Рассказ Плодовой Мушки
Рассказ Плодовой Мушки
В 1894 году пионер генетики Уильям Бэтсон (William Bateson) издал книгу под названием «Материалы по изучению прерывистой изменчивости в происхождении видов». Он собрал очаровательный, почти жуткий список генетических отклонений, и рассмотрел, как они могли бы пролить свет на эволюцию. У него были лошади с раздвоенными копытами, антилопы с единственным рожком в середине головы, люди с лишней рукой и жук с пятью ногами с одной стороны. В своей книге Бэтсон выдумал термин «гомеозис» для замечательного типа генетической вариации. Homoio означает по-гречески «остающийся таким же», и гомеотическая мутация (как мы теперь назвали бы ее, хотя, когда Бэтсон писал, термин «мутация» не был введен в обращение) – та, которая заставляет часть тела появляться на месте какой-либо другой части.
Нарисованный Бэтсоном жук с гомеотической мутацией, издано в 1894 году [18].
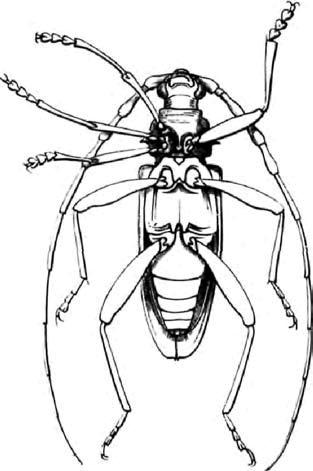
Собственные примеры Бэтсона включали пилильщика с ногой, растущей на месте, где должна была быть антенна. Только услышав об этой замечательной аномалии, Вы могли бы подозревать, вместе с Бэтсоном, что здесь должна быть важная подсказка к тому, как животные обычно развиваются. Вы с Бэтсоном были бы правы, и это – предмет настоящего рассказа. Такой специфический гомеозис – нога вместо антенны – был позже обнаружен у плодовой мушки дрозофилы и назван антеннопедия. Дрозофила («любительница росы») долго была любимым животным генетиков. Эмбриологию никогда не нужно путать с генетикой, но недавно дрозофила приняла на себя главную роль в эмбриологии, так же как в генетике, и это – рассказ об эмбриологии.
Эмбриональным развитием управляют гены, но существует два совершенно разных пути, которыми оно могло бы теоретически происходить. «Рассказ Мыши» представил их как проект и рецепт. Строитель сооружает дом, кладя кирпичи в места, определенные проектом. Повар делает пирог, не помещая крошки и смородины в указанные места, а внося компоненты через указанные процедуры, такие как просеивание, взбалтывание, взбивание и нагревание (Эта любимая аналогия сначала использовалась моим другом сэром Патриком Бэтсоном (Patrick Bateson), между прочим, родственником сэра Уильяма.). Учебники биологии ошибаются, если описывают ДНК как проект. Эмбрионы не делают ничего, отдаленно напоминающего следование проекту. ДНК не является описанием на любом языке, того, на что должно быть похоже законченное тело. Возможно, на какой-то другой планете живые существа развиваются с помощью эмбриологии проекта, но я считаю, что трудно представить, как бы она работала. Это должен быть совсем иной вид жизни. На нашей планете эмбрионы следуют за рецептами. Или, чтобы заменить это на другую, столь же несхожую с проектом аналогию, которая до некоторой степени подходит больше, чем рецепт: эмбрионы создаются, следуя за последовательностью оригами, инструкцией по складыванию бумажных фигурок.
Аналогия с оригами соответствует ранней эмбриологии лучше, чем поздней. Главная организация тела первоначально заложена рядом сворачиваний и выпячиваний слоев клеток. Как только главная схема тела благополучно оказывается на месте, более поздние стадии развития представляют в значительной степени рост, как будто эмбрион раздувается со всех сторон, как воздушный шар. Впрочем, это – очень специфичный вид воздушного шара, потому что различные части тела раздуваются с разной скоростью, тщательно управляемой скоростью. Это важное явление известно как аллометрия. «Рассказ Плодовой Мушки» имеет отношение главным образом к ранней, оригами-фазе развития, а не к более поздней, инфляционной.
Клетки не укладываются, как кирпичи по проекту, но именно поведение клеток определяет эмбриональное развитие. Клетки притягивают или отталкивают другие клетки. Они изменяют форму различными способами. Они выделяют химические вещества, которые могут распространиться за ее пределы и влиять на другие клетки даже на некотором расстоянии. Иногда они выборочно погибают, создавая формы удалением, как будто при работе скульптора. Как термиты, сотрудничающие, чтобы построить термитник, клетки «знают» что делать под влиянием соседних клеток, с которыми они оказываются в контакте, и благодаря градиенту концентрации химических веществ. Все клетки в эмбрионе содержат одни и те же гены, таким образом, не может быть, чтобы гены приводили к различиям в поведении одних клеток относительно других. Что действительно распознает клетка – какие из генов включены, что обычно отражается в генных продуктах, белках, которые она содержит.
В очень раннем эмбрионе клетка должна «знать», где она находится относительно двух главных направлений: спереди или сзади (передне-заднее направление), и вверху или внизу (спинно-брюшное). Что значит «знать»? В начальной стадии это означает, что поведение клетки определяется ее положением вдоль химических градиентов относительно каждой из этих двух осей. Такие градиенты неизбежно начинаются непосредственно в яйце, и находятся, поэтому, под контролем генов матери, а не собственных ядерных генов яйца. Например, существует ген, названный bkoid в материнском генотипе дрозофилы, который экспрессируется в «клетках-няньках», строящих ее яйца. Белок, созданный геном bicoid, транспортируется в яйцо, где концентрируется в одном конце, постепенно исчезая к другому. Итоговый градиент концентрации (и другие похожие) определяет передне-заднюю ось. Аналогичные механизмы под прямым углом определяют спинно-брюшную ось.
Эти маркирующие концентрации сохраняются в веществе клеток, которые образуются, когда яйцо впоследствии делится. Первые несколько делений происходят без какого-либо прибавления нового материала, и деления являются неполными: создается много отдельных ядер, но они полностью не отделены клеточными перегородками. Эту многоядерную «клетку» называют синцитий. Позже перегородки формируются, и эмбрион становятся по-настоящему клеточными. В продолжение всего этого, как я сказал, сохраняются первоначальные химические градиенты. Из этого следует, что ядра клеток в различных частях эмбриона будут плавать в различных концентрациях ключевых веществ, соответствующих первоначальным двумерным градиентам, и это заставит различные гены включаться в различных клетках (мы сейчас, конечно, не говорим о собственных генах эмбриона, больше о материнских). Так начинается дифференциация клеток, и проецирование этого принципа приводит к дальнейшей дифференциации на более поздних стадиях развития. Первоначальные градиенты, настроенные материнскими генами, уступают дорогу новым и более сложным градиентам, настроенным собственными генами эмбриона. Последовательные разветвления в линиях эмбриональных клеток рекурсивно производят дальнейшие дифференциации.
У членистоногих существует более широкомасштабное разделение тела, не на клетки, а на сегменты. Сегменты выстраиваются в линии, от передней части головы до конца брюха. У насекомых есть шесть главных сегментов, в которых антенны находятся на сегменте 2, на следующих сегментах находятся мандибулы, а затем другие ротовые части. Сегменты головы взрослого сжаты до маленьких пределов, таким образом, их расположение вдоль всей длины тела не слишком ясно, но его можно увидеть в эмбрионе. Три грудных сегмента (T1, T2 и T3), расположенных в ряд, более заметны, каждый имеет пару ног. T2 и T3 обычно имеют крылья, но у дрозофилы и других мух крылья есть только на T2 (Некоторые другие насекомые, такие как тараканы и жуки, летают только с помощью крыльев T3, изменив крылья T2 в твердые защитные надкрылья, названные элитра. Сверчки и кузнечики, как мы слышали, затем изменили элитры в звуковоспроизводящие органы.). Вторая пара «крыльев» изменена в жужжальца, маленькие булавообразные органы на T3, которые вибрируют и служат миниатюрными гироскопами, направляющими муху. У некоторых ранних ископаемых насекомых было три пары крыльев, одна пара на каждом из трех грудных сегментов. Позади грудных сегментов расположено большее число брюшных сегментов (11 у некоторых насекомых, восемь у дрозофилы, в зависимости от того, принимаете ли Вы во внимание гениталии на заднем конце). Клетки «знают» (в уже оправданном смысле), в каком сегменте они находятся, и они ведут себя соответственно. Каждая клетка различает, в каком сегменте она расположена, при посредничестве специальных регулирующих генов, Hox-генов, включенных в клетку. «Рассказ Плодовой Мушки» – главным образом рассказ о Hox-генах.
Вопрос бы стал ясным и легко объяснимым, если бы я мог теперь сказать Вам, что есть один Hox-ген для каждого сегмента, при этом все клетки данного сегмента имеют только свой собственный, пронумерованный, включенный в нее Hox-ген. Было бы еще лучше, если бы Hox-гены выстраивались вдоль хромосомы в том же самом порядке, что и сегменты, на которые они влияют. Что ж, это не столь здорово, но почти так. Hox-гены действительно расположены в правильном порядке вдоль одной хромосомы, и это замечательно – учитывая то, что мы знаем о работе генов. Но Hox-генов не достаточно для сегментов – только восемь. И есть более неприятное затруднение, которое я должен устранить. Сегменты взрослого не точно соответствуют так называемым парасегментам личинки. Не спрашивайте меня, почему (возможно, у Проектировщика был неудачный день), но каждый сегмент взрослого составлен из задней половины одного личиночного парасегмента плюс передней половины следующего. Если отдельно не оговорено, я буду использовать слово «сегмент», чтобы обозначить личиночный (пара-) сегмент. Что касается вопроса о том, как восемь Hox-генов подряд ответственны приблизительно за 17 подряд сегментов, это, отчасти, снова происходит благодаря уловке с химическим градиентом. Каждый Hox-ген в основном экспрессируется в одном сегменте, но он также экспрессируется, в уменьшающейся степени, если двигаться назад, в большом количестве следующих сегментов. Клетка знает, в каком сегменте она находится, сравнивая химические продукты более чем одного выше расположенного Hox-гена. Это немного более сложно, но здесь нет никакой потребности вдаваться в такие детали.
Восемь Hox-генов выстраиваются в два генных комплекса, физически разделенных вдоль одной и той же хромосомы. Их называют комплексом Antennapedia и комплексом Bithorax. Эти названия вдвойне неудачны. Комплекс генов называют в честь единственного члена этого комплекса, который не более важен, чем другие. Хуже того, сами гены обычно называют по названию того, что случается, если они работают не так как надо, а не по названию их нормальной функции. Было бы лучше назвать их чем-то вроде передний Hox-комплекс и задний Hox-комплекс. Однако мы упорно придерживаемся существующих названий.
Комплекс Bithorax состоит из последних трех Hox-генов, названных по историческим причинам в которые я не буду вдаваться, Ultrabithorax, Abdominal-A и Abdominal-B. Они затрагивают задний конец животного следующим образом. Ultrabithorax экспрессируется от сегмента 8 по всей длине до заднего конца. Abdominal-A экспрессируется от сегмента 10 до конца, и Abdominal-B экспрессируется от сегмента 13 до конца. Продукты этих генов образуются в уменьшающемся градиенте концентрации в направлении заднего конца животного от своих различных отправных точек. Так, сравнивая концентрации продуктов этих трех Hox-генов, клетка в следующей части личинки может различить, в каком сегменте она находится, и действовать соответственно. Та же история для переднего конца личинки, за который ответственны пять Hox-генов комплекса Antennapedia.

Клетки «думают», что они находятся в другом сегменте. Гомеотическая мутация плодовой мушки.
В таком случае Hox-ген является геном, чье жизненное предназначение состоит в том, чтобы знать свое местонахождение в теле и сообщать о нем другим генам в этой же клетке. Мы теперь готовы понять гомеотические мутации. Когда Hox-ген работает не так как надо, клетки в сегменте дезинформированы, в каком сегменте они находятся, и создают тот сегмент, в котором они «думают», что находятся. Так, например, мы видим, что нога растет в сегменте, в котором обычно выращивалась бы антенна. В этом есть точный смысл. Клетки в любом сегменте вполне способны к сборке структуры любого другого сегмента. Почему бы им этого не делать? Инструкции для изготовления любого сегмента скрываются в клетках каждого из сегментов. Это – Hox-гены, которые, при нормальных условиях, вызывают «правильные» инструкции для создания структуры, соответствующей каждому сегменту. Как справедливо подозревал Уильям Бэтсон, гомеотическая аномалия открывает показательное окно на то, как обычно работает система.
Вспомните, что у мух, необычно для насекомых, как правило, есть только одна пара крыльев, плюс пара гироскопических жужжалец. Гомеотическая мутация Ultrobithorax вводит в заблуждение клетки в третьем грудном сегменте, которые «думают», они находятся во втором грудном сегменте. Поэтому они сотрудничают, чтобы создать дополнительную пару крыльев, вместо пары жужжалец (см. рис.) Есть мутант мучного хрущака (Tribolium), у которого все 15 сегментов развились в антенны, по-видимому, потому что все клетки «думают», что они находятся в сегменте 2.
Это переносит нас в самую замечательную часть «Рассказа Плодовой Мушки». После того, как Hox-гены были обнаружены у дрозофилы, они стали выявляться повсеместно: не только у других насекомых, таких как жуки, но и почти у всех других животных, которых исследовали, включая нас самих. И – что действительно едва ли не слишком хорошо, чтобы быть правдой – они, очень часто, оказывается, делают то же самое, информируют делящиеся на сегменты клетки о том, где они находятся, и (даже лучше) выстраиваются в том же порядке вдоль хромосом. Давайте теперь вернемся к истории млекопитающих, которая была наиболее тщательно изучена благодаря лабораторной мыши – этой дрозофиле мира млекопитающих.
Млекопитающие, как и насекомые, обладают сегментированным планом тела, или, по крайней мере, модульным, повторяемым планом, который затрагивает основу и связанные структуры. Каждый позвонок можно представить себе как соответствующий одному сегменту, но не только кости повторяются по направлению от шеи к хвосту. Кровеносные сосуды, нервы, блоки мышц, хрящевые диски и ребра, где они есть, все придерживаются повторяющегося, модульного плана. Модульного, как у дрозофилы. Хотя у нижеупомянутой, связанной отдаленным родством дрозофилы есть такие же серии вдоль тела, как у млекопитающих, безусловно, вызывало бы беспокойство, если бы их не было также у ланцетника. Мой коллега Питер Холланд со своей исследовательской группой изучил этот вопрос, и их результаты удовлетворительны. Да, модульный план тела ланцетника опосредован (14) Hox-генами, и да, они расположены в правильном порядке вдоль хромосомы. В отличие от мыши, но подобно дрозофиле, существует только один ряд, а не четыре параллельных. По-видимому, вся группа была дублирована четыре раза где-нибудь вдоль линии, ведущей от Копредка 23 к современным млекопитающим, что сопровождалось некоторыми спорадическими потерями отдельных генов.
А как же другие животные, стратегически выбранные потому, что они могут рассказать нам о другом отдельном копредке? Hox-гены были теперь найдены у каждого животного, которого исследовали, кроме гребневиков и губок (см. Свидание 29 и 31 соответственно), включая морских ежей, мечехвостов, креветок, моллюсков, кольчатых червей, кишечнодышащих, асцидий, нематод и плоских червей. Мы могли это предположить, зная, что все эти животные происходят от Копредка 26, и у нас уже есть серьезные основания считать, что у Копредка 26 были Hox-гены, как у его потомков дрозофилы и мыши.
Книдарии, такие как гидра (они не должны присоединиться к нам до Свидания 28), являются радиально-симметричными – у них нет ни передне-задней оси, ни верхне-нижней. У них есть орально-аборальная ось. Не очевидно, во всяком случае, что она соответствует их длинной оси, поэтому каких действий можно было бы ожидать от их Hox-генов? Было бы неплохо, если бы они использовались для определения орально-аборальной оси, но пока не ясно, что это так. Так или иначе, у большинства книдарий есть только два Hox-гена, против восьми у дрозофилы и четырнадцати у ланцетника. Приятно, что один из этих двух генов напоминает передний комплекс дрозофилы, в то время как другой напоминает задний. У Копредка 28, которого мы разделяем с ними, по-видимому, было то же самое. Затем один из них был дуплицирован несколько раз в течение эволюции, создав комплекс Antennapedia, в то время как другой был дуплицирован в той же линии животных, создав комплекс Bithorax. Это как раз является своего рода способом увеличения генов в геноме (см. «Рассказ Миноги»). Но необходимо больше исследований, прежде чем мы узнаем, что вообще эти два гена делают при планировании тела книдарии.
Иглокожие радиально-симметричны, как киндарии, но радиально-симметричны вторично. Копредок 25, которого они разделяют с нами, позвоночными, был двусторонне-симметричным, как червь. У иглокожих различное количество Hox-генов – десять в случае морских ежей. Что делают эти гены? Скрывается ли в теле морской звезды пережиток наследственной передне-задней оси? Или Hox-гены проявляют свое влияние последовательно вдоль каждого пяти ее лучей? Это, казалось бы, имело бы смысл. Мы знаем, что Hox-гены экспрессируются в передних и задних лапах млекопитающих. Я не имею в виду, что множество Hox-генов от 1 до 13 экспрессируются по очереди, от плеча до кончиков пальцев. Это не так просто – как оно могло бы быть – потому что конечность позвоночного устроена не из модулей, которые следуют друг за другом на ее протяжении. Вместо этого там вначале находится одна кость (плечевая в передней конечности, бедренная в задней), затем две кости (лучевая и локтевая кость в передней конечности, голень и малоберцовая кость в задней), затем множество небольших костей достигают наибольшего количества в пальцах передней и задней конечности. Это веерное расположение, унаследованное от более очевидного веера плавников наших рыбоподобных предков, не предоставляет собой простой линейной зависимости Hox. Даже в этом случае Hox-гены вовлечены в развитие конечностей позвоночных.
По аналогии не было бы удивительно, если бы Hox-гены также экспрессировались в лучах морской звезды или офиуры (и даже морских ежей можно представить как о морских звезд, лучи которых завернулись вверх в пятиконечный свод, соединились концами и застегнули по бокам «молнии»). Кроме того, лучи морской звезды, в отличие от наших рук или ног, действительно являются последовательно модульными на своем протяжении. Трубчатые ножки со всей связанной с ними водопроводной системой являются структурными единицами, которые повторяются в двух параллельных рядах вдоль каждого луча: как раз то, что нужно для экспрессирования Hox-генов! Лучи офиур даже выглядят и ведут себя, как пять червей.
T. Г. Хаксли упомянул «большую трагедию науки – сражение красивой гипотезы уродливым фактом». Истинные факты Hox-генов иглокожих, возможно, не уродливы, но они не следуют за симпатичным примером, который я только что предложил. Происходит нечто другое, что имеет свою собственную, довольно удивительную красоту. Личинки иглокожего – крошечные, двусторонне-симметричные, плавающие в планктоне животные. Пятиконечное, радиально симметричное, живущее на дне взрослое животное не развивается как трансформация личинки. Вместо этого оно возникает как крошечный миниатюрный взрослый в теле личинки, который растет, пока, в конечном счете, остальная часть личинки не отбрасывается. Hox-гены экспрессируются в правильном линейном порядке, но не вдоль каждого луча. Вместо этого порядок экспрессирования следует по приблизительно круглому маршруту вокруг маленького взрослого. Если мы представили себе ось Hox как «червя», не существует пяти «червей», по одному для каждого луча. Есть единственный «червь», скрученный вокруг внутренней части личинки. Передний конец «червя» выращивает луч номер 1, задний конец «червя» выращивает луч номер 5. В таком случае, гомеотические мутации у морской звезды, как можно было бы ожидать, вырастят слишком много лучей. И без сомнения, мутантная морская звезда с шестью лучами известна и была зарегистрирована в книге Бэтсона. Есть также некоторые виды морских звезд, у которых намного большее количество лучей, и они, по-видимому, эволюционировали от гомеотических мутантных предков.
Hox-гены не были найдены ни у растений, ни у грибов, ни у одноклеточных организмов, обычно называемых простейшими. Но теперь мы подошли к трудности в терминологии, с которой необходимо справиться прежде, чем мы двинемся дальше. «Hox» был придуман как сокращение от «гомеобокс» (homeobox), но Hox-гены не синонимичны с гомеобокс-генами: они – их подмножество. У растений и грибов действительно есть гомеобокс-гены, но у них нет Hox-генов (Растения и грибы должны иметь системы контролирующих генов и химических градиентов, чтобы превратиться в правильную форму. MADS-box гены определяют эмбриологию цветов и могут произвести гомеотические мутации в цветах таким же образом, как Hox-гены у животных.).
«Гомео» происходит от «гомеозиса» Бэтсона, а «бокс» указывает на блок из 180 кодовых знаков, который есть у всех генов, известных как гомеобокс-гены, где-нибудь на их протяжении. Сам гомеобокс – это диагностическая последовательность из 180 кодовых знаков, а «гомеобокс-ген» – ген, который содержит последовательность гомеобокс где-нибудь на своем протяжении. Название Hox используется не для всех гомеобокс-генов, а только для линейных множеств генов, которые определяют положение вдоль тела животного и которые оказались гомологичными почти у всех животных.
Hox-семья гомеобокс-генов была первой из обнаруженных, но теперь известно много родственных семей. Например, существует семья генов под названием ParaHox, которая была сначала четко установлена у ланцетников, но которая, кроме того, встречается у всех животных, кроме (пока) гребневиков и губок. Кажется, что ParaHox-гены – «кузены» Hox-генов, в смысле, что они им соответствуют и расположены в том же порядке, что и Hox-гены. Они, конечно, возникли при дупликации от одного и того же предкового набора генов, что и Hox-гены. Другие гомеобокс-гены более отдаленно связаны с Hox и ParaHox, но формируют собственные семьи. Семья Pox обнаружена у всех животных. Особенно известен член этой семьи – Pax6, который соответствует гену, известному как ey у дрозофилы. Я уже упомянул, что Pax6 ответственен за приказ клеткам создавать глаза. Тот же ген делает глаза у столь разных животных, как дрозофила и мышь, даже притом, что созданные глаза у этих двух животных радикально различны. Подобно Hox-генам, Pax6 не сообщает клеткам, как сделать глаз. Он только говорит им, что здесь место, чтобы сделать глаз.
Довольно похожий пример – маленькая семья генов, названных tinman. Снова же, гены tinman присутствуют и у дрозофилы, и у мыши. У дрозофилы гены tinman ответственны за приказ клеткам делать сердце, и они обычно экспрессируются только в надлежащем месте, чтобы создать сердце дрозофилы. Как мы теперь можем ожидать, гены tinman также участвуют в приказах клеткам мыши создавать сердце в надлежащем для него месте.
Весь набор гомеобокс-генов представляет собой очень большое множество, разделенное на семейства и подсемейства, так же как сами животные разделены на семейства и подсемейства. Это похоже на случай гемоглобина, который мы исследовали в «Рассказе Миноги». Там мы узнали, что человеческий альфа-глобин действительно является более близким кузеном, скажем, альфа-глобина ящерицы, чем кузеном человеческого бета-глобина, который, в свою очередь, более близкий кузен бета глобина ящерицы. Точно так же человеческий tinman – более близкий кузен tinman плодовой мушки, чем человеческого Pax6. Есть возможность построить очень полное генеалогическое дерево гомеобокс-генов, которое существует бок о бок с генеалогическим деревом содержащих их животных. Оба генеалогических дерева одинаково обоснованны. Оба – настоящие родословные деревья, сформированные событиями расколов, которые случались в особые моменты геологической истории. В случае генеалогических деревьев животных события расколов представляют собой видообразования. В случае генеалогических деревьев гомеобогс-генов (или генов глобина), события расколов – дупликации гена в геномах.
Дерево гомеобокс-гена животных раскалывается на два больших класса, на AntP и PRD классы. Я не буду обстоятельно объяснять, что обозначают эти сокращения, потому что оба являются упрямо сбивающими с толку. Класс PRD включает Pax-гены и различные другие подклассы. Класс AntP включает Hox и ParaHox, и тоже различные другие подклассы. В дополнение к этим двум большим классам гомеобокс-генов животных существуют различные, более отдаленно связанные гомеобокс-гены, которые (ошибочно) называют «дивергентными». Они обнаружены не только у животных, но и у растений, грибов, а также у «простейших».
Только животные имеют истинные Hox-гены, и они всегда используются для одной и той же цели – чтобы установить информацию о местоположении внутри тела, независимо от того, разделено ли тело четко на дискретные сегменты. Хотя Hox-гены еще не были найдены у губок и гребневиков, это не означает, что такого не случится. Было бы не удивительно обнаружить, что они есть у всех животных. Это воодушевило бы моих коллег Джонатана Слэка (Jonathan Slack), Питера Холланда и Кристофера Грэма (Christopher Graham), тогда всех из Оксфорда, предложивших новое определение самого понятия «животное». До настоящего времени животные были определены в противоположность растениям, довольно неудовлетворительным негативным способом. Слэк, Холланд и Грэм предложили позитивный, особый критерий, имеющий результатом объединение всех животных и исключающий всех неживотных, таких как растения и простейшие. История с Hox показывает, что животные – не очень пестрая, несвязанная смесь типов, каждый со своим собственным фундаментальным планом тела, приобретенным и сохраненным в одинокой изоляции. Если Вы забудете морфологию и посмотрите только на гены, выяснится, что все животные – незначительные вариации на очень специфическую тему. Какое удовольствие быть зоологом в такое время.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Рассказ Евы
Рассказ Евы Есть впечатляющая разница между «родословными деревьями генов» и «родословными деревьями людей». В отличие от человека, который происходит от двух родителей, у гена есть только один родитель. Каждый из Ваших генов должен был быть получен или от Вашей матери,
Рассказ Секвойи
Рассказ Секвойи Люди спорят о том, какое одно место в мире Вы должны посетить прежде, чем умрете. Мой кандидат – лес Muir Woods, несколько севернее моста «Золотые Ворота». Или, если Вы считаете, что слишком поздно, я не могу вообразить лучшего места, чтобы быть похороненным (вот
Рассказ Taq
Рассказ Taq Достигнув нашего самого древнего свидания, собрав в нашем странствии всю жизнь, которую знаем, мы имеем возможность рассмотреть ее разнообразие. На самом глубинном уровне разнообразие жизни является химическим. Профессии, которыми заняты наши
Рассказ Stw 573
Рассказ Stw 573 Не думаю, что есть смысл придумывать причины, в силу которых ходить на двух ногах – это здорово. Будь так, шимпанзе делали бы то же самое, не говоря уже о других животных. Нет причины, по которой бег на двух или четырех конечностях быстрее или удобнее.
Рассказ Гориллы
Рассказ Гориллы Становление дарвинизма, которое пришлось на XIX век, привело к появлению двух противоположных взглядов на человекообразных обезьян. Противники Дарвина, хотя и согласившиеся принять идею эволюции, были в ужасе от возможного родства с грубыми,
Рассказ Орангутана
Рассказ Орангутана Возможно, заявление о наших давних связях с Африкой было поспешным. Что если наши предки покинули Африку около 20 млн лет назад и поселились в Азии, а 10 млн лет назад вернулись в Африку?Если так, то современные человекообразные обезьяны, включая тех,
Рассказ Гиббона[12]
Рассказ Гиббона[12] На рандеву № 4 мы встречаем крупную группу пилигримов. И теперь могут возникнуть проблемы с установлением родства. (Чем дальше, тем затруднительнее это сделать.) Существует двенадцать видов гиббонов, принадлежащих к четырем основным группам. Это Bunopithecus
Рассказ Айе-айе
Рассказ Айе-айе Один британский политик как-то описал конкурента (который позднее стал лидером партии) как человека, “в котором есть что-то ночное”. При взгляде на айе-айе складывается именно такое впечатление, и это не случайно: она ведет целиком ночной образ жизни. Из
Рассказ Шерстокрыла
Рассказ Шерстокрыла Шерстокрыл из Юго-Восточной Азии мог бы рассказать нам, каково это – парить во воздуху ночного леса. Но для нас, пилигримов, у него приготовлена гораздо более приземленная история. Мораль ее в том, что составленная нами схема с сопредками, рандеву и
Рассказ Мыши
Рассказ Мыши Из тысяч грызунов домашняя мышь (Mus musculus) может поведать кое-что особенное: ни один другой вид млекопитающих, кроме нашего собственного, мы не изучали так интенсивно. Именно на мышах, а не на пресловутых морских свинках держатся медицинские, физиологические и
Рассказ Бобра
Рассказ Бобра “Фенотип” – это то, что находится под влиянием генов. В целом под фенотипом понимают все, что относится к телу. Но есть тонкости, вытекающие из этимологии. Phamo по-гречески означает “показывать”, “обнаруживать”, “выявлять”, “выражать”, “раскрывать”.
Рассказ Гиппопотама
Рассказ Гиппопотама В школе, изучая греческий, я узнал, что “гиппос” означает “лошадь”, а “потамос” – “река”. Выходило, что гиппопотамы – “речные лошади”. Позднее, забросив греческий и занявшись зоологией, я узнал, что гиппопотамы не имеют отношения к лошадям. В
Рассказ Тюленя
Рассказ Тюленя В большинстве естественных популяций число самцов и самок примерно одинаково. На то есть эволюционные причины, и о них рассказал замечательный специалист по статистике и эволюционной генетике Рональд Э. Фишер. Представьте себе популяцию, количество в
Рассказ Броненосца
Рассказ Броненосца С зоологической точки зрения Южная Америка сродни Мадагаскару: как и Мадагаскар, она откололась от Африки – но не с восточной, а с западной стороны. Произошло это примерно тогда же или чуть позднее. Как и Мадагаскар, в течение почти всего времени
Рассказ Утконоса
Рассказ Утконоса Старое латинское название утконоса – Ornithorhynchus paradoxus [“парадоксальный птицеклюв”]. Когда его обнаружили, он показался ученым настолько нелепым, что присланный в музей экземпляр сочли чучелом из сшитых вместе частей тела млекопитающего и птицы. Кое-кто
Рассказ Taq
Рассказ Taq Итак, мы встретили практически все существующие формы жизни. Теперь можно окинуть взглядом открывшееся разнообразие. На самом глубоком уровне разнообразие жизни является химическим. Профессии, которыми владеют наши пилигримы, охватывают широкий диапазон