти осы — отличные охотники; они ловко схватывают и жалят свою жертву, а затем складывают добычу в норках, чтобы обеспечить пищей молодое поколение. Некоторые охотящиеся осы-одиночки ловят пауков и пронзают их жалом, прежде чем те успеют нанести ответный удар своими ядовитыми «зубами» — хелицерами; осы другого вида нападают на пчел, возвращающихся в улей. Характерной особенностью всех охотящихся ос является то, что каждый их вид нападает на каких-то определенных животных, инстинктивно предпочитая всем другим определенные виды пауков, мух, пчел, жуков или каких-нибудь других мелких животных. Отсюда естественно вытекает вывод: раз уж они нападают на насекомых только одного определенного вида, значит, они способны распознать свою жертву.
Наблюдать за охотой ос очень легко. В Англии встречаются два вида так называемых роющих ос, которые строят гнезда в песке. Одни из них — полевые бембексы — охотятся на насекомых, собирающихся на коровьем помете, а другие — носатые бембексы (Bembex rostratus) — устраивают засады в соцветиях дикой моркови, купыря и других растений, над которыми носятся тучи мух. Завидев свою жертву, оса подкрадывается к ней и затем, когда до мухи остается всего лишь несколько сантиметров, бросается на нее. Поведение роющих ос во время охоты было предметом очень многих тщательнейших наблюдений, и никто ни разу не видел, чтобы осы пытались напасть на каких-либо других насекомых или приносили в свои гнезда не мух, а насекомых другого вида. Естественно предположить, что осы должны обладать исключительно хорошим зрением; однако это не все. Среди мух, часто встречающихся на белых цветках дикой моркови и подобных ей растений, нередко можно обнаружить журчалок, у которых такое же полосатое брюшко, как у пчел и ос. Всегда считалось, что такая окраска приносит этим безобидным мухам определенную пользу, так как хищники принимают их за жалящих насекомых. Однако бембексы не дают сбить себя с толку: они ловят журчалок и не трогают пчел и ос. Это еще раз свидетельствует о том, что осы прекрасно «узнают» интересующие их объекты.
Все эти наблюдения над роющими осами в естественной обстановке подтверждают результаты лабораторных исследований, показавших, что сложные глаза насекомых намного чувствительнее, чем считалось раньше. К сожалению, пока еще невозможно дать исчерпывающее описание физиологического механизма функционирования сложного глаза. Вероятно, пройдут годы, прежде чем долгий и кропотливый труд исследователей приведет к созданию новой теории; этой последней, по-видимому, будет недоставать простоты мозаичной теории, делавшей сложный глаз столь подходящим объектом для популярных лекций по биологии.
Не у всех насекомых такое же хорошее зрение, как у роющих ос или стрекоз. У многих глаза состоят всего из нескольких омматидиев и могут лишь отличать свет от темноты или воспринимать простейшие геометрические формы; в качестве примера приведем гусениц бабочек-совок, обитающих на юге Англии и ведущих ночной образ жизни. Эти гусеницы живут на деревьях, питаясь листвой дубов, яблонь или хвоей сосен. Иногда они падают с веток и, если это случается, сразу же направляются к дереву и вновь взбираются вверх по его стволу. Не торопитесь, однако, делать поспешные выводы о высоких умственных способностях этих гусениц, которые якобы знают, куда им следует ползти: если вы стоите поблизости, наблюдая за ними, они с той же вероятностью устремятся к вашим ногам и начнут карабкаться вверх. Они будут взбираться вверх и по палке, воткнутой в землю. Возвращением гусениц к пище управляет очень простой физиологический механизм, который при обычном течении событий является вполне адекватным. Каждый глаз гусеницы представляет собой группу из 6 отделенных друг от друга омматидиев, называемых стеммами. Конечно, такие глаза не могут сформировать удовлетворительное изображение окружающего гусеницу мира, однако они способны очень хорошо различать вертикальные края предметов. Когда гусеница поворачивает голову из стороны в сторону, граница светлого и темного, образуемая вертикальным краем предмета, перемещается в поле зрения стемматов, стимулируя их один за другим. Чем шире объект, тем большее воздействие оказывает он на гусеницу; поэтому она ползет к ближайшему дереву, а не растрачивает попусту свои силы на то, чтобы добраться до более отдаленного; кроме того, она предпочитает взбираться на толстые деревья, а не на тонкие палки.
Бабочка колибри-бражник также реагирует только на очень простые признаки предметов. Осенью эти бабочки подыскивают подходящие щели, забираются в них и впадают в спячку. Мы воспринимаем щель как некоторую трехмерную совокупность линий и теней, а для бражника это просто темный объект, который кажется ему подходящим убежищем, если его поперечник составляет примерно 5 см.
На этих двух примерах мы видим, какие простые и специфичные особенности мира насекомых определяют их поведение. То многообразие зрительных деталей, которое необходимо нам и доставляет нам удовольствие, оказывается ненужным и бесполезным для насекомых, так же как и для лягушки с ее «детектором насекомых», поскольку простая нервная система этих животных оказалась бы перегруженной, если бы не отсеивались излишние подробности.
До сих пор мы рассматривали лишь один аспект зрения насекомых: восприятие формы предметов; однако уже давно известно, что некоторые насекомые различают цвета и что диапазон воспринимаемых ими цветов отличается от нашего. Более 80 лет назад было экспериментально доказано, что насекомые видят ультрафиолетовый свет, который совершенно не воспринимается глазом человека. Проведенный эксперимент отличался чрезвычайной простотой: на площадку, где стояла бутыль с сероуглеродом, выпускали муравьев одного из тех видов, которые предпочитают жить в темноте. Для нас сероуглерод — светлая прозрачная жидкость, но поскольку он поглощает ультрафиолетовый свет, муравьям он казался темным, и они собирались около бутыли. Мы не можем даже представить себе, что это такое — видеть ультрафиолетовый свет; возможно, что весь мир окрашивается при этом в темно-пурпурные тона. Однако не только эта особенность отличает цветовое зрение насекомых от нашего. Если бы мы представили себе, как выглядит мир насекомых, нам показалось бы, что тот, кто его раскрасил, взялся совершенно не за свое дело.
Видимый насекомыми спектр цветов был определен в классических экспериментах Карла Фриша. Этот исследователь приобрел известность благодаря своим опытам на пчелах; Фриш обнаружил, что возвращающиеся с нектаром пчелы сообщают своим соплеменницам о направлении к источнику нектара и о расстоянии до него при помощи своеобразного танца на сотах, траектория которого напоминает восьмерку. Опыты по изучению цветового зрения Фриш также проводил на пчелах. Он приучал их кормиться сахарным сиропом из чашечки, поставленной на лист синего картона. После того как у пчел вырабатывался условный рефлекс, они прилетали на синий картон даже тогда, когда на нем не было сахарного сиропа. Если вокруг синего листа картона раскладывали серые листы различных оттенков, пчелы все равно продолжали садиться на синий лист; это доказывает, что они реагировали именно на цвет, а не просто на интенсивность отражаемого куском картона света. Последовательно приучая пчел реагировать на различно окрашенные листы картона, удалось обнаружить, что пчелы способны различать шесть цветов: ультрафиолетовый, голубовато-зеленый, фиолетовый, «пчелиный» пурпурный, желтый и синий. Эти шесть цветов составлены из трех основных цветов — ультрафиолетового, желтого и синего. На фиг. 22 схематически представлены видимые спектры пчелы и человека.
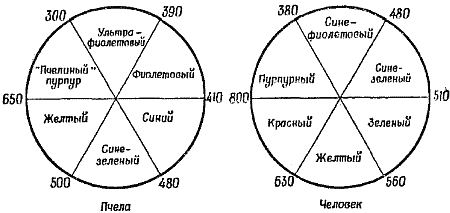
Фиг. 22. Круговые диаграммы цветового зрения пчелы и человека
Первичными цветами для пчелы являются ультрафиолетовый, желтый и голубой, а для человека — сине-фиолетовый, красный и зеленый. Промежуточные цвета образуются при смешивании двух первичных. Цифрами обозначены длины волн в миллимикронах. По сравнению с человеком пчела может видеть цвета в более коротковолновой части спектра.
Видимая пчелами часть спектра смещена по отношению к нашей в сторону более коротких волн. Они видят ультрафиолетовый свет, но не видят красный. Однако некоторые насекомые, особенно жуки-светляки и дневные бабочки (например, черепаховки), видят красный цвет, тогда как для других красный — то же самое, что и черный.
Видимую для насекомых часть спектра можно определить либо косвенным способом в опытах с условными рефлексами, либо непосредственно — измеряя число нервных импульсов, приходящих от клеток ретинулы глаза в ответ на освещение его лучами различного цвета; в результате можно построить такие же диаграммы, как на фиг. 22. Однако эти опыты вряд ли дают нам точное представление о том, как видят мир пчелы или бабочки. Особенно осторожно нужно относиться к результатам экспериментов с условными рефлексами. Капустница выбирает место для кладки яиц, «барабаня» передними ножками по поверхности зеленого листа. Если предлагать ей разноцветные листки бумаги, она будет барабанить по зеленому или голубовато-зеленому, т. е. по листку, окрашенному под цвет листвы. Вместе с тем капустницы питаются нектаром цветков, и такие же движения, как на цветках, они совершают на красных, желтых, синих и фиолетовых листках бумаги, но зеленый цвет или любые оттенки серого не вызывают у них никакой реакции. Отсюда ясно, что, используя сахарный сироп в качестве подкрепления, в экспериментах с условными рефлексами мы не получим адекватного представления о цветовом зрении капустницы.
Окраска цветков тесно связана с наличием у насекомых цветового зрения. Считается, что окрашенные, выделяющие нектар цветки появились на земле примерно в то же время, когда появились летающие насекомые. Насекомые прилетают за нектаром и, переходя с цветка на цветок, переносят пыльцу. Окрашенные венчики возникли у цветков в процессе эволюции потому, что они привлекают насекомых. Однако некоторых насекомых привлекает цветочный аромат, а нередко именно слабо окрашенные цветки обладают если и не очень приятным, то по крайней мере сильным запахом.
Решая вопрос о том, окрашен цветок или нет, нам следует помнить, что насекомые иначе, чем мы, воспринимают его окраску, особенно если он отражает ультрафиолетовый свет. Маки кажутся нам красными, но пчелы не видят красного цвета; поэтому им маки представляются не красными, а, если можно так сказать, «ультрафиолетовыми». Белые цветки обычно воспринимаются пчелами как голубовато-зеленые, поскольку они не отражают ультрафиолетовых лучей, а те цветы, которые кажутся нам синими или фиолетовыми, для пчел могут иметь различные оттенки — в зависимости от того, какое количество ультрафиолетовых лучей они отражают.
Окраска некоторых цветков не только привлекает насекомых своей яркостью, но и указывает им направление к расположенному в нижней части цветка нектарнику. Для этого на лепестках имеются радиально расходящиеся линии, контрастные по отношению к общей окраске цветка; той же цели служит расположенный в центре цветка узор в виде так называемого «бычьего глаза». Эти рисунки, известные как указатели меда, служат насекомым ориентирами. Если учесть, что пчелы воспринимают окраску цветков не так, как мы, то мы не удивимся, обнаружив на фотографиях, сделанных в ультрафиолетовом свете, особые узоры на цветках, кажущихся нам окрашенными однотонно. На фото XI, A показаны два цветка лапчатки, которые мы воспринимаем как равномерно желтые, а на фото XI, Б показаны те же самые цветки, сфотографированные в ультрафиолетовом свете, с ясно различимым указателем меда в виде «бычьего глаза» в центре цветка.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Родопсин и зрение
Родопсин и зрение Лаборатория погружена во мрак. Лишь в двух углах большого помещения, заставленного стеллажами с приборами, слабо лучатся красным светом фонари, которые привычнее было бы видеть в комнате фотографа. Привыкнув к темноте, начинаешь различать лица людей,
Зрение
Зрение Зрение — первейшее чувство у многих животных, особенно у птиц, рыб, осминогов, каракатиц, кальмаров, обезьян и у человека, потомка последних.Из всех животных, пожалуй, лучше всего изучен глаз лягушки. Его сетчатка, получив зрительную информацию, тут же ее оценивает и
ГЛАВА 2. Рентгеновское зрение
ГЛАВА 2. Рентгеновское зрение — Теперь смотрите, одновременно крикнули зебра и жираф. — Вы хотите знать, как это бывает? Раз-два-три! Где же ваш завтрак? Леопард смотрел, и эфиоп смотрел, но они видели только полосатые и пятнистые тени в лесу, но никаких признаков зебры или
Глава 7. Зрение
Глава 7. Зрение Зрение — явление редкое. Глаза, по крайней мере, в общепринятом смысле, отсутствуют у растений, а также у грибов, водорослей и бактерий. Даже среди животных глазами обладают далеко не все. Царство животных делят на тридцать восемь типов, каждому из которых
Глава 18 Зрение днем
Глава 18 Зрение днем Среди посетителей смотровой галереи на самом верху Эмпайр Стейт Билдинг мы видели недавно двух юношей, которые спорили между собой о марках и моделях автомашин, проезжавших внизу по улице, по дну городского каньона. Отличить мужчину от женщины среди
Глава 19 Ночное зрение
Глава 19 Ночное зрение Найдется ли сегодня хоть один цивилизованный человек, который с наступлением сумерек, когда становится слишком темно, чтобы читать газету, не скажет: «Давайте включим свет». При этом он не заметит, что в природе наступил самый волнующий час — ночная
Зрение
Зрение Зрительная система реагирует на световые раздражители. В физическом смысле свет — это электромагнитное излучение с различными длинами волн, от сравнительно коротких (красный) до более длинных (синий). Мы видим объекты потому, что они отражают свет. Цвета, которые
Глава I О свете
Глава I О свете Поведение человека в жизни зависит от чувств, среди которых чувство зрения – наиболее разностороннее и благородное; несомненно, что изобретения, служащие для его усиления, являются самыми полезными из всех остальных. Трудно найти другое изобретение, в
Глава 4. Некоторые прикладные аспекты в свете теорий сбалансированного и адекватного питания
Глава 4. Некоторые прикладные аспекты в свете теорий сбалансированного и адекватного питания 4.1. Вводные замечания Наращивание производства продуктов питания — важнейшая задача, стоящая перед человечеством. Но это лишь одна сторона дела. Другая, не менее важная,
Глава 9. Трофические аспекты происхождения жизни в свете трофологии. Некоторые биологические аспекты трофологии
Глава 9. Трофические аспекты происхождения жизни в свете трофологии. Некоторые биологические аспекты трофологии 9.1. Вводные замечания Из предшествующего изложения ясно, что трофология в целом останется собранием прикладных направлений, если не будут учтены многие
Глава 4. Некоторые прикладные аспекты в свете теорий сбалансированного и адекватного питания
Глава 4. Некоторые прикладные аспекты в свете теорий сбалансированного и адекватного питания 4.1. Вводные замечания Наращивание производства продуктов питания — важнейшая задача, стоящая перед человечеством. Но это лишь одна сторона дела. Другая, не менее важная,
Глава 9. Трофические аспекты происхождения жизни в свете трофологии. Некоторые биологические аспекты трофологии
Глава 9. Трофические аспекты происхождения жизни в свете трофологии. Некоторые биологические аспекты трофологии 9.1. Вводные замечания Из предшествующего изложения ясно, что трофология в целом останется собранием прикладных направлений, если не будут учтены многие
Зрение
Зрение Тремя путями воспринимают свет насекомые: всей поверхностью тела, простыми глазками и сложными, так называемыми фасеточными, глазами.Как показали опыты, всей поверхностью тела чувствуют свет гусеницы, личинки водяных жуков, тли, жуки (даже слепые пещерные), мучные
Глава 9. Зрение{9}
Глава 9. Зрение{9} Только однажды за все годы моей научной работы мне довелось найти глаз ископаемого животного. Это произошло не в палеонтологической экспедиции, а в подсобном помещении лавки минералов в небольшом городке на северо-востоке Китая. Мы с моим коллегой Гао