Эндокринная система
Эндокринная система
Эндокринный орган отличается тем, что выделяет вещество, необходимое для регуляции клеточной активности каких-то других органов, непосредственно в кровяное русло (термин происходит от греч. endo — внутри и krinein — выделять). Такие органы называются эндокринными железами, а секретируемые ими вещества — гормонами (от греч. hormao — возбуждаю). Каждый гормон влияет на уровень функционирования специфических систем клеток-мишеней — обычно временно повышает их активность. Гормоны — сильнодействующие агенты, поэтому для получения специфического эффекта достаточны ничтожные их количества. Восприимчивые к гормонам клетки снабжены специальными поверхностными молекулами — «рецепторами», которые реагируют даже на очень низкие концентрации гормонов. После соприкосновения рецептора с гормоном внутри клетки происходит ряд изменений.
Эндокринные органы и их гормоны
По традиции эндокринную и нервную системы с их регулирующими и интегрирующими функциями считали отдельными, но параллельно действующими системами. Нейроны выделяют свои химические передатчики — медиаторы — в синаптическую щель для регуляции активности других нейронов. Эндокринные клетки секретируют свои химические передатчики — гормоны — в кровь, которая разносит их ко всем клеткам, имеющим специфические рецепторы (рис. 67). Некоторые вещества действуют в обеих системах; они могут быть и гормонами (т.е. продуктами эндокринных желез), и медиаторами (продуктами определенных нейронов). Такую двоякую роль выполняют норадреналин, соматостатин, вазопрессин и окситоцин, а также передатчики диффузной нервной системы кишечника, например холецистокинин и вазоактивный кишечный полипептид.
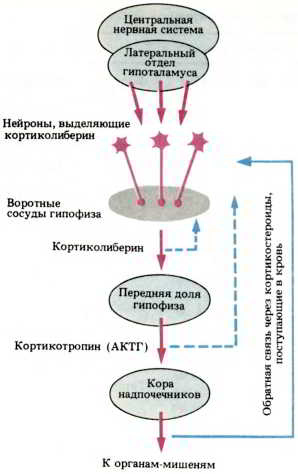
Рис. 67. Связь между центральной нервной системой и эндокринной системой. Нейроны гипоталамуса вырабатывают кортиколиберин, который попадает в переднюю долю гипофиза через систему воротного кровообращения. Нейроны гипофиза в ответ на это выделяют кортикотропин (АКТГ), стимулирующий секрецию кортикостероидов корой надпочечников. Уровень кортикостероидов в крови, действуя как сигнал обратной связи, заставляет гипофиз или центральную нервную систему в целом продолжать или приостанавливать этот процесс.
Железы, входящие в состав эндокринной системы, — это гипофиз с его независимо функционирующими передней и задней долями, половые железы, щитовидная и паращитовидные железы, кора и мозговой слой надпочечников, островковые клетки поджелудочной железы и секреторные клетки, выстилающие кишечный тракт. Важнейшие сведения об эндокринных железах приведены в табл. 4.1.
Таблица 4.1. Эндокринная система
Орган или ткань Гормон Клетки-мишени Вызываемые эффекты Гипофиз, передняя доля Фолликулостимулирующий гормон Половые железы Овуляция, сперматогенез Лютеинизирующий гормон - // - Созревание яйцеклеток и сперматозоидов Тиреотропный гормон Щитовидная железа Секреция тироксина Адренокортикотропный гормон Кора надпочечников Секреция кортикостероидов Гормон роста (соматотропин) Печень Секреция соматомедина Все клетки Синтез белков Пролактин Молочные железы Рост желез и секреция молока Гипофиз, задняя доля Вазопрессин Почечные канальцы Задержка воды в организме Артериолы Повышение кровяного давления Окситоцин Матка Сокращение Половые железы Эстроген Многие органы Развитие вторичных половых признаков Тестостерон - // - Влияние на рост мышц, молочных желез Щитовидная железа Тироксин - // - Повышение интенсивности обмена веществ Паращитовидные железы Кальцитонин Кость Задержка кальция Кора надпочечников Кортикостероиды Многие органы Мобилизация энергетических ресурсов; сенсибилизация адренэргических рецепторов в сосудах; торможение образования антител и воспалительных процессов Альдостерон Почки Задержка натрия Мозговое вещество надпочечников Адреналин Сердечно-сосудистая система, кожа, мышцы, печень и другие органы Симпатическая активация Островки поджелудочной железы Инсулин Многие органы Усиленное поглощение глюкозы клетками Глюкагон Печень, мышцы Повышение уровня глюкозы в крови Соматостатин Островки поджелудочной железы Регуляция секреции инсулина и глюкагона Слизистая кишечника Секретин Экзокринные клетки поджелудочной железы Секреция пищеварительных ферментов Холецистокинин Желчный пузырь Выведение желчи Вазоактивный кишечный полипомид Двенадцатиперстная кишка Усиление моторики и секреции; увеличение кровотока Тормозящий пептид - // - Торможение моторики и секреции Соматостатин - // - То жеПо традиции гипофизу приписывалась также роль «главной железы» эндокринной системы. Однако новые данные о том, что клетки передней доли гипофиза сами находятся под контролем гипоталамических нейронов, заставили пересмотреть этот взгляд. Передняя доля содержит несколько различных типов эндокринных клеток, каждый из которых вырабатывает один из гипофизарных гормонов и регулируется специфическими гормонами гипоталамуса. Гипоталамус связан с гипофизом небольшой локальной сетью кровеносных сосудов, так называемой воротной системой гипофиза, которая доставляет кровь от основания гипоталамуса к передней доле гипофиза. Гипоталамические нейроны выделяют в кровь этой сети свои гормоны, а соответствующие клетки гипофиза реагируют на эти гормоны после их связывания специфическими поверхностными рецепторами.
До сих пор идентифицированы шесть гипоталамических гормонов, избирательно воздействующих на клетки передней доли гипофиза. Каждый из этих гормонов — продукт специфической группы нервных клеток, расположенных в перивентрикулярной или средней зоне гипоталамуса (рис. 9 и 68). Четыре гормона стимулируют синтез и секрецию гормонов клетками-мишенями, а два — тормозят.

Рис. 68. Специфические группы клеток передней доли гипофиза с помощью гормонов управляют определенными эндокринными органами, расположенными в разных областях тела. Каждая из этих групп гипофизарных клеток находится под контролем стимулирующих или тормозящих факторов, выделяемых нейронами гипоталамуса в систему воротного кровообращения гипофиза. АКТГ — адренокортикотропный гормон (кортикотропин); КЛ — кортиколиберин; ЛГ — лютенизирующий гормон; ЛЛ — люлиберин; СЛ — соматолиберин; ТЛ — тиреолиберин; ТТГ — тиреотропный гормон; ФЛ — фоллиберин; ФСГ — фолликулостимулирующий гормон.
Поскольку эти специфические нейроны оказывают на гипофиз мощное влияние, истинной «главной железой» эндокринной системы следует считать головной мозг и, в частности, гипоталамус. Первым звеном в гипоталамическом контроле над эндокринной системой служит передача гормональных посредников через воротную систему гипофиза. Те же гипоталамические нейроны могут образовывать в мозгу и другие связи — синаптические. В этом случае их секреторные продукты выступают в роли нейромедиаторов. Например, соматостатиновые нейроны перивентрикулярной зоны и некоторые клетки коры больших полушарий и гиппокампа используют одни и те же медиаторы, а соматостатин, образующийся в островках поджелудочной железы, действует как «локальный гормон», регулируя секрецию инсулина и глюкагона.
По мнению ряда ученых, медиаторы, выделяемые нейронами, могут также действовать как локальные гормоны в вегетативной и центральной нервной системе. Если это так, то нейроны центральной нервной системы, использующие медиаторы подобным образом, будут в чем-то аналогичны нейронам локальных сетей, регулирующим поток информации внутри отдельных областей ЦНС.
Таким образом, весь процесс, с помощью которого мозг соотносит нужды организма с требованиями окружающей среды, вернее было бы рассматривать как одну из нейроэндокринных функций. Некоторые адаптации осуществляются в локальных участках вегетативной нервной системы и координируются специфическими локальными гормонами, а другие реализуются в более глобальных масштабах с помощью веществ-посредников, выделяемых в кровяное русло.
Эндокринология и гомеостаз
Некоторые фундаментальные концепции традиционной эндокринологии тесно связаны с представлением о гомеостазе. Секреция некоторых гормонов, например тироксина, регулируется очень жестко. Однако концентрации большинства других гормонов могут в широких пределах изменяться для поддержания постоянства ряда физиологических параметров при непрерывном изменении сиюминутных потребностей организма. Например, скорости секреции инсулина и глюкагона сильно колеблются, чтобы удерживать концентрацию глюкозы в крови в допустимых пределах. Изменения уровней альдостерона (см. выше табл. 4.1) и вазопрессина отражают необходимость сохранять постоянный объем крови путем регуляции водно-солевого баланса. Концентрации адреналина и норадреналина зависят от степени общей активности организма и могут быть разными в различных локальных сосудистых сетях. Это позволяет им регулировать силу и частоту сердечных сокращений, а также избирательно воздействовать на сосуды, чтобы обеспечивать приток крови к определенным системам органов в соответствии с потребностями.
Однако оставим в стороне изменчивость гормональных уровней и рассмотрим нейроэндокринные воздействия как способ поддержания «заданных значений», определяющих постоянство системы.
Физиологические «заданные значения»
Температура тела, уровень глюкозы в крови, кровяное давление, концентрация солей — таковы лишь некоторые из тщательно регулируемых физиологических параметров здорового организма. Концепция «заданных значений» помогает понять, каким образом ЦНС, вегетативная нервная система и нейроэндокринные компоненты совместно участвуют в регуляции этих и иных параметров.
Представим себе, что организм функционирует таким, например, образом, чтобы сохранять постоянные значения температуры тела и концентраций глюкозы, солей и кислорода в крови. Если соответствующие датчики улавливают какое-нибудь отклонение, они приводят в действие адаптивные механизмы, с тем чтобы вернуть величину к заданному уровню. Такие системы работают по принципу отрицательной обратной связи между периферийными датчиками и центральным управляющим звеном. На примере трех физиологических систем — регуляции температуры тела, давления и аппетита — мы проиллюстрируем основные особенности такого рода обратной связи.
Терморегуляция. За температурой тела следят наружные терморецепторы кожи и внутренние терморецепторы в перивентрикулярной зоне гипоталамуса. Внутренние рецепторы, измеряющие истинную температуру крови, по-видимому, наиболее существенны для автоматической регуляции температуры тела. Эксперименты с введением небольших зондов прямо в гипоталамус подопытных животных показали, что нейроны, расположенные около переднего края перивентрикулярной зоны гипоталамуса, могут активироваться в результате подъема или падения температуры артериальной крови. Одновременно по приказу гипоталамуса вегетативная нервная система приводит в действие механизмы сохранения, выработки или отдачи тепла.
Если гипоталамус ощутит снижение температуры тела, периферические вегетативные ганглии осуществят переброску крови от кожи к более глубоким структурам; под их воздействием шерсть или перья встанут дыбом, чтобы увеличить слой прилегающего к коже теплого воздуха. Так называемая гусиная кожа, которая появляется у вас на теле, если вы замерзли, — остаток этого механизма. Для согревания тела служит также дрожь, при которой образуется дополнительное тепло. Если гипоталамические датчики улавливают подъем температуры артериальной крови, включаются механизмы, повышающие теплоотдачу: кровь приливает к коже и тепло быстрее переходит во внешнюю среду; может начаться потоотделение, еще более эффективно охлаждающее тело.
Даже при «эмоциональном перегреве» у вас возможно потоотделение (физически ненужное!), которое должно будто бы охладить ваш пыл. Во время сильной тревоги у вас «леденеют» руки и ноги — вам хочется как бы сжаться, спрятаться внутрь себя от вызывающих беспокойство мыслей. Сегодня, однако, умозрительные рассуждения о психологическом «значении» физиологических реакций при отклонениях от комфортного эмоционального состояния так и остаются рассуждениями. Повышение тонуса симпатической нервной системы в напряженных ситуациях — вот вполне достаточное объяснение тому, почему у вас холодеют руки и ноги, сильнее бьется сердце, сохнет во рту, а глаза, не мигая, смотрят в одну точку.
Когда температура тела выходит за рамки нормальных колебаний — например, при инфекционных заболеваниях — это означает, что механизмы выработки и сохранения тепла действуют чересчур активно. Белые кровяные тельца (лейкоциты) у заболевшего выделяют пока еще мало изученное вещество — лейкоцитарный пироген, который активизирует эти механизмы удержания тепла, тем самым повышая температуру тела. Быстрый подъем температуры у больного часто парадоксальным образом сочетается с ощущением холода. Так возникает всем известный озноб, сопровождающий наступление лихорадочного состояния.
«Цель» лихорадки неясна. Некоторые, но далеко не все болезнетворные бактерии действительно плохо переносят высокую температуру, и в этом случае жар может облегчать организму борьбу с инфекцией. В качестве одного из примеров назовем бледную спирохету, возбудитель сифилиса. (Вирусы, вызывающие катар верхних дыхательных путей, не погибают, так как вдыхаемый через нос воздух снижает местную температуру до 37°С.) Повышенная температура тела способствует также усилению активности некоторых клеток, вырабатывающих антитела, и может ускорять продвижение белых кровяных телец к инфицированной области, где они участвуют в уничтожении болезнетворных агентов.
Повышенная способность приводить в действие механизмы сохранения или отдачи тепла — один из аспектов процесса адаптации. Вы, вероятно, замечали, что зимой легче переносить холод, а летом — жару. Ваш организм в буквальном смысле предугадывает требования окружающей среды. Если человек часто занимается физкультурой, он намного легче потеет, так как его мозг учится все быстрее и быстрее приводить в действие координированные программы, связанные с преодолением физических нагрузок.
Регуляция давления и объема крови. Каждый уровень центральной и периферической нервной системы в какой-то мере участвует в поддержании постоянства кровообращения. Рецепторы давления, или барорецепторы, регистрируют фактическое давление крови в крупных артериях, расположенных выше сердца, — сонных артериях и в дуге главной артерии — аорты. При избыточном давлении происходит активация рецепторных нейронов в стенках этих артерий. Отростки этих нейронов передают соответствующую информацию первичному передаточному ядру в продолговатом мозге, откуда идут тормозные сигналы к вазомоторным центрам, а также к периферической симпатической нервной системе, в результате чего снижается ее активность. Одновременно усиливаются парасимпатические влияния на сердечно-сосудистую систему.
Нормальное кровяное давление — это результат непрерывной борьбы за контроль над сердечно-сосудистой системой. Симпатические нервы контролируют крупные кровеносные сосуды, а частота и сила сердечных сокращений регулируется как симпатическим, так и парасимпатическим отделами. Чувствительные к давлению барорецепторы, по-видимому, ответственны в первую очередь за установление нормальной точки отсчета, около которой происходят колебания при координации деятельности сердечнососудистой системы. Барорецепторы наиболее чувствительны к изменениям, выходящим за пределы нормальных средних величин давления крови (которые соответствуют давлению столбика ртути высотой 100-120 мм).
Помимо барорецепторов аорты и сонных артерий в верхних отделах сердца — предсердиях — имеются другие рецепторы растяжения, чувствительные к давлению крови. Поскольку давление крови, поступающей в правое предсердие из венозной системы, более точно отражает объем крови, чем среднее давление, рецепторы предсердий действуют на иной информационной основе. Они активируются в случае чрезмерного увеличения объема крови, как это бывает при задержке соли в организме или при ранних явлениях застойной сердечной недостаточности. Это приводит к тому, что клетки, расположенные в стенках самого предсердия, выделяют гормон, действующий на почечные канальцы и ускоряющий выведение солей.
На более высоких уровнях управления относительную концентрацию солей в плазме регулируют нейроны гипоталамуса: они приводят в действие механизмы задержки воды или ее ускоренного выведения. Если концентрация солей в плазме превышает определенный уровень, крупные нейроны медиальной зоны гипоталамуса выделяют вазопрессин, который переходит из их аксонов прямо в венозную кровь задней доли гипофиза. Вазопрессин с кровью достигает клеток дистальных почечных канальцев и заметно усиливает обратное всасывание ими воды. Это приводит к задержке жидкости и снижению концентрации солей.
В регуляции объема крови участвует еще одно взаимодействие между мозгом и почками. В почках есть специальные клетки, называемые юкстагломерулярными. Они активируются при падении кровяного давления, например вследствие сильной кровопотери, особенно при повышении активности симпатической нервной системы. Эти почечные клетки выделяют в кровь фермент ренин, который воздействует на синтезируемый печенью низкомолекулярный белок ангиотензиноген. Процесс его расщепления продолжается при участии других ферментов в легких и мозге и в конце концов приводит к образованию белкового фрагмента ангиотензина II.
Ангиотензин II вызывает сокращение мускулатуры артерий и тем самым — быстрое и сильное повышение кровяного давления. Он воздействует также на некоторые рецепторные клетки гипоталамуса. Через еще не вполне изученные клеточные механизмы ангиотензин стимулирует усиленное потребление воды, что, конечно, ведет к увеличению количества жидкости в организме. Действительно, после прямой инъекции ничтожно малых доз (одной миллионной доли микрограмма) ангиотензина II в желудочки мозга животное, незадолго до того уже удовлетворившее жажду, снова начинает жадно пить, чем бы оно в тот момент ни занималось. Потребность в питье часто наблюдается у раненых солдат и других людей, потерявших много крови.
УДАР ПО ГОЛОВЕ
«Я мужчина, и я хотел вести ту жизнь, которую, по моему мнению, должен вести нормальный мужчина...»
Митч Хеллер нормально рос — играл в хоккей, закончил колледж, получил специальность инженера, женился. Однако после автомобильной аварии, при которой он получил сильный ушиб головы, все изменилось.
«Примерно через месяц я заметил, что секс стал меньше интересовать меня. Я был не на высоте... Я знал, что во мне что-то происходит, но не знал, что именно.»
Нарушилось не только половое влечение. У Митча начались изменения вторичных половых признаков — например, исчезли волосы на лице и груди, число сперматозоидов в семенной жидкости уменьшилось и продолжало снижаться. Все эти признаки свидетельствовали о падении уровня тестостерона, но как это могло быть связано с травмой головы?
Д-р Уильям Кроули из Массачусетской больницы имел на этот счет некоторые соображения. Очевидно, был поврежден участок мозга, противоположный месту внешнего удара (явление «противоудара»), где находились нейроны гипоталамуса, секретирующие гонадолиберин (фактор, стимулирующий секрецию гонадотропных гормонов гипофизом). Это привело к тому, что семенники перестали получать от гипофиза стимул к выработке тестостерона. Кроули предложил больному носить механический насос, через который осуществлялась автоматическая инъекция гонадолиберина. Каждые два часа через иглу, введенную подкожно в области живота, гормон поступал в организм.
Применение насоса быстро привело к желаемым результатам. Потенция и рост волос на теле постепенно восстановились. Через шесть месяцев почти нормализовалось количество сперматозоидов, и жена Митча забеременела. Возможно, что нормальные функции гипоталамуса со временем восстановятся, но если даже этого не произойдет, худшие опасения Митча позади.

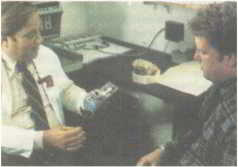

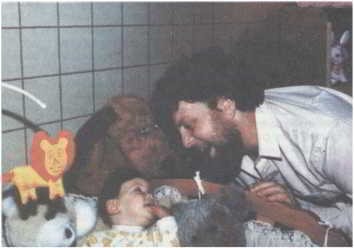
Рис. 69. Вверху: Митч с женой до лечения. Ниже: д-р Кроули объясняет, как действует насос. Внизу: Митч с дочерью.
Информация поступает и в сердечно-сосудистые центры нижней части продолговатого мозга от сосудистых рецепторов давления, а также от хеморецепторов, которые реагируют на концентрации кислорода и СО2 в циркулирующей крови. Эти центры усиливают импульсную активность нейронов в системах, управляющих дыхательными и сердечнососудистыми функциями. Активация центров, расположенных в боковых частях продолговатого мозга, приводит к возбуждению периферической симпатической системы и повышению кровяного давления, а центров средней части — к возбуждению парасимпатических отделов и снижению кровяного давления. Первичные передаточные нейроны продолговатого мозга передают информацию о давлении и токе крови центрам более высокого уровня, находящимся в гипоталамусе и ретикулярной активирующей системе (см. ниже), которые связаны с корой больших полушарий и мозжечка. Эти структуры в свою очередь принимают существенное участие в регуляции кровоснабжения мышц в соответствии с текущими потребностями.
У лиц с хронически повышенным давлением крови, по-видимому, слишком высоки «заданные значения» этого показателя. Первопричину такой аномалии в большинстве случаев очень трудно установить. Здесь могут играть роль все уровни контроля над сосудистой системой. В одних случаях к повышению давления приводят заболевания почек, в других — чрезмерная активность симпатической нервной системы. В зависимости от вероятной причины врач выбирает наиболее подходящий путь лечения болезни: хирургическое вмешательство, мочегонные для воздействия на почки или другие лекарства.
Регуляция аппетита. Условия, при которых возникает чувство голода, — это до некоторой степени индивидуальная особенность. Есть или не есть, что есть и когда — решение этих вопросов в сильной степени зависит от высших интегративных функций коры больших полушарий. На наш выбор оказывают влияние и социальные моменты. При обилии и разнообразии доступных пищевых продуктов люди, по крайней мере в большинстве развитых стран, решают не только когда следует начинать есть, но и что именно есть, как долго и когда нужно остановиться. Эти часто неосознаваемые решения, связанные с принятием пищи, нередко бывают причиной таких нарушений аппетита, как его полная потеря (анорексия) или, наоборот, «волчий аппетит» (гипербулимия). Подобные расстройства не наблюдаются у лабораторных или диких животных, хотя чрезмерное потребление пищи можно иногда вызвать в эксперименте с помощью слабого хронического болевого раздражения (источником его может быть, например, скрепка на хвосте у крысы) или наблюдать у крыс и мышей с наследственной формой ожирения.
Структурные компоненты системы, регулирующей аппетит, выделить гораздо труднее, чем элементы систем терморегуляции или поддержания нормального кровяного давления. Пока неизвестно даже, от какого именно фактора (или факторов) зависит установление «заданного уровня» аппетита.
Рецепторы растяжения в стенках желудка обеспечивают один из видов информации о пустом желудке. Наряду с урчащими звуками и осознанием того, что подошло время принимать пищу, импульсы от механорецепторов, возникающие при пустом желудке, служат отчетливым сигналом к началу еды. Однако животные и после перерезки сенсорных нервов желудка едят нормально и не страдают потерей или избытком веса, несмотря на отсутствие у них обратной связи от периферии.
При снижении концентрации глюкозы в крови изменяется активность некоторых нейронов вентрикулярной зоны гипоталамуса. Таким образом, эти нейроны, по-видимому, чувствительны к уровню глюкозы. Другие гипоталамические нейроны чувствительны к содержанию в крови жирных кислот, а возможно, и инсулина. Действуя совместно с рецепторами глюкозы, эти нейроны могут активировать «системы аппетита» в латеральной зоне гипоталамуса и побуждать к приему пищи. При электрической или химической стимуляции вентромедиального ядра боковой области гипоталамуса животные едят чаще и дольше, чем контрольные, а животные с обширными повреждениями этой области мозга могут вообще прекратить потребление пищи. Однако очень небольшие повреждения в непосредственной близости от вентромедиального ядра вызывают, наоборот, постоянное переедание у крыс и кошек, что приводит к чрезвычайно сильному ожирению.
До сих пор не было обнаружено гормонов или медиаторов, которые возбуждали бы или подавляли аппетит. Исключение, возможно, составляет холецистокинин — локальный гормон, выделяемый слизистой кишечника на ранних стадиях пищеварительного процесса: он действует как мощный фактор, сдерживающий аппетит. Этот гормон, по-видимому, активирует сенсорные волокна тех ветвей блуждающего нерва, которые передают висцеральную информацию обратно в продолговатый мозг. При перерезке этих ветвей сигналы обратной связи о процессе пищеварения утрачиваются. Холецистокинин выполняет также важные медиаторные функции во многих участках головного мозга, но связан ли какой-либо из этих участков с регуляцией потребления пищи, пока неизвестно.
У большинства организмов потребление пищи хорошо сбалансировано с расходом энергии, но как обеспечивается этот баланс, еще не ясно.
Другие системы «заданных значений». Другие внутренние регуляторные системы не менее сложны. В репродуктивной системе, например, существует целый ряд петель эндокринной обратной связи, которые наряду с прямым воздействием центральной и периферической нервной системы позволяют индивидууму подготовиться, выбрать партнера и осуществить все необходимое для продолжения рода.
Другие сложные системы контроля обеспечивают повышение секреции кортикостероидов во время стрессов или быстрое ее подавление, когда опасность миновала (см. гл. 6). Поскольку на многие внутриклеточные процессы влияют ионы кальция, находящиеся вне клетки, их концентрации тоже тщательно регулируются. В этой регуляции (в которой нервная система играет лишь незначительную роль), участвуют клетки паращитовидных желез, секретирующих кальцитонин и паратгормон, а также клетки почек, печени, костей и даже кожи, синтезирующие витамин D, необходимый для всасывания содержащегося в пище кальция.
Регуляция общего уровня активности
До сих пор мы рассматривали внутренние процессы организма, которые регулируются мозгом. Координируя их, мозг осуществляет общий надзор за тем, что требуется для выполнения текущей деятельности, предугадывает, что понадобится в ближайшем будущем, и приспосабливает имеющиеся ресурсы к возникающим нуждам. Обычно мозг выполняет эти обязанности, не затрагивая деятельности, происходящей на уровне сознания, — за исключением, может быть, тех случаев, когда нужно сделать выбор. Ведет ли эта тропа к водопою? Вкусная ли это еда или меня от нее тошнило? Можно ли этим путем наверняка попасть в деревню?
Если, вообще говоря, мы не осознаем того, что происходит у нас внутри, то на что же в таком случае тратится наше «мыслительное» время? Происходит ли в нашем мозгу такая же интеграция данных, когда мы осматриваем окружающий мир и определяем для себя — иногда с помощью слов, иногда без них, — что мы должны сделать, каким именно способом и что препятствует выполнению задуманного?
Мы действительно должны соединить наши знания о внешнем мире с теми двигательными программами, которыми мы пользуемся, чтобы поддерживать контакты с этим миром. Мы ощущаем мир с помощью специализированных сенсорных систем. Поступающая информация идет по параллельным каналам к вертикальным клеточным ансамблям коры головного мозга, а затем — по последовательным иерархическим путям внутри мозга. Здесь снова происходит ее объединение. В результате создается общая сенсорная «картина», которую мы используем для того, чтобы подогнать наши двигательные программы к текущей ситуации и сравнить наши возможности действия с аналогичными ситуациями из прошлого опыта. Мир, в котором мы живем, беспрестанно меняется, и наши чувства должны четко улавливать эти изменения, если мы хотим их верно анализировать. Теперь мы должны рассмотреть вопрос: как нам удается сохранять необходимую остроту наблюдений?
Если вам случится быть в том месте, где работают другие люди, улучите момент и понаблюдайте за ними. Вы довольно быстро сумеете сделать кое-какие выводы. Например, без труда отличите тех, кто дремлет, по то и дело смежающимся векам и опускающейся голове: активность парасимпатической системы после еды подавляет действие норадреналина, который стимулировал бы выполнение рабочих обязанностей.
Оценить другие моменты может быть труднее. Представьте, что вы сидите в классе и внимательно слушаете вашего преподавателя. Можете ли вы одновременно уделять внимание передвижению других людей в классной комнате, или тому, что происходит в коридоре, или мыслям о том, что вы оставили машину в неположенном месте, чтобы успеть на урок? И какова степень внимания, с которым вы слушаете учителя? Может быть, вы предаетесь своим мыслям и возвращаетесь к уроку лишь тогда, когда учитель делает неожиданные быстрые движения? Можно ли, глядя на вас, сказать, внимательны вы или нет, и что это вообще такое — «быть внимательным»?
Сон и бодрствование
Специалисты, изучающие мозг, пользуются понятием «поведенческий статус». На одном конце спектра различных состояний находится полное бодрствование, на другом — глубокий сон. Хотя мы большую часть дня пребываем в состоянии бодрствования, степень нашего внимания непрерывно изменяется. Точно так же, когда мы спим, глубина нашего сна различна. Эта глубина, или фаза сна, тоже меняется. Относительную глубину сна можно оценить по характеру и амплитуде волн электрической активности, зафиксированной на электроэнцефалограмме (рис. 70).

Рис. 70. Проявление различных уровней бодрствования и сна в меняющемся характере и амплитуде мозговых волн, регистрируемых с помощью электродов, прикрепленных к коже головы. Такая запись — электроэнцефалограмма (ЭЭГ) — отражает активность большого числа нейронов в области мозга, расположенной ближе всего к поверхностным электродам. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных.
Фазы сна. У людей с помощью электроэнцефалографии можно выделить пять фаз, или степеней глубины, сна. Для человека, спокойно сидящего или лежащего с закрытыми глазами, характерен альфа-ритм, при котором частота регистрируемых «волн» составляет 8-12 Гц (фаза 1). Когда наступает сон, основной ритм замедляется, амплитуда отдельных пиков электрического потенциала снижается (фаза 2). Исследователи, изучающие сон, объясняют это флуктуациями более или менее случайной нейронной активности. На стадии более глубокого сна появляются эпизоды с большей частотой волн, время от времени прерывающие медленную низковольтную активность (фаза 3). Сонные веретена — это всплески активности с частотой 12-15 Гц. Продолжительность их не превышает одной секунды. По мере дальнейшего углубления сна колебания потенциала замедляются и преобладают уже дельта-волны с частотой около 4 Гц (фаза 4). И наконец, после достижения наиболее глубокой фазы сна электрическая активность вновь приобретает характер быстрых колебаний с низкой амплитудой; при этом появляются периоды, во время которых происходят быстрые сокращения глазных мышц. Эта пятая фаза, или сон с быстрыми движениями глаз (БДГ-сон), сопровождается также почти полным расслаблением скелетной мускулатуры. Люди, разбуженные во время этой фазы, часто говорят, что видели сны, поэтому фазу БДГ считают периодом сновидений. Судя по результатам других исследований, сновидения, вероятно, могут иметь место и во время глубокого медленно-волнового сна, например на стадии дельта-волн.
Биологические основы сна и бодрствования. Биологические основы активного бодрствования с трудом поддаются изучению. По мнению некоторых ученых, первый шаг к их выяснению состоит в том, чтобы установить, какие системы мозга ответственны за наступление сна и его отдельных стадий. Обычный подход при изучении локализации мозговых функций — стимуляция тех или иных участков мозга или их повреждение — помог определить, какие области наиболее тесно связаны со стадиями сна и бодрствования.
В начале 1950-х годов работы, проведенные итальянским физиологом Моруцци и американским физиологом Мэгуном, показали, что наиболее важной зоной является внутренняя область варолиева моста и ствола мозга. Согласно анатомическим данным, волокна от этой области идут вверх к ядрам таламуса, имеющим связи с корой. Существование таких связей означает, что ретикулярная формация моста и ствола могла бы эффективно воздействовать на кору больших полушарий. Электростимуляция ретикулярной формации моста действительно возбуждает в коре активность, регистрируемую электроэнцефалограммой. С другой стороны, повреждения моста ведут у животных к необратимому переходу в коматозное состояние.
Регистрируя активность определенных нейронов моста в области ретикулярной формации, в дальнейшем удалось установить, что активность некоторых из этих нейронов специфическим образом изменяется как раз перед сменой стадий сна. Например, у отдельных клеток перед началом фазы глубокого БДГ-сна резко возрастала частота импульсации — в 50-100 раз по сравнению со спокойным бодрствующим состоянием. Тот факт, что повышение частоты разряда начиналось за некоторое время до того, как на ЭЭГ появлялись признаки БДГ-сна, определенно свидетельствует в пользу участия этих клеток в событиях, приводящих к смене фазы.
Две другие группы нейронов варолиева моста обнаруживали тенденцию к противоположному изменению активности при смене фаз сна. Интерес к этим нейронам первоначально возник в связи с тем, что были получены некоторые данные о химической природе их медиаторов, наводившие на мысль о возможной роли этих клеток в регулировании сна. Это были две следующие группы: 1) скопление норадреналин-содержащих нейронов в области голубого пятна и 2) скопление серотонин-содержащих нейронов в дорсальном ядре шва (см. рис. 71). Как показали записи активности отдельных нейронов этих ядер, она максимальна во время бодрствования, постепенно снижается на ранних стадиях медленноволнового сна и к концу его почти полностью угасает; во время БДГ-сна эти нейроны бездействуют.

Рис. 71. Системы регуляции сна. Здесь показаны основные участки мозга, посылающие волокна к ретикулярной активирующей системе. Видно, как в цепях, выходящих из голубого пятна (медиатор — норадреналин) и ядер шва (медиатор — серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
Как теперь полагают, нейроны моста, активирующиеся на стадии БДГ-сна, и нейроны ядер голубого пятна и шва, неактивные во время этой фазы, связаны друг с другом. Можно было бы просто предположить, что первые, возбуждаясь, тормозят активность вторых. Однако эта гипотеза не позволяет понять, что затормаживает первую группу клеток в конце фазы БДГ и что потом восстанавливает их активность. Возможно, что сходный тип активности свойствен и многим другим, пока еще неисследованным нейронам моста и что характер их импульсации еще лучше коррелирует с чередованием фаз сна и бодрствования.
Эти исследования сослужили еще одну службу: они навели на мысль, что медиаторы серотонин и норадреналин играют определенную роль в регуляции общего уровня активности. Например, некоторые транквилизаторы вызывают не только длительное понижение активности и сонливость, но и истощение мозговых запасов серотонина и норадреналина. Кроме того, поражения ядер голубого пятна и дорсального ядра шва могут вызывать резкие нарушения нормального сна, и в особенности подавлять фазу БДГ.
Связи норадреналин- и серотонинэргических нейронов относятся к категории сетей дивергентного типа с одним входом (см. гл. 2). Из скоплений таких клеток их дивергирующие аксоны направляются во многие области мозга, и благодаря такому широкому распространению эти клетки могут влиять на множество других нейронов. Вот почему они потенциально способны играть роль в регуляции таких крупномасштабных явлений, как бодрствование и сон.
Детальные наблюдения над активностью нейронов голубого пятна тоже говорят в пользу высказанных предположений. У бодрствующих животных, взаимодействующих с внешним миром, наблюдаются краткие периоды усиленной активности этих нейронов во время переработки новой сенсорной информации, поступающей извне. Эти клетки активируются тогда, когда животное подвергается воздействию любого рода новых стимулов-прикосновений, света, звуков или запахов. Вместе с другими сведениями о действии норадреналина, выделяемого нейронами голубого пятна, эти данные позволяют предполагать, что понижение активности голубого пятна создает условия для наступления сна. Можно думать также, что с возбуждением этого участка связано появление кратких периодов повышенного реагирования на новые раздражители. Таково научное, неврологическое истолкование понятий «бодрствование» или «внимание».
Центральные интегрирующие механизмы
Клеточные механизмы, лежащие в основе таких сложных общих состояний, как внимание, бодрствование и сон, все еще далеко не ясны. Как показывают приведенные выше факты, о ряде групп нейронов было получено достаточно данных, чтобы можно было уже начать формулировать доступные для проверки гипотезы о некоторых клеточных взаимодействиях, лежащих в основе тех или иных форм поведения.
Однако весьма вероятно, что для понимания некоторых клеточных основ интегративных способностей мозга полезными вскоре окажутся молекулярные объяснения (см. гл. 7). Известно, например, что в регуляции переходов из одного общего состояния в другое участвуют, вероятно, и другие системы, организованные по типу дивергентных сетей с одним входом. Одну из таких систем образуют дофаминэргические нейроны моста, другую — холинэргические (выделяющие ацетилхолин) нейроны промежуточного мозга и ретикулярной формации моста. Обе нейронные системы посылают аксоны к множеству клеток-мишеней и, по-видимому, служат для координации их деятельности.
Гиперактивность некоторых дофаминэргических нейронов характерна для больных шизофренией, у которых неоправданные сопоставления сенсорных данных приводят к искаженному восприятию событий (см. гл. 9). Снижение активности холинэргических, а также адренэргических (выделяющих норадреналин) нейронов может наблюдаться при болезни Альцгеймера, для которой характерна неспособность больных соотносить текущую сенсорную информацию с прошлым опытом (см. гл. 10).
Эти системы наряду со многими другими системами моста и гипоталамуса, организованными по типу дивергентных сетей с одним входом, медиаторы которых пока неизвестны, обладают такой структурой, которая почти наверняка позволяет им осуществлять интегративные функции. Возможно, есть и иные типы нейронных сетей, а также иные особенности действия медиаторов, которые тоже обеспечивают координацию работы сенсорных и двигательных подсистем.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
2. ЗРИТЕЛЬНАЯ СИСТЕМА
2. ЗРИТЕЛЬНАЯ СИСТЕМА Зрение – один из важнейших органов чувств человека. Оно эволюционно приспособлено к восприятию узкой части диапазона электромагнитных излучений (видимый свет). Зрительная система даёт мозгу более 90% сенсорной информации. Зрение – это
3. СЛУХОВАЯ СИСТЕМА
3. СЛУХОВАЯ СИСТЕМА В связи с возникновением речи как средства межличностного общения, слух у человека играет особую роль. Акустические (звуковые) сигналы представляют собой колебания воздуха с разной частотой и силой. Они возбуждают слуховые рецепторы, находящиеся в
4. ВЕСТИБУЛЯРНАЯ СИСТЕМА
4. ВЕСТИБУЛЯРНАЯ СИСТЕМА Вестибулярная система играет важную роль в пространственной ориентации человека. Она получает, передаёт и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при
5. СОМАТОСЕНСОРНАЯ СИСТЕМА
5. СОМАТОСЕНСОРНАЯ СИСТЕМА В соматосенсорную систему входят система кожной чувствительности и чувствительная система скелетно-мышечного аппарата, главная роль в которой принадлежит
7. ВКУСОВАЯ СИСТЕМА
7. ВКУСОВАЯ СИСТЕМА В процессе эволюции вкус формировался как механизм выбора или отвергания пищи. Выбор предпочитаемой пищи отчасти основан на врождённых механизмах, но в значительной мере зависит от связей, выработанных в онтогенезе. Вкус, так же как и обоняние, основан
2.1. Что такое система?
2.1. Что такое система? Термин «система» обычно применяется для того, чтобы указать на собранность, организованность группы элементов и отграниченность её от других групп и элементов. Давалось множество определений системы, которые характеризовали её, выделяли из
Эндокринная функция половых желез
Эндокринная функция половых желез На функции половых желез влияют релизинг-гормоны гипофиза и гормоны, выделяемые непосредственно половыми железами.Мужские половые гормоны.Гормоны, выделяемые мужскими половыми железами по своей природе являются стероидами и
1.3. Система человекознания
1.3. Система человекознания Ни один объект живой природы не интересует человека в такой степени, как он сам. Вот почему многие науки занимаются этим «объектом». Оразовалась определенная совокупность знаний о человеке, которую можно назвать «человекознанием». Однако из-за
Система приматов
Система приматов Направление, ведущее к современному отряду приматов, выделилось в классе млекопитающих еще в мезозойскую эру, около 70 млн лет назад, являясь одним из древнейших. Уже в то время наметилась магистральная линия их эволюции – развитый головной мозг,
Система FCI:
Система FCI: В соответствии с решением Генерального собрания FCI в Иерусалиме в 1987 году принято следующее деление зарегистрированных пород собак на группы, которое вступило в силу с 1 января 1990 года.1 группа — овчарки (пастушьи и скотогонные собаки, за исключением
Нервная система
Нервная система Как известно, нервная система впервые появляется у низших многоклеточных беспозвоночных. Возникновение нервной системы — важнейшая веха в эволюции животного мира, и в этом отношении даже примитивные многоклеточные беспозвоночные качественно
Эндокринная система
Эндокринная система Эндокринный орган отличается тем, что выделяет вещество, необходимое для регуляции клеточной активности каких-то других органов, непосредственно в кровяное русло (термин происходит от греч. endo — внутри и krinein — выделять). Такие органы называются
6.3. Эндокринная функция двенадцатиперстной кишки
6.3. Эндокринная функция двенадцатиперстной кишки Как отмечено выше, к началу 50-х годов нами были преодолены технические трудности, связанные с полным и атравматичным удалением двенадцатиперстной кишки и с пересадкой панкреатического и общего желчного протоков в тощую
6.3. Эндокринная функция двенадцатиперстной кишки
6.3. Эндокринная функция двенадцатиперстной кишки Как отмечено выше, к началу 50-х годов нами были преодолены технические трудности, связанные с полным и атравматичным удалением двенадцатиперстной кишки и с пересадкой панкреатического и общего желчного протоков в тощую