Зрение
Зрение
Зрительная система реагирует на световые раздражители. В физическом смысле свет — это электромагнитное излучение с различными длинами волн, от сравнительно коротких (красный) до более длинных (синий). Мы видим объекты потому, что они отражают свет. Цвета, которые мы различаем, определяются тем, какую из частей видимого светового спектра отражает или поглощает предмет.
Немецкий физик Герман Гельмгольц, изучавший во второй половине прошлого века глаза животных, установил, что зрительная информация отображается на сетчатке точно так же, как и в любой простой камере с линзой: глаз создает перевернутое и уменьшенное изображение предметов. С этих простых сведений началось накопление того богатства знаний о зрительной системе, которым мы сейчас располагаем. Действительно, мы куда лучше понимаем, как реконструируется зрительный образ окружающего нас мира, чем то, как интерпретируется любая другая сенсорная информация.
Прежде чем познакомиться со структурой и функциями зрительной системы, мы должны сначала рассмотреть, как устроены отдельные ее компоненты. Затем мы проследим за процессом переработки внешних стимулов нейронами различных интегрирующих уровней. И наконец, мы познакомимся с некоторыми выводами психологов о том, как мы видим мир.
Строение зрительной системы
Основные структурные компоненты зрительной системы (рис. 44) — это 1) глаз, в котором наиболее важны части, связанные с фокусировкой изображения и его рецепцией; 2) зрительные нервы, передающие зрительную информацию выходных нейронов сетчатки ядрам таламуса и гипоталамуса; 3) три пары ядер — латеральные коленчатые тела, верхние бугорки четверохолмия (в таламусе) и супрахиазменные ядра гипоталамуса; 4) первичная зрительная кора, которая получает информацию от таламических ядер. Из первичной зрительной коры информация затем поступает в другие области коры, связанные со зрением.
Глаз. Глаз — у млекопитающих единственный орган, специально приспособленный для фоторецепции. Он состоит из «камеры» и собственно фоторецепторного органа. Из частей камеры следует упомянуть: 1) роговицу — тонкую изогнутую прозрачную оболочку, с которой начинается процесс фокусирования световых лучей; 2) хрусталик — линзу, которая завершает этот процесс; 3) радужную оболочку — круговую мышцу, которая изменяет количество попадающего в глаз света, расширяя или сужая отверстие, находящееся в ее центре,- зрачок.
Хрусталик подвешен, как гамак, внутри своей подвижной капсулы. Если мышцы, удерживающие хрусталик, сокращаются или расслабляются, то это изменяет натяжение капсулы, а в результате и кривизну хрусталика. Изменение фокусирующей способности хрусталика обусловлено тем, что он может становиться более плоским или более выпуклым в зависимости от расстояния между объектом и зрителем; такое приспособление называется аккомодацией.
Размеры зрачка — отверстия в радужной оболочке -тоже влияют на то, что и как мы видим. Понаблюдайте за вашим другом, разглядывающим какой-нибудь предмет. Когда он подносит его к глазам, зрачок сужается. Уменьшенный размер зрачка не дает лучам света проходить через хрусталик далеко от его центра и позволяет получить более четкое изображение. Теперь попросите своего друга закрыть глаза на полминуты или около того, а затем вновь открыть их. С близкого расстояния вы увидите, что зрачки, довольно сильно расширенные после того, как ваш друг открыл глаза, тотчас сузились, чтобы приспособиться к освещению в комнате. Автоматический контроль за изменениями в размерах зрачка осуществляют нервные волокна, оканчивающиеся в непроизвольной мускулатуре радужной оболочки.
Некоторым людям нужны очки, чтобы хорошо видеть. Это связано с тем, что аккомодация хрусталика оказывается недостаточной, если сетчатка расположена слишком близко или слишком далеко от задней поверхности хрусталика. Глаз, в котором расстояние между хрусталиком и сетчаткой слишком велико, может фокусироваться только на близких предметах. Такой дефект мы называем близорукостью (миопией). Глаз, в котором сетчатка расположена слишком близко к хрусталику, хорошо фокусируется на далеких, но не на близких предметах. Это дальнозоркость (гиперметропия). По мере того как человек стареет, хрусталик становится более жестким и мышцы уже не могут осуществлять необходимую аккомодацию; тогда самые ближние точки, на которых может фокусироваться глаз, удаляются от него все больше и больше. Когда оказывается, что для ясного видения у человека «слишком короткие руки», он надевает очки и все снова приходит в порядок.
Астигматизм, или искажение зрительных изображений, связанное с неправильной кривизной роговицы, не имеет ничего общего с нарушением расстояния от хрусталика до сетчатки. Для исправления астигматизма весьма пригодны контактные линзы - как бы плавая над поверхностью роговицы в слое слезной жидкости, они компенсируют отклонение ее от правильной формы.
Часть глаза, воспринимающая изображение, — это сетчатка. На первый взгляд может показаться, что сетчатка устроена совсем не так, как нужно. Фоторецепторные клетки — палочки и колбочки — не только расположены в слое, наиболее удаленном от хрусталика, но и повернуты от пучка падающего света, так что их светочувствительные кончики засунуты в промежутки между темноокрашенными эпителиальными клетками.
Под микроскопом видна высокоорганизованная слоистая структура сетчатки (см. рис. 44). Здесь можно различить пять типов нейронов, каждый из которых размещается в пределах своего специфического слоя. Палочки и колбочки (1) соединены с биполярными нейронами (2), которые в свою очередь связаны с ганглиозными клетками (3), посылающими свои аксоны в составе зрительного нерва к вставочным нейронам мозга. Каждая палочка и каждая колбочка соединена с несколькими биполярными клетками, а каждая биполярная — с несколькими ганглиозными. Эта иерархическая структура обеспечивает дивергирующую переработку первичного сигнала, повышающую вероятность его обнаружения. В сетчатке имеются также два типа тормозных нейронов, включенных в локальные сети: горизонтальные клетки (4) и амакриновые клетки (5). Они ограничивают распространение зрительного сигнала внутри сетчатки.
Если с помощью тончайших электродов регистрировать активность отдельных ганглиозных клеток в то время, когда пятно света проходит по сетчатке, мы увидим, что каждая ганглиозная клетка имеет собственное рецептивное поле — небольшой участок сетчатки, в пределах которого свет оказывает наиболее интенсивное возбуждающее или тормозящее влияние на данную клетку. Имеются ганглиозные клетки двух типов — с on-центром и с off-центром. Клетки с on-центром возбуждаются светом, падающим в центр рецептивного поля, но затормаживаются, если свет падает на его периферию. На свет, падающий вне рецептивного поля, клетка вообще не реагирует. Ганглиозная клетка с off-центром затормаживается светом в центре поля, но возбуждается, если свет падает на его края. Синаптические взаимодействия между таламическими интегрирующими нейронами, связанными с ганглиозными клетками того и другого типа, обеспечивают контрастность деталей, которая так важна для четкого видения предметов.
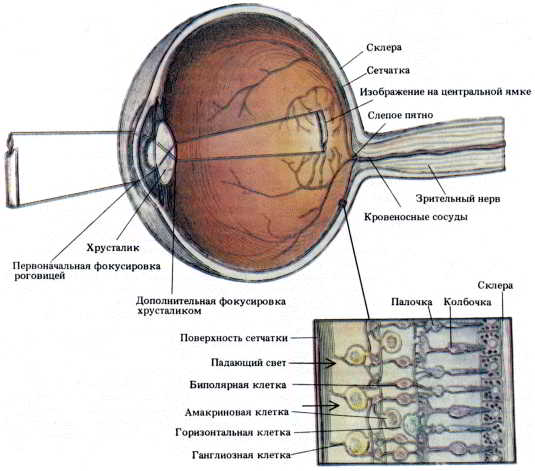
Рис. 44. Глаз в поперечном разрезе. Изображены хрусталик и поддерживающий его аппарат, радужная оболочка и сетчатка с ее светочувствительными детекторами. Внизу показаны детали организации клеток внутри сетчатки. Здесь видно, что первичные рецепторы — палочки и колбочки — расположены в слое, наиболее далеком от падающего света. Сигналы от первичных рецепторов конвергируют на биполярные клетки, а сигналы от биполярных клеток — на ганглиозные клетки. Аксоны ганглиозных клеток сходятся к месту отхождения зрительного нерва (слепому пятну), где рецепторов нет. Эти аксоны образуют зрительный нерв, который передает информацию в мозг. Локальные сети из горизонтальных и амакриновых клеток могут распространять или ограничивать волну активности, исходящую от регистрируемых первичными рецепторами изображений.

Рис. 45. Для того чтобы обнаружить слепое пятно правого глаза, закройте левый глаз и смотрите на черную точку, расположенную слева, постепенно приближая к себе рисунок. Когда фигура приблизится к глазам на расстояние около 25 см, король Чарлз «потеряет голову». (Из Rushton.)
Распределение палочек и колбочек во внутреннем слое сетчатки тоже организовано определенным образом. Колбочки сосредоточены в той части сетчатки, где изображение наиболее четко фокусируется роговицей и хрусталиком. Это место, где острота зрения максимальна, называется центральной ямкой. На этом маленьком участке нет других видов клеток, и на поперечном срезе насыщенная колбочками ямка выглядит как небольшое углубление. Колбочки реагируют на различные цвета; одни чувствительны главным образом к синему цвету, другие — к красному, третьи — к желтому. За пределами центральной ямки колбочки в небольшом количестве равномерно распределены по всей сетчатке.
Палочки чувствительны к яркости отраженного света, но не к цвету. Располагаясь плотнее всего по краям центральной ямки, они в большем количестве, чем колбочки, встречаются и в остальной сетчатке.
Нормальная слоистая структура сетчатки начинается сразу же за пределами ямки. Слой, первым лежащий на пути световых лучей, состоит из аксонов ганглиозных клеток. Эти аксоны со всей сетчатки сходятся в одном месте в стороне от центральной ямки и образуют пучок — зрительный нерв, передающий зрительную информацию мозгу. Там, где собираются аксоны ганглиозных клеток, не остается места для каких-либо рецепторов или других нейронов. Поэтому свет, падающий на сетчатку в этом месте, остается невидимым. Мы никогда не догадываемся о существовании этой «дыры», или «слепого пятна», так как высшие зрительные центры помогают нам заполнить пробел в сенсорной информации. Однако рис. 45 убедит вас, что слепое пятно все же существует.
Зрительный нерв и зрительный тракт. Аксоны ганглиозных клеток, собранные в зрительном нерве, направляются к основанию передней части гипоталамуса, где оба нерва сходятся вместе, образуя хиазму (перекрест). Здесь происходит частичный обмен волокнами с разделением их на перекрещивающиеся и неперекрещивающиеся пучки. Дальше зрительные пути снова расходятся в виде правого и левого зрительных трактов.
Представьте себе, что вы смотрите на зрительную систему человека сверху. С этой удобной позиции вы могли бы увидеть, что все аксоны ганглиозных клеток с той половины сетчатки, которая ближе к носу, переходят в области хиазмы на противоположную сторону. В результате информация ибо всем, что проецируется на внутреннюю (носовую) половину сетчатки левого глаза, переходит в правый зрительный тракт, а о том, что проецируется на носовую часть сетчатки правого глаза, — в левый зрительный тракт (см. рис. 46). Информация же от наружных (височных) половин обеих сетчаток идет по неперекрещенным путям. После хиазмы все стимулы, относящиеся к левой стороне внешнего мира, воспринимаются правой половиной зрительной системы, и наоборот.
Объединение аксонов зрительных нервов в зрительный тракт носит не случайный характер. Волокна перекрещиваются таким образом, что аксоны из соответственных участков обеих сетчаток встречаются и вместе направляются к таламусу. Когда вы смотрите прямо перед собой, все предметы, не находящиеся на средней вертикали, попадают на рецептивные поля клеток носовой (внутренней) половины сетчатки одного глаза и височной (наружной) половины сетчатки другого глаза. Таким образом, каждая точка внешнего пространства проецируется на соответственные (корреспондирующие) точки обеих сетчаток. Дальнейшие отображения всей совокупности таких точек в зрительной системе называются ретинотопическими проекциями поля зрения (рис. 47). Ретинотопическая организация характерна для всей структуры зрительной системы.
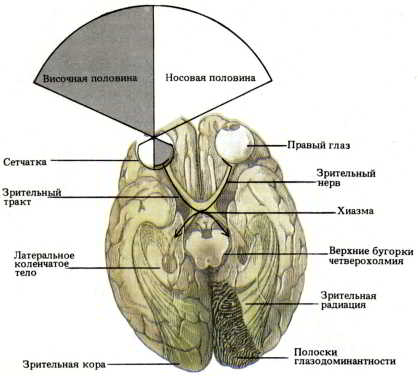
Рис. 46. Схематическое изображение зрительных путей (вид сверху). Образы, воспринимаемые палочками и колбочками носовой (внутренней) половины каждой сетчатки, передаются ганглиозным клеткам, аксоны которых перекрещиваются в хиазме. Образы, воспринимаемые рецепторами височной (наружной) половины каждой сетчатки, передаются ганглиозным клеткам, аксоны которых не перекрещиваются. Таким образом, правая сторона зрительной системы получает информацию об объектах, расположенных слева от средней линии, и наоборот.
Аксоны зрительного тракта подходят к одному из четырех воспринимающих и интегрирующих центров второго порядка. Ядра латерального коленчатого тела и верхних бугорков четверохолмия — это структуры-мишени, наиболее важные для осуществления зрительной функции. (Эти таламические образования, соответствующие в нашей «географической» схеме населенным пунктам, получили названия за свой внешний вид — такими они представлялись ранним исследователям мозга.) Коленчатые тела образуют «коленоподобный» изгиб, и одно из них — латеральное (т. е. лежащее дальше от срединной плоскости мозга) — связано со зрением. Бугорки четверохолмия — это два парных возвышения на поверхности таламуса, из которых верхние имеют дело со зрением. Третья структура — супрахиазменные ядра гипоталамуса (они расположены над зрительным перекрестом) — используют информацию об интенсивности света для координации наших внутренних ритмов (см. гл. 5). И наконец, глазодвигательные ядра координируют движения глаз, когда мы смотрим на движущиеся предметы.
Латеральное коленчатое ядро. Аксоны ганглиозных клеток образуют синапсы с клетками латерального коленчатого тела таким образом, что там восстанавливается отображение соответствующей половины поля зрения. Эти клетки в свою очередь посылают аксоны к клеткам первичной зрительной коры — зоны в затылочной доле коры, имеющей, согласно нашей схеме, статус «государства».
Верхние бугорки четверохолмия. Сейчас мы подошли к очень интересной и важной анатомической особенности зрительной системы. Многие аксоны ганглиозных клеток ветвятся, прежде чем достичь латерального коленчатого ядра. В то время как одна ветвь соединяет сетчатку с этим ядром, другая идет к одному из нейронов вторичного уровня в верхнем бугорке четверохолмия. В результате такого ветвления создаются два параллельных пути от ганглиозных клеток сетчатки к двум различным центрам таламуса. При этом обе ветви сохраняют свою ретинотопическую специфику, т. е. приходят в пункты, в совокупности образующие упорядоченную проекцию сетчатки. Нейроны верхнего бугорка, получающие сигналы от сетчатки, посылают свои аксоны к крупному ядру в таламусе, называемому подушкой. Это ядро становится все крупнее в ряду млекопитающих по мере усложнения их мозга и достигает наибольшего развития у человека. Крупные размеры этого образования позволяют думать, что оно выполняет у человека какие-то особые функции, однако истинная его роль пока остается неясной.
Наряду с первичными зрительными сигналами нейроны верхних бугорков получают информацию о звуках, исходящих от определенных источников, и о положении головы, а также переработанную зрительную информацию, возвращающуюся по петле обратной связи от нейронов первичной зрительной коры. На этом основании полагают, что бугорки служат первичными центрами интегрирования информации, используемой нами для пространственной ориентации в меняющемся мире.
Зрительные поля коры большого мозга. Проекции изображений видимого мира от каждого из латеральных коленчатых ядер передаются по волокнам так называемой зрительной радиации в правую и левую части первичной зрительной коры. Однако эти проекции на корковом уровне уже не представляют собой точных отображений внешнего мира. Область коры, получающая информацию от центральной ямки — зоны наивысшей остроты зрения, примерно в 35 раз больше участка, отображающего кружочек той же величины на периферии сетчатки. Таким образом, информация, идущая от центральной ямки, имеет для коры неизмеримо большее значение, чем информация от других частей сетчатки.
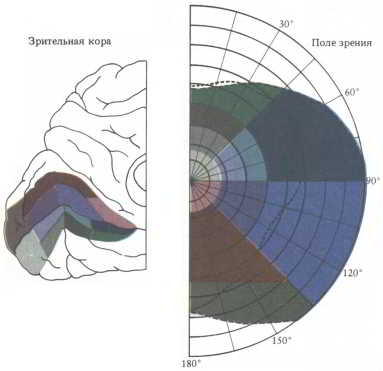
Рис. 47. Когда зрительная информация, получаемая ганглиозными клетками сетчатки, передается первичной зрительной коре, она распределяется там в соответствии с локализацией ее источников в сетчатке. Образы, воспринимаемые в районе центральной ямки — в зоне максимальной плотности палочек и наивысшей остроты зрения, -проецируются на значительно большую область зрительной коры, чем образы, воспринимаемые периферией сетчатки.

Рис. 48. Различная плотность нейронов внутри слоев зрительной коры (вид под микроскопом). У поверхности коры нейронов сравнительно мало. Дальше можно видеть по меньшей мере три главных слоя, а опытный микроморфолог выявит шесть главных слоев.
Первичную зрительную кору называют также «полем 17» или «стриарной корой». Она состоит из весьма упорядоченных слоев и представляет собой структуру, уникальную по своей сложности во всей нервной системе. Для всей коры большого мозга характерна слоистая структура, состоящая, как правило, из шести слоев — от I до VI, начиная с внешней поверхности. Слои различаются по количеству содержащихся в них нейронов. Однако в зрительной коре человека и обезьян эти слои в свою очередь подразделяются, что особенно характерно для слоев IV и V. У приматов можно выявить более 12 слоев зрительной коры, причем слой IV, например, состоит из подслоев IVa, IVb и IVc, в которых опытный глаз гистолога может уловить дальнейшее подразделение.
Другие зрительные зоны коры
Изучая тонкую слоистую структуру коры и распределение в ней клеток и волокон, ученые смогли получить важные сведения о том, какие еще корковые зоны участвуют в дальнейшей переработке зрительной информации. Обнаруженные при этом связи указывают на ряд важных принципов организации зрительных функций коры.
Как показали наблюдения над больными с травмами головы, а также экспериментальные исследования на животных, области коры, связанные со зрением, не ограничиваются первичной зрительной корой. С помощью специальных методик удалось проследить связи от клеток поля 17 к специфическим клеткам слоя IV тех областей, которые лежат в непосредственной близости к полю 17. Эти зрительные зоны называют полями 18 и 19, «престриарной» или вторичной зрительной корой. Однако зрительные пути на этом не заканчиваются. Клетки полей 18 и 19 передают информацию специфическим клеткам некоторых других областей коры большого мозга; кроме того, от них идут связи к зрительным интегрирующим центрам более низкого уровня — таким, как подушка таламуса.
Участки коры, в которых происходит переработка зрительной информации, взаимосвязаны. Было показано, что у обезьян вся затылочная зона и больше половины височной зоны коры содержат зрительные нейронные сети. Изучая характер связей между зрительными полями, ученые смогли сделать некоторые выводы о последовательности операций на «конвейере» переработки зрительной информации. Нейроны коленчатого тела и подушки проецируются в слой IV поля 17, поле 17 — в слой IV полей 18 и 19, а эти поля -обратно в слои I и VI поля 17. Конечно, необходимы новые детальные исследования, чтобы выяснить, насколько типична такая организация связей. Однако тот факт, что определенные закономерности все-таки существуют, сам по себе утешителен.
Изучая таким образом связи между слоями и зонами, исследователи выявили по меньшей мере еще пять уровней интеграции зрительной информации в коре. «Наивысшим» из них оказался уровень, связанный со зрительными полями лобной коры. Они примыкают к так называемой ассоциативной коре, где происходит объединение различных видов сенсорной информации. Возможно, что эта корковая зона имеет прямые связи и с лимбической системой.
Анализ таких сетей наводит на мысль, что выделение каких-то общих зрительных черт, вероятно, происходит на каждом из высших уровней, представленных этими взаимосвязанными зрительными областями коры. Теперь мы подошли к вопросу о том, какие именно элементы видимого мира распознаются и анализируются нейронами первичной зрительной зоны и более высоких уровней. Но прежде, чем ответить на этот вопрос, мы должны рассмотреть некоторые общие особенности кортикальной организации.
Переработка сигналов кортикальными нейронами
Объединение клеток и клеточных связей внутри коры в горизонтальные слои могло бы навести на мысль, что главные взаимодействия в мозгу осуществляются в горизонтальных плоскостях. Однако в 1930-х годах испанский цитолог Рафаэль Лоренте де Но, впервые занявшийся детальным изучением ориентации нейронов коры, высказал предположение, что корковые процессы имеют локальный характер и происходят в пределах вертикальных ансамблей, или колонок, т. е. таких структурных единиц, которые охватывают все слои коры снизу доверху. В начале 60-х годов эта точка зрения получила убедительное подтверждение. Наблюдая реакции кортикальных клеток на сенсорные стимулы при медленном продвижении тонких электродов сквозь толщу коры, американский физиолог Вернон Б. Маунткасл сравнивал характер регистрируемых ответов внутри вертикально организованных структур. Первоначально его исследования касались тех областей коры, где имеется проекция поверхности тела и нейроны реагируют на сигналы от рецепторов, находящихся в коже или под кожей, но в дальнейшем справедливость полученных выводов была подтверждена и для зрительной системы. Главный вывод заключался в том, что сенсорные сигналы, идущие от одного и того же участка, возбуждают группу нейронов, расположенных по вертикали.
Вертикальные колонки нейронов более или менее сходного типа распространены во всей коре больших полушарий, хотя размеры и плотность клеток в них варьируют. Поэтому ученые считают, что переработка информации в коре зависит от того, как эта информация достигает кортикальной зоны и как ее передают связи между клетками внутри данной вертикальной колонки. Продукт деятельности любой такой колонки можно весьма приближенно сравнить с результатами многоступенчатых математических вычислений, при которых одни и те же операции выполняются в одинаковом порядке независимо от того, какие исходные данные были введены: например, «возьмите номер вашего дома, отбросьте последнюю цифру, разделите оставшееся число на 35, округлите частное, и вы получите в ответе номер ближайшей поперечной улицы».
Информация, с которой имеют дело кортикальные колонки, — зрительная для зрительной коры, тактильная для тактильной, слуховая для слуховой и т. д. -конечно, уже была подвергнута частичной переработке первичными воспринимающими и интегрирующими центрами. Результаты деятельности одной корковой колонки с помощью специфических внутрикортикальных синаптических связей передаются затем другой колонке для дальнейшей обработки данных.
Любая корковая колонка содержит примерно одинаковое число клеток — 100 или около того, будь то мозг крысы, кошки, обезьяны или даже человека. Большие способности отдельных особей внутри вида с определенным строением коры обусловлены большим числом колонок в коре и нервных волокон, связывающих их между собой внутри отдельных корковых зон.
Теперь, имея представление о вертикальных связях клеток, входящих в горизонтальные слои коры, мы можем вернуться к рассмотрению специфических клеток зрительной системы.
Избирательная реакция нейронов на отдельные особенности зрительных стимулов
Некоторые ганглиозные клетки сетчатки возбуждаются тогда, когда свет падает в центр их рецептивного поля, и затормаживаются, если он попадает на его периферию; другие — наоборот. Можно сказать, что одни клетки сетчатки положительно реагируют на бублик, а другие — на дырку от него. На сплошное круглое пятно света такие клетки могут вообще никак не ответить, так как тормозящее действие света, попадающего в центр рецептивного поля, уравновешивает активирующий эффект освещения краев.
Эксперименты, проведенные американским физиологом Стивеном Куффлером в середине 50-х годов, показали, почему ученым не удавалось понять, как «видит» сетчатка, если они использовали рассеянный свет. Такой свет стимулировал многие соседние нейроны с рецептивными полями разного типа (с on-центрами, с off-центром и др.), и обусловленное этим «уравнивание» эффекта ослабляло реакцию исследуемых ганглиозных клеток. Но, как обнаружил Куффлер, воздействие небольшими локальными стимулами ведет к хорошо воспроизводимой реакции определенных групп ганглиозных клеток.
Спустя несколько лет Дэвид Хьюбел и Торстен Визель применили тот же метод дискретных стимулов для активации нейронов латерального коленчатого тела у кошки и обезьяны. Ответные реакции оказались весьма сходными с теми, которые наблюдались раньше при изучении рецептивных полей ганглиозных клеток сетчатки. В коленчатом теле тоже выявились нейроны с характерными рецептивными полями, напоминающими по форме маленькие бублики, в которых возбуждающей областью был либо центр, либо периферия. Тормозящий эффект периферии или же центра прямо определялся тем, какая ганглиозная клетка активировала данную клетку коленчатого тела. На основании этих результатов Хьюбел и Визель сделали вывод, что обработка зрительной информации начинается со сравнения количества света, падающего на любой небольшой участок сетчатки, с уровнем света вокруг него. Продвигая электрод вертикально вниз через слои коленчатого тела, они обнаружили ряд клеток, которые возбуждались под действием стимулов из одних и тех же частей поля сетчатки. При этом клетки, лучше реагировавшие на сигналы от правого глаза, располагались непосредственно над или под клетками, «предпочитавшими» информацию от левого глаза.
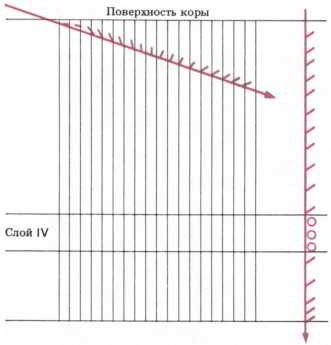
Рис. 49. Ориентация рецептивных полей (указана боковыми штрихами) группы клеток зрительной коры, встретившихся при введении микроэлектрода под малым углом к поверхности в зону выше слоя IV.
Затем Хьюбел и Визель применили те же методы анализа к нейронам слоя IV первичной зрительной коры (поле 17), куда поступает информация от латерального коленчатого тела. И здесь реакции были сходны с реакциями клеток сетчатки и коленчатого тела. Однако нейроны, расположенные выше и ниже слоя IV, почему-то не реагировали на стимулы, соответствующие небольшим бубликообразным рецептивным полям сетчатки. Зрительные раздражители, состоящие из черных точек на белом фоне или наоборот, вызывали лишь слабую и непостоянную реакцию. Чем же объяснялась потеря специфической восприимчивости?
Решить эту загадку случайно помогла реакция одной клетки. Кружки, вызывавшие энергичный ответ в слое IV, практически не стимулировали клеток слоя V, но тонкая черная линия, оказавшаяся на краю поля раздражителя, вдруг вызывала быструю ответную реакцию. Вскоре суть дела стала ясна. Почти все клетки коры, расположенные выше или ниже слоя IV, предпочтительно реагировали на стимулы в форме краев или же светлых или темных линий на контрастном фоне. После того как была выявлена роль формы, дальнейшие исследования показали, что различные нейроны отдают предпочтение линиям, расположенным под разными углами. Некоторые специализированные клетки реагировали только на движущиеся или только на неподвижные линии или «края» (т. е. границы между темными и светлыми участками), другие отвечали на движение в определенном направлении (см. рис. 55, а также рисунки 56 и 57 с относящимся к ним текстом). Отдельные кортикальные клетки, расположенные выше и ниже слоя IV, реагировали на линии той или иной длины, а также на характер контраста (светлая линия на темном фоне или наоборот).
СЛУЧАЙНОЕ ОТКРЫТИЕ
К 1962 г. были разработаны методы регистрации электрической активности ганглиозных клеток сетчатки и клеток латерального коленчатного тела при воздействии стимулов на сетчатку. Работая с наркотизированными кошками, Дэвид Хьюбел и Торстен Визель начали изучать реакцию клеток первичной зрительной коры. Многие клетки слоя IV реагировали подобно клеткам сетчатки и латерального коленчатого тела, проявляя высокую активность, когда на их рецептивные поля падали небольшие пучки света. Однако нейроны, расположенные выше и ниже слоя IV, казалось, вовсе не реагировали на раздражитель до той поры, пока случайное наблюдение не помогло экспериментаторам разгадать их тайну. Предоставим слово Хьюбелу:
«Вначале вы никак не могли добиться разрядов в этих клетках. Мы проецировали пучки света по всему экрану, но это не помогло. И вот однажды, скорее по чистой случайности, мы стали создавать на экране небольшие пятна света и обнаружили, что „работает“ черная точка, но как — мы не могли понять до тех пор, пока не выяснили: виной всему кусок стекла, который мы вставляем в проектор. Он посылал на экран четкую и тонкую, едва заметную линию. Каждый раз, когда это происходило, мы получали ответ. Более того, ответ возникал только в том случае, если линия была ориентирована в определенном направлении, а не в каком-либо другом...»
После этого Хьюбел и Визель начали проверять реакцию клеток верхнего и нижнего слоев зрительной коры на изображения в виде линий, полосок или палочек. Было обнаружено, что эти клетки избирательно реагируют не только на само удлиненное изображение, но и на его ориентацию под определенным углом (см. рис. 49).

Рис. 50. Дэвид Хъюбел и Торстен Визель.

Рис. 51. Кошка в экспериментальном аппарате.
«Простые» клетки коры, подобно ганглиозным клеткам (или нейронам коленчатого тела), могут реагировать только на контраст между центром и окружающей областью. Ответная реакция «сложных» клеток избирательна в отношении таких особенностей, как ориентация, форма контура, перемещение и т. п. «Простые» клетки коры почти наверняка активируются сочетанием возбуждающих и тормозящих влияний, исходящих из соответствующих пунктов коленчатого тела. «Сложные» клетки, очевидно, могут извлекать дополнительную информацию о размерах, форме и движении сигналов.

Рис. 52. Изображения, которые мы встречаем в печати, целиком состоят из точек. При большом увеличении (слева) изображение нельзя «прочитать». При меньших увеличениях точки постепенно сливаются, и в конце концов возникает распознаваемый образ. Так же может происходить и поступление информации в зрительную систему.
Но как из взаимодействий между всеми этими нейронами рождаются те реальные, «телесные» образы, которые мы видим? Если вы взглянете на фотографию в газете через лупу, то обнаружите, что изображение состоит из точек. В темных местах эти точки крупнее и почти сливаются, в светлых — значительно мельче. Когда видишь эти детали сильно увеличенными, вряд ли поймешь, что изображено на картинке. Лишь тогда, когда вы смотрите невооруженным глазом, точки пропадают и появляется изображение. Упрощенно говоря, ответные реакции ганглиозных клеток сетчатки, активируемых ими клеток в коленчатом теле и «простых» клеток зрительной коры составляют систему распознавания точек мозгом (рис. 52).
Линии и прямые края — удобные примеры образов, распознаваемых «сложной» клеткой зрительной коры. Но то, что видят наши глаза, не сводится к этим образам. При более детальном исследовании оказывается, что «сложные» клетки поля 17 реагируют, по-видимому, чуть раньше простых клеток. Значит, мысль о том, что внутри кортикальной колонки информация от «простых» клеток как бы считывается «сложными», несмотря на свою привлекательность, пока не находит подтверждения. Может быть, более верным будет предположение, что нейроны — опознаватели точек в сетчатке и коленчатом теле — служат своего рода «фильтрами» для некоторых видов зрительных стимулов. Когда корковые нейроны получают отфильтрованные данные, мир может «выглядеть» как фото в газете, если смотреть на него с такого расстояния, когда точки уже начинают сливаться, но связное изображение еще не появилось. Линии и края в этом случае можно рассматривать как элементы изображения, контуры и другие особенности которых могут пройти сквозь сито первичных уровней переработки информации.
Два глаза — один мир
Можно описать многие аспекты того, как мы видим, но они еще не получили точного биологического объяснения. Возможно даже, что некоторые стороны этого процесса вообще еще не выявлены. Переработкой зрительной информации занята значительная часть нашего мозга, но насколько велика эта часть, ученые затрудняются сказать даже приблизительно.
Мы знаем, что у нас два глаза, но мы почти всегда видим только один внешний мир. Эта способность объединять информацию, идущую от обоих глаз, основана на двух важнейших свойствах зрительной системы.
Во-первых, движения наших глаз, когда мы осматриваем ими окружающее, сложным образом скоординированы. Если вы, глядя на острый край какого-нибудь предмета, легонько надавите сбоку на глазное яблоко, то в этот миг увидите оба изображения, из которых складывается одно. Для слияния изображений особенно важны нейроны верхних бугорков четверохолмия. Эти клетки лучше реагируют на движущиеся раздражители. Они тоже организованы в вертикальные колонки, клетки которых отвечают на сигналы, идущие из одних и тех же участков поля зрения. Оказалось, что клетки, расположенные в самом низу колонки, активируются непосредственно перед тем, как происходит спонтанное движение глаз. Их активность служит пусковым механизмом для глазодвигательных нейронов; последние вызывают сокращение соответствующих мышц, а те перемещают глаз таким образом, чтобы участок поля зрения, где что-то движется, проецировался на центральную ямку. Так, поворачивая вместе оба глаза, мы «обращаем свое внимание» туда, где блеснула вспышка света или что-то передвинулось, чтобы получше рассмотреть это «что-то».
Клетки, расположенные в глубоких слоях верхних бугорков, получают также слуховую информацию и реагируют на звук. Слуховая информация, объединяющаяся в этих клетках со зрительной, вызывает посылку сигналов на более низкий уровень — клеткам среднего мозга, управляющим мышцами глазного яблока. С помощью этих мышц вы переводите взгляд туда, где, как сообщает ваш слух, в этот момент что-то происходит.
Во-вторых, проекции видимого мира на сетчатках обоих глаз отображаются в поле 17 в виде двух почти идентичных проекций, которые затем объединяются межкорковыми связями каким-то еще не вполне понятным образом. Ученым, однако, известно, что по крайней мере на уровне коленчатого тела и поля 17 благодаря довольно сложной системе проводящих путей зрительная информация от каждого из двух глаз остается пространственно обособленной. У наркотизированных животных клетки слоя IV поля 17 реагируют на импульсы, идущие от обоих глаз. В клетках, расположенных выше и ниже слоя IV, ответные реакции носят более сложный характер. Здесь, как правило, некоторые клетки лучше реагируют на сигналы от одного глаза, чем от другого; иными словами, влияние одного глаза на такие клетки «доминирует» над влиянием другого глаза. Действительно, можно проследить за ходом нервных путей от определенных участков поля зрения одного глаза через связи в коленчатом теле вплоть до зрительной коры. Здесь эти пути подходят к чередующимся «колонкам глазодоминантности», которые формируются на расстояниях примерно 0,4 мм друг от друга и пронизывают всю толщу коры. Если взглянуть сверху на колонки глазодоминантности поля 17, то те из них, которые связаны с одним глазом, сольются в изогнутые гребешки, очень напоминающие кожные узоры на пальцах (см. рис. 53).
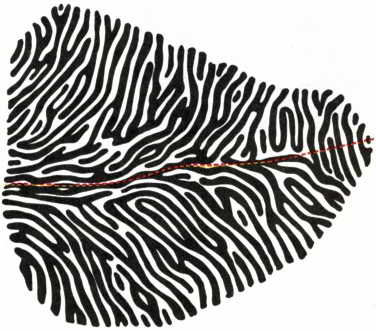
Рис. 53. Узор из колонок глазодоминантности в зрительной коре. Нейроны, максимально реагирующие на сигналы от левого или от правого глаза, образуют чередующиеся гребни (на рисунке белые или черные). При введении электрода под любым углом встречаются клетки того и другого типа. Рисунок гребней, напоминающий отпечатки пальцев, у разных индивидуумов различен.
При изучении этих колонок были выявлены удивительные факты, касающиеся формирования коры. Если один глаз будет закрыт от рождения, то нейроны коленчатого тела, с которыми связаны ганглиозные клетки сетчатки этого глаза, и соответствующие им колонки доминантности в коре не смогут нормально развиваться. И хотя сетчатка закрытого глаза полностью сохранит свою чувствительность (в чем можно убедиться, открыв этот глаз), ее связи не будут обеспечивать полноценной ответной реакции в коленчатом теле или коре. Колонки доминантности, связанные с закрытым глазом, окажутся более узкими, чем в норме. В то же время влияние глаза, функционировавшего с рождения, распространится на значительно большую, чем обычно, область коры. Эти эксперименты показывают, что степень связи между сенсорными нейронами и соответствующими клетками коры может регулироваться уровнем активности сенсорной системы.
Зрительные пути правого и левого глаза могут служить наглядной иллюстрацией параллельных цепей (так же, как и слуховые пути от обоих ушей, если бы мы рассматривали эту систему). Зрительная информация от рецепторных клеток сетчатки каждого глаза идет практически параллельными путями до зрительной коры (см. рис. 46).
Наши два глаза с удвоенными зрительными путями не просто «уравновешивают» лицо или обеспечивают резерв на случай выхода из строя одного глаза. Они работают сообща для достижения суммарного эффекта. Разница в положении глаз обусловливает незначительные различия в идущей параллельными путями зрительной информации, а это в свою очередь позволяет нам видеть предметы в трех измерениях. Когда эта информация объединяется в зрительных интеграционных центрах коры, мы видим один трехмерный мир.
Деятельность других параллельных путей тоже обогащает наше зрительное восприятие. Различные аспекты информации, получаемой от каждого глаза, передаются по трем параллельным каналам. Информация о специфике образа (распознавание «точек») поступает через латеральное коленчатое тело в первичную зрительную кору. Информация, касающаяся движения, по различным аксонам направляется от сетчатки к верхним бугоркам четверохолмия и к полю 17 зрительной коры. Сигналы об уровне рассеянного света идут в супрахиазменные ядра. Вся эта информация, передаваемая по различным, но параллельным путям, в конце концов вновь объединяется в интегрирующих сетях коры и воссоздает полную картину того, что мы видим.
Этот общий принцип разделения первичной информации по отдельным перерабатывающим каналам для последующего ее воссоединения, как мы увидим дальше, широко используется как в сенсорных, так и в двигательных системах.
Цвет — особое качество зрительного ощущения
Цвет — это одно из качеств, которое едва ли нуждается в описании. Каждый знает разницу между черно-белым и цветным кино. Однако о восприятии цвета следует кое-что сказать.
Мы уже упоминали о существовании трех типов колбочек — специализированных цветочувствительных рецепторов сетчатки. Физиологическое отображение цвета начинается именно с этих клеток. Хотя тремя основными цветами обычно считают красный, синий и желтый, ганглиозные клетки дают оптимальную реакцию на красный, синий и зеленый цвета.
Анализ пигментов, содержащихся в колбочках, и прямая регистрация активности этих рецепторов в идеальных экспериментальных условиях подкрепляют представление о том, что для восприятия каждого из трех первичных цветов — красного, желтого и синего — существует особый тип колбочек. Но когда физиологи занялись изучением выходных сигналов от сетчатки и исследовали ответы ганглиозных клеток при воздействии того или иного чистого цвета, ситуация усложнилась и, если хотите, стала более интересной. Эксперименты показали, что ганглиозные клетки и активируемые ими нейроны латерального коленчатого тела реагируют так, как будто существует не три, а четыре первичных цвета: красный, желтый, синий и зеленый. Но если нет таких колбочек, пигмент которых обусловливал бы специфическую реакцию на зеленый цвет, то как объяснить полученные результаты?
Один из ключей к разгадке тайны восприятия зеленого цвета был получен в результате несложных экспериментов. Людей расспрашивали о цветах, которые они видят в определенных условиях. Если смотреть на серое пятно, окруженное ярко-зеленым кольцом, то серый цвет начинает приобретать красноватый оттенок. Если некоторое время фиксировать взором ярко-красный предмет, а потом закрыть глаза, то возникнет так называемый последовательный образ этого предмета, окрашенный в зеленый цвет. Этот хроматический эффект последовательного контраста и есть источник так называемого «зеленого свечения», которое можно увидеть, если пристально смотреть на заходящее солнце. Последовательный образ синего предмета окрашен в желтый цвет (это легче увидеть, если синий предмет находится на черном фоне).
Таким образом, получается, что синий и желтый цвета, так же как красный и зеленый, как-то связаны между собой. Но эти сопоставления, возможно, не покажутся вам верными. Ведь вы знаете, что для получения зеленого цвета нужно смешать синюю и желтую краски. Как же происходит восприятие зеленого цвета?
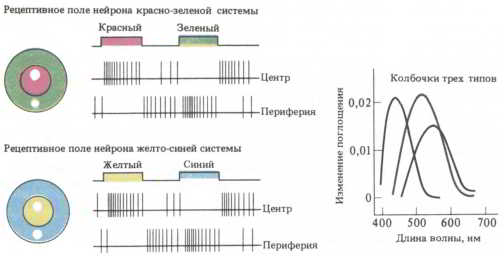
Рис. 54. Возможный способ кодирования цветов в сетчатке. Вверху: ганглиозная клетка активируется красным цветом в центре поля и зеленым на периферии. Внизу: активность ганглиозных клеток, получающих входные сигналы от «желтых» колбочек в центре поля и «синих» — на периферии. У этих цветоспецифических рецептивных полей отмечается антагонизм центра и периферии. (Колбочек, воспринимающих зеленый цвет, не существует. Это качество возникает благодаря конвергенции горизонтальных нейронов локальных сетей в пределах сетчатки.)
Восприятие цвета начинается с того, что специализированные колбочки распознают один из трех первичных цветов. Колбочки связаны с биполярными клетками, а те — с ганглиозными. Решающее значение для восприятия зеленого цвета имеют нейроны локальных сетей сетчатки — горизонтальные клетки.
Одна из теорий, лучше других согласующаяся с экспериментальными данными, носит название теории оппонентных цветов. Она была впервые сформулирована в XIX веке немецким физиологом Эмилем Герингом. По его мнению, некоторые цвета являются «антагонистами»: желтый и синий, красный и зеленый, черный (отсутствие цвета) и белый (сочетание всех цветов). Эксперименты, проведенные спустя сто лет и основанные на регистрации активности отдельных клеток, дали именно те результаты, которых можно было ожидать, исходя из этой теории. Ганглиозные клетки, воспринимающие красный цвет в центре рецептивного поля, на его периферии воспринимают зеленый, и наоборот. Клетки, реагирующие на желтый цвет в центре, чувствительны к синему на периферии, и наоборот. Колбочки активируются светом определенного цвета. Благодаря взаимодействию с горизонтальными клетками происходит комбинирование различных «цветовых» сигналов при их конвергенции на ганглиозные клетки сетчатки. Вот почему ганглиозные клетки распознают цвета-«оппоненты», и зеленый выступает антагонистом красного (см. рис. 54).
Недавние исследования показали, что цветовая специфичность сигналов от сетчатки сохраняется и в зрительной коре. Клетки, расположенные в верхних слоях зрительной коры, обладают цветоспецифическими рецептивными полями и реагируют на цвета-«оппоненты». В то же время они не обнаруживают избирательности по отношению к ориентации линий или краев. На этом основании Дэвид Хьюбел предположил, что система переработки цветовой информации отделена от системы, перерабатывающей ориентационную информацию, но действует параллельно с ней.
Предметное и пространственное зрение
Обычно мы не расчленяем зрительное восприятие на видение одним или двумя глазами, цветное или черно-белое зрение, пока не возникают какие-нибудь неполадки. Большей частью мы просто видим.
Незамеченными остаются и некоторые другие аспекты обработки зрительной информации. Один из таких аспектов заключается в том, что зрительная система позволяет определить, где именно находится в окружающем нас пространстве данный объект и что он собой представляет. До недавнего времени полагали, что эти два процесса протекают раздельно, начиная с довольно ранних стадий обработки входных сигналов. Функции, связанные с пространственной информацией, приписывали верхним буграм четверохолмия, так как именно оттуда исходят команды к мышцам, поворачивающим глаза при фиксации взором объектов. Способность распознавать признаки объекта объясняли их последовательным анализом. Однако проведенные недавно эксперименты показали, что эти представления, вероятно, ошибочны: похоже, что оба вида зрительного анализа зависят от потока информации, идущего от коленчатого тела к полю 17, и от различных систем, которым поле 17 передает эту информацию для дальнейшей обработки.
Распознавание признаков предмета. Недавно была исследована способность обезьян запоминать очень сложные особенности предметов, чтобы получать пищевое вознаграждение. Так, например, животные должны были выбрать деревянный квадрат с нанесенными на него полосами, а не точно такой же предмет, но без полос. После того как обезьяны запоминали отличительный признак, у них удаляли небольшие участки коры в одном из тех мест, где прослеживались пути, участвующие в обработке зрительной информации. Оправившихся после операции животных вновь подвергали тем же тестам. После двустороннего удаления той части височной доли, куда поступает зрительная информация, животное еще могло видеть — оно брало в руки предметы, чтобы получить пищу, но выбирать квадраты с полосками оно уже не могло. Этот хирургический метод в сочетании с выявлением сетей и регистрацией электрической активности показал, что функция «распознавания признаков» связана с участком височной доли у нижнего края коры.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Родопсин и зрение
Родопсин и зрение Лаборатория погружена во мрак. Лишь в двух углах большого помещения, заставленного стеллажами с приборами, слабо лучатся красным светом фонари, которые привычнее было бы видеть в комнате фотографа. Привыкнув к темноте, начинаешь различать лица людей,
ГЛАВА 6 Зрение в ином свете
ГЛАВА 6 Зрение в ином свете Глаза насекомых и других членистоногих представляют собой сложные органы, состоящие из множества одинаковых элементов. В каждом таком элементе, называемом омматидием (фиг. 20), представлены все составные части глаза; соседние омматидии отделены
Зрение
Зрение Зрение — первейшее чувство у многих животных, особенно у птиц, рыб, осминогов, каракатиц, кальмаров, обезьян и у человека, потомка последних.Из всех животных, пожалуй, лучше всего изучен глаз лягушки. Его сетчатка, получив зрительную информацию, тут же ее оценивает и
Обоняние и цветное зрение развивались в противофазе
Обоняние и цветное зрение развивались в противофазе Эволюция цветного зрения у наземных позвоночных хорошо изучена. Способность различать цвета определяется светочувствительными белками колбочек — опсинами, которые могут быть «настроены» на разную длину волны. В
2.16. Цветовое зрение
2.16. Цветовое зрение Весь видимый нами спектр электромагнитных излучений заключён между коротковолновым (длина волны 400 нм) излучением, которое мы называем фиолетовым цветом, и длинноволновым излучением (длина волны 700 нм), называемым красным цветом. Остальные цвета
У кого зрение лучше – у человека или у собаки?
У кого зрение лучше – у человека или у собаки? Ночное зрение у собаки в 3–4 раза лучше, чем у человека, а дневное – примерно в 3 раза ниже, чем у человека.Если бы мы захотели проверить дневное зрение собаки с помощью обычной проверочной таблицы, которая висит в кабинете
ГЛАВА 2. Рентгеновское зрение
ГЛАВА 2. Рентгеновское зрение — Теперь смотрите, одновременно крикнули зебра и жираф. — Вы хотите знать, как это бывает? Раз-два-три! Где же ваш завтрак? Леопард смотрел, и эфиоп смотрел, но они видели только полосатые и пятнистые тени в лесу, но никаких признаков зебры или
Глава 7. Зрение
Глава 7. Зрение Зрение — явление редкое. Глаза, по крайней мере, в общепринятом смысле, отсутствуют у растений, а также у грибов, водорослей и бактерий. Даже среди животных глазами обладают далеко не все. Царство животных делят на тридцать восемь типов, каждому из которых
Слух, зрение и обоняние
Слух, зрение и обоняние Муравей сам по себе существо мудрое... Ф. Бэкон. Как муравьи видят. Часто человек судит о других по себе, и поэтому, например, наблюдая муравья, удивляется: «Вот какой глупый, не обращает внимания на мертвую муху». Муха лежит от муравья в десяти
Глава 18 Зрение днем
Глава 18 Зрение днем Среди посетителей смотровой галереи на самом верху Эмпайр Стейт Билдинг мы видели недавно двух юношей, которые спорили между собой о марках и моделях автомашин, проезжавших внизу по улице, по дну городского каньона. Отличить мужчину от женщины среди
Глава 19 Ночное зрение
Глава 19 Ночное зрение Найдется ли сегодня хоть один цивилизованный человек, который с наступлением сумерек, когда становится слишком темно, чтобы читать газету, не скажет: «Давайте включим свет». При этом он не заметит, что в природе наступил самый волнующий час — ночная
Зрение
Зрение Зрительная система реагирует на световые раздражители. В физическом смысле свет — это электромагнитное излучение с различными длинами волн, от сравнительно коротких (красный) до более длинных (синий). Мы видим объекты потому, что они отражают свет. Цвета, которые
4.3. Внутреннее духовное зрение и подготовка
4.3. Внутреннее духовное зрение и подготовка Хотя практика пранаямы имеет дело с нашими дыхательными циклами, она не попадает в категорию каких-либо гимнастических упражнений, где дыхание выполняет пассивную функцию, как уже объяснялось в п. 1.7. Пранаяма – это активный
Глава 9. Зрение{9}
Глава 9. Зрение{9} Только однажды за все годы моей научной работы мне довелось найти глаз ископаемого животного. Это произошло не в палеонтологической экспедиции, а в подсобном помещении лавки минералов в небольшом городке на северо-востоке Китая. Мы с моим коллегой Гао