Движение
Движение
«Объединители» и «дробители» этого мира пытаются проникнуть в суть явлений разными путями. Приверженцы классического подхода — объединители — хотят найти то, что объединяет вещи. Они ищут законы, управляющие функциями целого. Мыслители романтического склада — дробители — концентрируют свое внимание на частностях — внешнем виде, звуках и т.п. Классики считают толкования романтиков неприемлемыми («Ты говоришь, что любишь меня, но ведь я — это не только пара больших карих глаз»), а романтиков не устраивают требования классиков («В этой книге о мозге слишком много деталей — лучше прямо скажите, что именно я должен знать»).
Мы поставили своей целью дать классическое толкование того, как организован мозг, и тех принципов, на основе которых, по нашим представлениям, он работает. Но наряду с твердо установленными фактами, структурными схемами и данными о нейронных связях приводятся и взятые из жизни примеры, касающиеся разнообразных проявлений человеческой деятельности. Когда мы перейдем от сенсорных систем к моторным, для понимания того, как мозг заставляет тело двигаться, нам нужно будет использовать оба упомянутых выше подхода.
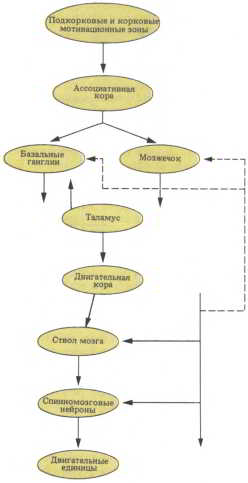
Рис. 56. Основные компоненты двигательной системы. Участки пирамидного тракта соединены иерархическим способом: двигательная кора ? спинной мозг ? мышечный эффектор. Показаны также области параллельной активности, лежащие за пределами пирамидного тракта, и мозжечковые компоненты двигательной системы.
Очень упрощенно мы можем представить себе, что «конвейер», действующий в моторной системе, движется в направлении, противоположном его движению в сенсорных системах. В сенсорной системе информация берет начало на периферии, где она воспринимается сенсорными датчиками и передается вверх в коре мозга. В моторной системе главный поток информации направлен от двигательной коры к периферии, где работают мышечные структуры — эффекторы, осуществляющие движение (рис. 56). Двигательная система тоже включает ряд иерархических уровней и путей параллельной обработки информации, и для эффективности ее работы тоже необходимы сенсорные проекции тела в мозгу. Все это относится к движениям любого типа, будь то простое почесывание носа или элегантный и сложный соскок с параллельных брусьев, во время которого гимнаст выполняет двойное сальто назад с полным переворотом.
Мышцы и суставы
Почти все мышцы нашего тела идут от одной кости к другой через соединяющий их сустав. Когда под действием двигательного нерва мышца сокращается, ее укорочение заставляет одну из этих двух костей двигаться. Есть два исключения из этого «правила двух костей»: глазные мышцы, вызывающие движения глазного яблока, и мышцы языка. Но даже и в этих случаях справедливо другое общее правило: каждой мышце, вызывающей определенное движение (например, сгибание в суставе, поворот глазного яблока, сужение зрачка), противостоит другая, при сокращении которой происходит противоположное движение. Работа таких пар мышц-антагонистов имеет решающее значение для поддержания выпрямленного положения тела или иной позы вопреки действию силы тяжести.
При активации двигательных нервов в нервно-мышечном синапсе происходит выброс химического медиатора ацетилхолина, который передает мышце команду сокращаться. (Многие другие нейроны тоже используют ацетилхолин для передачи информации различным клеткам-мишеням.) На мембране мышечного волокна ацетилхолин связывают специализированные крупные молекулы, называемые рецепторами (это не то, что мы называем рецепторами в сенсорных системах). Воздействие ацетилхолина на рецепторы может быть усилено никотином и, наоборот, блокировано растительным ядом кураре. (Кураре успешно применяется при охоте с помощью лука, так как животное, пораженное стрелой, смоченной в кураре, уже не может убежать.) При заболевании, называемом «миастения гравис», мышцы также утрачивают способность реагировать на ацетилхолин, но по другой причине: разрушаются ацетилхолиновые рецепторы на мышечных клетках.
В большинстве случаев наши мышцы сокращаются лишь тогда, когда мы этого хотим. Поэтому такие движения называются произвольными (см. рис. 8). Но даже тогда, когда мы делаем то движение, которое решили сделать, мы не осознаем, какие мышцы и каким образом в нем участвуют. За редкими исключениями, мы фактически не умеем приводить в действие ту или иную мышцу в отдельности. И тем не менее общий термин «произвольные» отделяет движения этого типа от рефлекторных движений, которые имеют место в тех, например, случаях, когда вы, неосторожно коснувшись раскаленной плиты, отдергиваете руку, еще не успев почувствовать боль.
Независимо от внешней причины двигательной активности мышечные волокна активируются только по приказу двигательного нерва. Поэтому мы говорим о двигательном аксоне и мотонейроне (спинномозговом нейроне, которому этот аксон принадлежит) как об общем конечном пути для управления движением. Любое отдельное мышечное волокно контролируется только одним мотонейроном, но один мотонейрон может контролировать много мышечных волокон с помощью разветвлений своего аксона. Число волокон, управляемых одним мотонейроном, варьирует в зависимости от того, насколько тонкими должны быть движения мышцы. В глазодвигательных мышцах на каждый нейрон приходится примерно по три мышечных волокна; в мышцах, приводящих в движение бедро, на один нейрон приходится сотня мышечных волокон.
Сила, которую может развить мышца, зависит от числа содержащихся в ней мышечных волокон. У мотонейронов, контролирующих крупные мышцы, такие как бицепсы или мышцы голени, аксоны должны иметь много разветвлений, чтобы иннервировать все мышечные волокна, причем веточки аксонов в этом случае гораздо толще, чем у нейронов, управляющих мелкими мышцами пальцев.
Спинной мозг — вместилище мотонейронов
Двигательный нейрон (мотонейрон) и его аксон вместе с мышечными волокнами, которые он контролирует, называют двигательной единицей. Такие единицы можно приближенно сравнить с входными участками сенсорных систем в том отношении, что они ближе всего к внешнему миру. В этом смысле спинной мозг занимает в нейронном «конвейере» примерно такое же положение, как сетчатка в зрительной системе. И спинной мозг, и сетчатка представляют собой скопления нейронов, несколько отодвинутые с периферии вглубь и осуществляющие важные этапы интеграции и фильтрации сигналов с использованием локальных сетей. Относительно простые виды интеграции, возможные на уровне спинного мозга, являются, однако, лишь прелюдией по отношению к более мощным и детализированным двигательным актам, которые могут выполняться под управлением спинного мозга при получении им соответствующих команд из двигательных центров коры больших полушарий.
Спинальные рефлексы. Мышцы снабжены также и чувствительными нервами. Эти нервы участвуют в проприоцепции — позволяют нам чувствовать положение и движения собственного тела. Сенсорные датчики находятся либо в глубине мышц, в специальных комплексах, называемых мышечными веретенами, либо в сухожилиях, там, где мышцы прикрепляются к кости. Эти датчики информируют спинной мозг или двигательные центры более высоких уровней о том, какое напряжение развивает в данный момент мышца. Эта информация помогает определить положение сустава, а тем самым и исходную позицию для начала следующего движения.
Когда доктор во время осмотра проверяет ваши рефлексы, он ударяет молоточком по сухожилию ниже коленной чашки. От этого удара растягивается и расположенное выше сухожилие, прикрепленное непосредственно к четырехглавой мышце бедра. В результате активируются находящиеся в этом сухожилии рецепторы, которые по сенсорным волокнам передают возбуждение спинальным мотонейронам, и последние заставляют мышцу бедра сократиться, а вашу ногу — подпрыгнуть (рис. 57, слева). Весь рефлекс совершается очень быстро, обычно меньше чем за секунду, — так проворно эти нейроны управляются со своими местными делами.
Другие локальные решения, которые также принимаются спинным мозгом, связаны, например, с болевыми раздражителями. Если вас когда-нибудь ударяло током при попытке вытащить из тостера застрявший там кусок хлеба, то вы, возможно, заметили, что ваша рука взлетала в воздух еще до того, как вы почувствовали боль. Под контролем спинного мозга ваша пострадавшая конечность автоматически отдергивается в результате ее сгибания в суставах. При таких неврологических заболеваниях, как рассеянный склероз и амиотрофический боковой склероз, одна из неприятностей состоит в том, что сенсорные нервы не обеспечивают должного выполнения сгибательных рефлексов. В результате больные страдают от частых и длительных соприкосновений с повреждающими объектами.
Реципрокный контроль мышц-антагонистов. Если вы, находясь в сидячем положении, наступите на кнопку, то можете даже не обратить внимания на то, что вы отдернули пораненную ступню, согнув ее. Но вместо этого вы можете заметить, что ваша другая нога отреагировала противоположным движением — разгибанием ступни. Такое движение конечностей называется «перекрестным разгибанием» (рис. 57, справа). Связи между мотонейронами, контролирующими этот рефлекс, устанавливаются в спинном мозге еще до рождения. (Даже младенец, если его держать в вертикальном положении, чтобы он мог свободно двигать ногами, перебирает ими как при ходьбе. Эти движения обусловлены в основном активацией рецепторов кожи и сухожильных рецепторов растяжения.) Сенсорные нервные волокна, расположенные на подошве одной стопы, непосредственно активируют спинальные мотонейроны, которые вызывают сокращение мышц-сгибателей конечности, подвергшейся раздражению. Ветви тех же самых сенсорных волокон возбуждают мотонейроны, контролирующие разгибатели другой ноги.
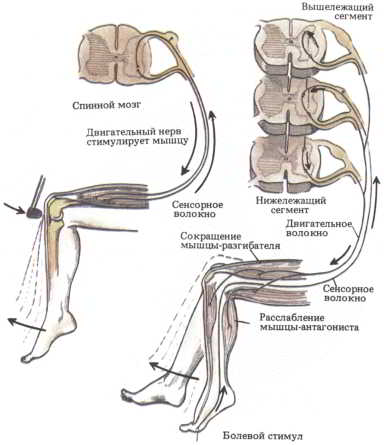
Рис. 57. Когда происходит простое рефлекторное действие, рецепторы растяжения мышцы-разгибателя непосредственно вызывают активацию мотонейронов этой мышцы, что ведет к ее сокращению. При перекрестном рефлекторном действии внутренние связи данного сегмента спинного мозга позволяют периферическим кожным рецепторам и рецепторам растяжения вызывать координированные мышечные сокращения без участия высших уровней двигательной системы. В зависимости от схемы связей команды, поступающие к мотонейронам, управляют мышцами-антагонистами — сгибателями или разгибателями.
Этот реципрокный мышечный контроль и свойственная спинному мозгу перекрестная иннервация позволяют уравновешивать движения наших рук и ног во время ходьбы и почти всех других видов физической активности. Что, например, происходит, когда вы вытягиваете руку вперед и пытаетесь держать ее неподвижно, показывая на какое-нибудь пятно на стене? Мышцам, удерживающим руку в поднятом положении, противодействуют другие, не позволяющие ей подняться слишком высоко. Постоянный контроль за равновесием между группами мышц-антагонистов обеспечивают проприоцептивные нервы, оперативно сообщающие об относительном напряжении и длине соответствующих мышц. Рецепторы, находящиеся внутри сокращенной мышцы, активируются, когда мышца растягивается под действием своего антагониста. Сухожильные рецепторы растяжения возбуждаются по мере развития напряжения в мышце, прикрепленной к кости данным сухожилием. Если у вас устали мышцы плечевого пояса, ваша рука начинает опускаться. Мышечные волокна растягиваются и возбуждают мотонейроны, контролирующие плечевую мускулатуру. В то же время падение мышечного напряжения снижает активность сухожильных рецепторов, и их тормозящее воздействие на мотонейроны ослабевает. В результате усиливается сокращение плечевой мускулатуры и восстанавливается ее воздействие на руку.
С помощью внутренних, локальных систем спинного мозга контроль над всеми этими изменениями осуществляется автоматически, как только выбрана соответствующая программа движения. Однако само решение поднять руку вверх и указывать ею на какое-либо пятно на стене принимается центром более высокого порядка. Первоисточник приказов, получаемых мотонейронами спинного мозга, — это нейроны двигательной коры.
Двигательная кора
Части коры, ведающие движением, были впервые обнаружены при изучении параличей у больных с локальными повреждениями мозга или инсультами. В каждом из больших полушарий имеется полоска коры, связанная с двигательной функцией. Обе эти моторные зоны примыкают к участкам коры с первичными сенсорными проекциями поверхности тела, имеющимися в каждом из полушарий. Прежде считали, что корковая моторная область организована по аналогии с соседней сенсорной областью, т.е. в соответствии с проекцией, отражающей поверхность тела. Эта точка зрения казалась вполне правдоподобной — при стимуляции небольших участков моторной коры в определенных частях тела регистрировались незначительные мышечные сокращения. Но поверхность тела и здесь отображается в искаженном виде: губы, кисти и пальцы занимают гораздо большую площадь, чем ноги и туловище (рис. 58).
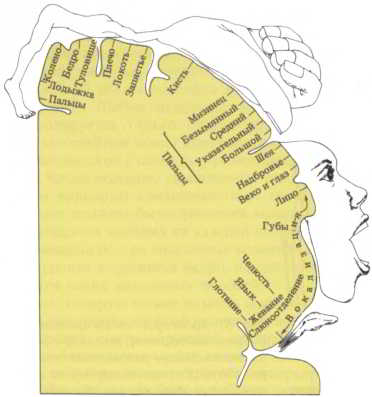
Рис. 58. Двигательная область коры головного мозга человека. На этой карте показаны участки двигательной коры, стимуляция которых приводит к сокращению определенных групп мышц. В частности, отдельные области, по-видимому, могут кодировать угловое положение суставов, приводимых в движение соответствующими мышцами.
Сравнительно недавние работы с регистрацией активности отдельных клеток двигательной зоны с помощью микроэлектродов подсказывают иное объяснение пространственному расположению точек, возбуждающих определенные мышцы. Нейроны в двигательной коре, так же как и в сенсорной зоне, по-видимому, организованы в вертикальные колонки. Как показывают микроэлектродные записи, клетки двигательной коры, связанные по вертикали и образующие функциональную моторную колонку, действительно контролируют группу связанных между собой мышц. В дальнейшем выяснилось, что, как это ни странно, соседние нейроны в двигательной колонке по-разному ведут себя при выполнении движений: одни возбуждаются, другие затормаживаются, с третьими не происходит никаких изменений.
Как сейчас полагают, важнейшая функция корковой двигательной колонки состоит в том, чтобы обеспечивать определенное положение сустава, а не просто активировать те или другие мышцы. В зависимости от исходной позиции сустава данная колонка должна воздействовать либо на мышцы-сгибатели, либо на разгибатели, чтобы придать суставу желаемый угол. С этой точки зрения корковая моторная колонка — это небольшой ансамбль двигательных нейронов, влияющих на все мышцы данного сустава. В несколько более общей форме можно сказать, что кора кодирует наши движения не путем приказов о сокращении отдельных мышц, а путем команд, обеспечивающих определенное положение суставов.
Нейроны коры, непосредственно связанные с мотонейронами спинного мозга, называются клетками Беца по имени впервые описавшего их русского анатома XIX века. Они лежат в глубине двигательной коры и относятся к самым крупным нейронам головного мозга. Их аксоны сходятся в толстый пучок нервных волокон, называемый пирамидным трактом. Дойдя до спинного мозга, аксоны клеток Беца перекрещиваются — пучок, идущий от правого полушария, переходит на левую сторону, и наоборот. Вот почему инсульт или иное повреждение правой моторной зоны вызывает паралич левой половины тела.
Где источник возбуждения, активирующего нейроны двигательной коры? На этот вопрос в настоящее время, по-видимому, можно ответить так: это возбуждение возникает в нейронах сенсорной коры на самом позднем этапе переработки всех форм сенсорной информации. На этом этапе появляются сигналы высокого уровня абстракции, отражающие положение конечностей и необходимость быстрого выполнения тех или иных движений. Эта информация, включающая полные данные о положении суставов и напряжении мышц, и служит той основой, руководствуясь которой моторная кора инициирует определенные движения.
Прежде чем закончить наш обзор двигательной системы, мы должны кратко рассмотреть еще несколько важных структур, которые тоже регулируют выполнение специфических, направленных произвольных движений. Это базальные ганглии и мозжечок.
Базальные ганглии
Термин «базальные ганглии» звучит не совсем понятно, но он попросту означает, что эти ганглии (т.е. скопления нервных клеток — от греч. ganglion — узел и basalis от греч. basis — основание) лежат у основания больших полушарий. Клетки базальных ганглиев возникают на ранних стадиях развития мозга (см. гл. 1). Если еще раз прибегнуть к нашим географическим аналогиям, то комплекс базальных ганглиев можно сравнить с группой штатов, составляющих, например, Средний Запад или Солнечный берег. К базальным ганглиям относятся четыре нервных образования: полосатое тело (стриатум), бледный шар (паллидум), субталамическое ядро и черная субстанция (substantia nigra) (рис. 59). Эти названия отражают либо местоположение структур (субталамическое — расположенное под таламусом), либо их внешний вид (striatum — полосатый; pallidum — бледный; nigra — черная).

Рис. 59. Базальные ганглии — скопления серого вещества, входящие в экстрапирамидную часть двигательной системы. Информационные взаимодействия между базальными ганглиями координируют крупные мышечные движения, запуская и останавливая их.
Полосатое тело получает информацию (включая все виды сенсорной информации и сведения о состоянии активности двигательной системы) почти от всех областей коры большого мозга. Характерные для этой структуры полосы образованы пучками сильно миелинизированных аксонов, соединяющих ее с двигательной и сенсорной корой. Полосатое тело получает также «сырую» сенсорную информацию от таламических ядер еще до того, как она поступает для переработки в кору. Третий источник информации — дивергентные связи от черной субстанции. Это последнее образование оказывает воздействие на двигательную систему с помощью медиатора дофамина, который в настоящее время хорошо изучен; поэтому оно заслуживает особого внимания.
У лиц, страдающих болезнью Паркинсона, дофаминэргические (т.е. выделяющие в качестве медиатора дофамин) нейроны черной субстанции отмирают. В начале нашего века при вскрытиях было обнаружено, что у больных паркинсонизмом в этой области отсутствует черный пигмент. Когда медиатор отмирающих нейронов был идентифицирован как дофамин, стало возможным объяснить потерю цвета гибелью самих нейронов и утратой дофамина. Затем была установлена прямая связь между этими изменениями и характерными начальными симптомами заболевания — неспособностью приступить к выполнению произвольных движений, сочетающуюся с дрожанием головы и рук, когда больной сидит спокойно. Хотя в полосатом теле больше дофаминэргических синапсов (на единицу объема), чем в любой другой области мозга, он выделяется здесь, вероятно, меньше чем в одной пятой части всех синаптических связей. И тем не менее утрата дофаминэргических волокон и осуществляемого ими контроля катастрофически нарушает нормальную работу двигательной системы. Больных, однако, можно в течение некоторого времени успешно лечить препаратом L-ДОФА (дигидроксифенилаланином), который поддерживает тающие при паркинсонизме запасы дофамина.
Регистрация активности нейронов полосатого тела показала, что их разряды начинаются непосредственно перед началом движений определенного типа — медленных целенаправленных перемещений конечности из одной области пространства в другую. Когда вы, закрыв глаза, пытаетесь рукой коснуться кончика носа, большая часть этого движения — перемещение руки из исходного положения почти до самого носа — осуществляется под контролем базальных ганглиев. Именно этот вид движений и нарушается у больных паркинсонизмом.
У животных с разрушенными в эксперименте дофаминэргическими нейронами, аксоны которых оканчиваются в полосатом теле, наступает критический период, во время которого они как будто утрачивают способность начинать двигательные акты, даже такие важные, как еда и питье. Если им предлагают пищу с сильным запахом, повышение сенсорной активации отчасти помогает им преодолеть это затруднение. Люди, страдающие болезнью Паркинсона, тоже могут временно преодолевать дефекты двигательной системы — в экстренных ситуациях, если, например, видят приближающийся автомобиль в тот момент, когда как раз собираются сойти с тротуара. У таких больных редко возникают трудности с речью или движениями глаз; это позволяет думать, что подобные функции осуществляются без участия дофамина.
Больные паркинсонизмом должны принимать L-ДОФА в течение всей оставшейся жизни. Однако эксперименты по пересадке дофаминэргических нейронов, проводимые сейчас в Швеции, вселяют надежду на возможность более радикального лечения этого недуга в будущем.
Мозжечок
Как видно из самого названия (мозжечок — уменьшительная форма от слова «мозг»), это действительно малый мозг. Он имеет необычайно регулярную структуру, и поверхность его во много раз увеличивают многочисленные складки (рис. 60). На разрезе его отдельные складчатые дольки по виду будут напоминать листья. Они так и называются листки мозжечка. В каждом из них, изгибаясь вместе с наружной поверхностью, повторяется одна и та же слоистая структура. Один из двух наиболее заметных слоев содержит очень крупные нейроны — клетки Пуркинье, образующие отдельный слой. В другом слое находятся клетки-зёрна, которые образуют скопления толщиной в несколько клеток непосредственно под нейронами Пуркинье.

Рис. 60. Срединная поверхность мозжечка (вид слева). Показаны листовидные образования, каждое из которых содержит сильно изрезанную слоистую структуру. В ней имеются зоны, богатые волокнами или же клетками. Каждый лист контролирует активность в пределах определенных групп мышц.
Информация поступает в мозжечок из коры больших полушарий, ствола мозга и спинного мозга. Спинной мозг передает сведения о положении конечностей, туловища, головы, шеи и глаз. Всю эту информацию интегрируют клетки Пуркинье. Эти нейроны дают разряды очень частых импульсов и большую часть времени находятся в активном состоянии, что, по-видимому, свидетельствует об их постоянном контроле над положением туловища, конечностей и головы. Нейроны Пуркинье в свою очередь посылают информацию крупным нейронам глубинных ядер мозжечка, соответствующих по своему рангу отдельным деревням в нашей географической схеме. Информация, которую выдают эти ядра, видоизменяет активность нейронов моторной коры.
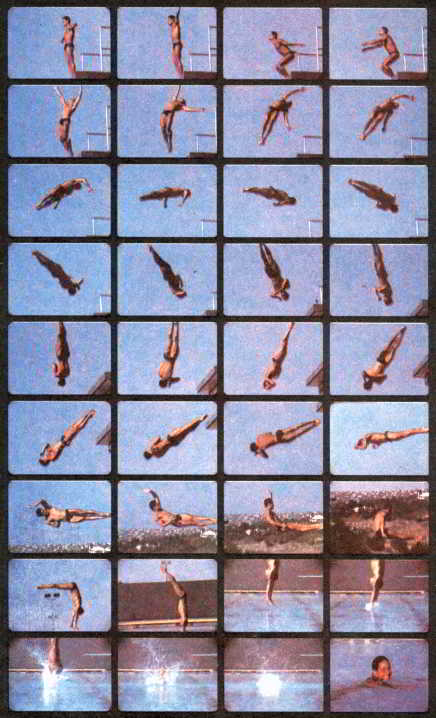
Рис. 61. Ныряние — один из естественных способов перехода с суши в воду. Но выполнение прыжка с 30-метровой вышки с двумя одновременными вращательными движениями есть нечто выходящее за рамки всякой целесообразности. Такой прыжок — это изысканная, исключительно сложная серия движений, исполняемых ради самого движения, ради его трудности и красоты. В борьбе за совершенство управление телом у Грега Луганиса должно было достичь такой степени точности и согласованности, которые находятся на крайнем пределе нейробиологических возможностей человека.
Несмотря на изящную регулярную структуру мозжечка и хорошо изученные нейронные сети, конкретная роль его в двигательной функции еще далеко не ясна. Весьма грубые наблюдения, сделанные при повреждениях или раздражении мозжечка у людей, указывают на важную роль этого органа в регуляции мышечного тонуса, необходимого для поддержания позы. То, как мозг справляется с этой задачей, показывают тесты, предлагаемые людям для оценки степени опьянения, требуется пройти по прямой линии или стоять прямо с закрытыми глазами.
Кроме того, при осуществлении более тонких движений мозжечок определяет, где находятся в каждый данный момент части тела, и сравнивает их действительное положение с тем, которое должно быть. Похоже, что мозжечок располагает копией той программы движения, которой руководствуются нейроны двигательной коры. Мозжечок согласовывает активность моторной коры и спинного мозга, обеспечивая более «гладкое» выполнение контролируемых ими тонких движений. Когда вы пытаетесь дотронуться пальцем до кончика носа, общим движением руки по направлению к носу руководят базальные ганглии, но именно мозжечок управляет заключительным прикосновением.
Мозжечок играет также важную роль при выполнении быстрых последовательных и одновременных движений, таких, например, как уверенные движения рук опытной машинистки или музыканта. Справляется он и с более примитивными задачами вроде одновременного похлопывания по голове и почесывания груди.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
«Движение сопротивления»
«Движение сопротивления» Достижения генетиков радуют далеко не всех. В мире уже появились профессиональные борцы с «индустриальной пищей». Они уничтожают посевы с генетически измененными культурами, отказываются от консервов из трансгенных томатов и других
Движение — простейшая форма поведения
Движение — простейшая форма поведения Тропизмы Первое наиболее четкое различие между животными и растениями ясно каждому: растения не могут передвигаться, тогда как животные этим свойством обладают. И тем не менее именно движение растений (поворот к солнцу цветов)
Нейтральные мутации и генетический дрейф — движение без правил
Нейтральные мутации и генетический дрейф — движение без правил Ландшафт приспособленности — образ яркий и полезный, но, как и всякая модель, он несовершенен. Многие аспекты эволюционного процесса с его помощью отразить трудно или невозможно. Реальный ландшафт
Глава 6. Движение
Глава 6. Движение Природы зуб и коготь ал” — это, должно быть, одна из самых цитируемых, к месту и не к месту, аллюзий на Дарвина в англоязычной литературе. Но как бы не к месту ее иногда ни цитировали, эта фраза верно передает если не истинную суть естественного отбора, то,
9.3. Амебоидное движение
9.3. Амебоидное движение Амебы движутся с помощью объемного тока своей цитоплазмы в вырастающие выступы — псевдоподии. В норме они перемещаются по поверхности твердых объектов посредством непрерывного растягивания своих передних концов. Но если до псевдоподий
Движение живых объектов
Движение живых объектов Глядя на то, как двигается тот или иной объект, уже можно сказать, живое ли это существо или просто лист, летящий по ветру. А можно сказать и намного больше. Можно, например, увидеть, что это человек, и увидеть, что он делает. Информации для этого
Движение
Движение О том, что гормоны участвуют в обеспечении движения, известно со времен введения понятия «гормон». Удаление половых желез у самцов приводит к постепенному, но постоянному снижению двигательной активности. В дальнейшем было обнаружено, что она также падает после
3. Ощущение и движение
3. Ощущение и движение Жара повсюду. Ее нельзя больше не замечать. Воздух раскален, как в доменной печи, — он настолько горячий, что мои глаза, защищенные очками, кажутся прохладными по сравнению с остальным лицом. Руки тоже прохладные, но на тыльной стороне перчаток
Движение
Движение «Объединители» и «дробители» этого мира пытаются проникнуть в суть явлений разными путями. Приверженцы классического подхода — объединители — хотят найти то, что объединяет вещи. Они ищут законы, управляющие функциями целого. Мыслители романтического склада
Внутреннее ухо — движение желе и колебание волосков
Внутреннее ухо — движение желе и колебание волосков Представьте себе, что мы заходим в слуховой проход, проходим сквозь барабанную перепонку, мимо трех косточек среднего уха и оказываемся глубоко внутри черепа. Здесь расположено внутреннее ухо — заполненные