Рассказ Бархатного Червя
Рассказ Бархатного Червя
Если современная зоология признает что-нибудь похожее на полноценный миф о зарождении, то это – Кембрийский взрыв. Кембрийский период – первый период фанерозойского эона, последние 545 миллионов лет, в течение которых животная и растительная жизнь, как мы знаем, внезапно стала появляться в окаменелостях. До кембрийского периода окаменелости были или крошечными следами, или загадочными тайнами. От начала кембрийского периода был шумный зверинец многоклеточной жизни, более или менее правдоподобно предваряющий нашу собственную. Внезапность, с которой многоклеточные ископаемые появляются в начале кембрийского периода, внушает метафору взрыва.
Креационисты любят Кембрийский взрыв, потому что он, похоже, вызывает в их тщательно доведенном до нищеты воображении своего рода палеонтологический приют, населенный сиротскими типами: животными без прошлого, как будто они внезапно неожиданно материализовались из ничего, вместе с дырками в своих носках (Бертран Рассел, конечно.). В другой крайности, перегретые романтикой зоологи любят Кембрийский взрыв за его ауру «аркадской идиллии», зоологического возраста невинности, в котором жизнь танцевала в бешенном и радикально отличном эволюционном темпе: безгрешная вакханалия скачущей импровизации перед падением в серьезный прагматизм, который преобладает с тех пор. В «Расплетая радугу» я цитировал следующие слова выдающегося биолога, который, возможно, к настоящему времени, изменил свое мнение:
Вскоре после того, как были изобретены многоклеточные формы, разразился великий взрыв эволюционной новизны. Каждый почти постиг смысл многоклеточной жизни, радостно опробуя все ее возможные ветвления в своего рода диком танце беспечного исследования.
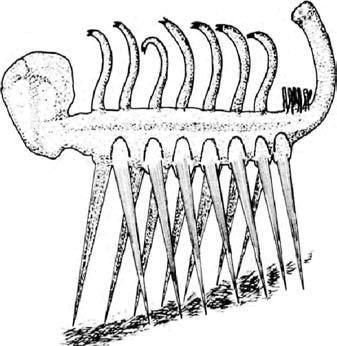
Бредовый взгляд на кембрийскую галлюцигению – реконструирована вверх тормашками.
Если есть животное, ставшее в большей степени, чем другие, причиной бредового взгляда на кембрийский период, это – галлюцигения. На чем она стоит? Не принимая во внимание галлюцинаций, Вы могли бы подозревать, что такое маловероятное существо никогда в своей жизни не стояло. И Вы были бы правы. Оказывается галлюцигения – а Саймон Конвей Моррис (Simon Conway Morris) выбрал это название намеренно – была первоначально реконструирована вверх тормашками. Именно поэтому она стоит на этих невероятных ходулях- зубочистках. Одинокий ряд «щупалец» вдоль спины был ногами, согласно более недавней, перевернутой интерпретации. Один ряд ног – она балансировала, как будто на натянутом канате? Нет, новые ископаемые, обнаруженные в Китае, показывают второй ряд, и современная реконструкция выглядит так, как если бы она чувствовала себя как дома в реальном мире и была способна там выжить. Галлюцигения больше не классифицируется как «таинственное чудо» с неясными и, вероятно, давно исчезнувшими родственными связями. Вместо этого, вместе со многими другими кембрийскими ископаемыми, она теперь ориентировочно помещена в тип лопастеногих, у которого есть современные представители в форме перипатуса и других «онихофор» или «бархатных червей», встреченных нами на Свидании 26.

Могла чувствовать себя как дома в реальном мире. Ископаемая Hallucigenia fortis из Чэнцзян, Южный Китай.
Галлюцигения – современная реконструкция.
Во времена, когда кольчатые черви, как полагали, были близкими родственниками членистоногих, онихофоры часто навязывались как «промежуточное звено» – «заполнение пробела» между ними, хотя это не вполне полезное представление, если хорошо подумать о том, как действует эволюция. Кольчатые черви теперь помещены к спиральным, в то время как онихофоры, вместе с членистоногими, к линяющим. Перипатус с его древними родственными связями разумно помещен среди современных странников, чтобы поведать рассказ о Кембрийском взрыве.
Современные онихофоры широко распространены в тропиках, и особенно в Южном полушарии. Тот, что изображен ниже, Peripatopsis moseleyi – из Южной Африки. Все современные оникофоры живут на суше, в опавших листьях и влажных местах, где они охотятся на улиток, червей, насекомых и другую мелкую добычу. В кембрийском периоде, конечно, галлюцигения и ее отдаленные предки перипатус и перипатопсис жили, наряду со всеми другими, в море.
Связь галлюцигении с современными онихофорами все еще вызывает споры, и мы должны помнить, что большая творческая фантазия обязательно вмешивается между стертым и раздавленным ископаемым в камне и реконструкцией, которая, в конечном счете, нарисована, часто в смелом цвете, на странице. Было даже предположение, что галлюцигения вообще не могла быть отдельным животным, а была частью какого-то неизвестного животного. Это был бы не первый случай, когда была сделана такая ошибка. Реконструкции кембрийских сцен некоторых ранних художников включали плавающее, подобное медузе существо, по-видимому, навеянное консервированными кольцами ананаса, которые оказались частью челюстного аппарата таинственного хищного животного, аномалокариса (см. ниже). Другие кембрийские ископаемые, например Aysheaia, конечно, весьма похожи на морские версии перипатуса, и это укрепляет право перипатуса рассказать этот кембрийский рассказ.
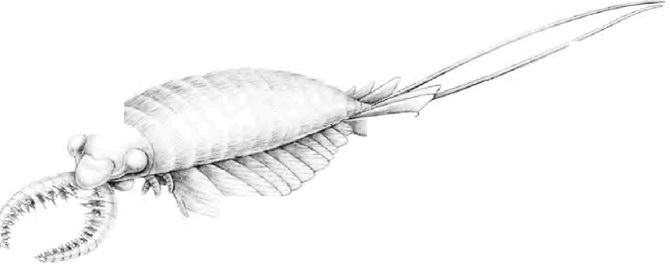
Вне зоологической сферы? Anomalocaris saron, из Чэнцзян.
Большинство окаменелостей из любой эры являются останками твердых частей животных: позвонков, панцирей членистоногих или раковин моллюсков или плеченогих. Но есть три кембрийских залежи окаменелостей – одна в Канаде, одна в Гренландии и одна в Китае – где причудливые условия, к большому счастью для нас, сохранили также и мягкие части. Это – сланец Бёрджесс в Британской Колумбии, Сириус Пассет на севере Гренландии, и район Чэнцзян в Южном Китае (Четвертый участок, Orsten («зловонный камень») в Швеции, сохраняет мягкие тела другим способом.). Сланец Бёрджесс был первоначально обнаружен в 1909 году и получил известность 80 лет спустя благодаря «Удивительной жизни» Стивена Гулда. Сириус Пассет, участок на севере Гренландии, был обнаружен в 1984 году, но пока менее изучен, чем два других. В том же году Hou Xianguang обнаружил окаменелости Чэнцзян. Доктор Hou – один из тех, кто совместно разрабатывал красиво иллюстрированную монографию «Кембрийские окаменелости Чэнцзян, Китай», изданную в 2004 году – к счастью для меня, непосредственно перед тем, как эта книга вышла в печать.
Окаменелости Чэнцзян теперь датированы 525 миллионами лет. Они почти ровесники с Сириус Пассет, и приблизительно на 10 или 15 миллионов лет старше, чем сланец Бёрджесс, но эти знаменитые залежи окаменелостей содержат похожую фауну. Это большое количество лопастеногих, многие более или менее похожи на морские версии перипатуса. Есть морские водоросли, губки, черви различных видов, плеченогие, похожие в значительной степени на современных, и загадочные животные неясных родственных связей. Есть большое количество членистоногих, включая ракообразных, трилобитов и многих других, отдаленно напоминающих ракообразных или трилобитов, но, возможно, принадлежащих к своим собственным, довольно обособленным группам. Большие (в некоторых случаях более метра), очевидно, хищные аномалокарисы и их ближайшие родственники найдены в Чэнцзян, так же как в сланце Бёрджесс. Никто полностью не уверен, кем они были – вероятно, дальними родственниками членистоногих – но они, должно быть, были эффектными. Не все «таинственные чудеса» сланца Бёрджесс были найдены в Чэнцзян, например опабиния, известная своими пятью глазами.
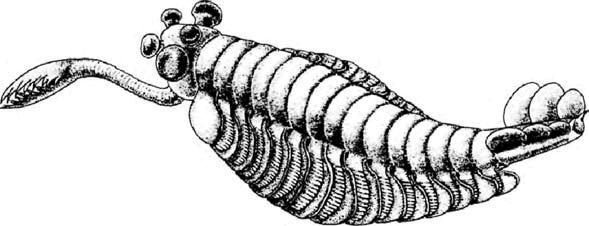
Известная пятью глазами Opabinia regalis, найденная в сланце Бёрджесс, Канада. Рисунок Мэрианн Коллинз.
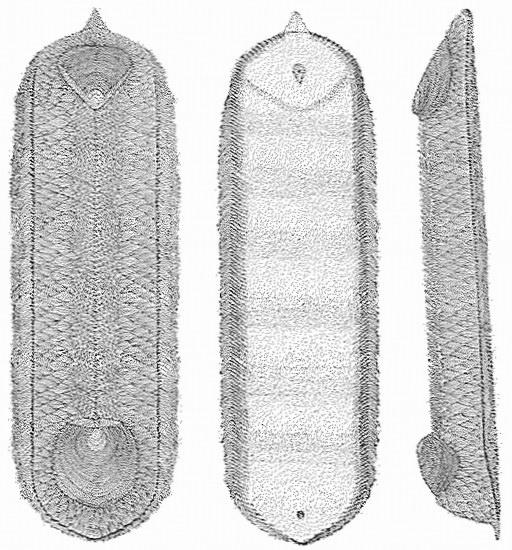
Разрушает мистическое благоговение перед большими типами
Halkieria evangelista из Сириус Пассет, Гренландия, датируемая нижним кембрием. Рисунок Саймона Конвея Морриса.
Фауна Сириус Пассет из Гренландии включает красиво названную халкиерию. Она, как считалось, была древним моллюском, но Саймон Конвей Моррис (Simon Conway Morris), который описал многих из странных существ кембрийского периода, полагает, что у нее есть сходство с тремя главными типами: моллюсками, плеченогими и кольчатыми червями. Это радует мое сердце, потому что помогает разрушить почти мистическое благоговение, с которым зоологи расценивают большие типы. Если мы относимся к своей эволюции серьезно, это должно иметь место, потому что, поскольку мы движемся в прошлое и приближаемся к пунктам свиданий с ними, типы станут все больше похожими друг на друга, все более близко связанными. Отвечает или нет халкиерия всем требованиям, волновало бы, если бы не было древнего животного, которое объединяет кольчатых червей, плеченогих и моллюсков. Обратите внимание на раковины, по одной на каждом конце, на иллюстрации выше.
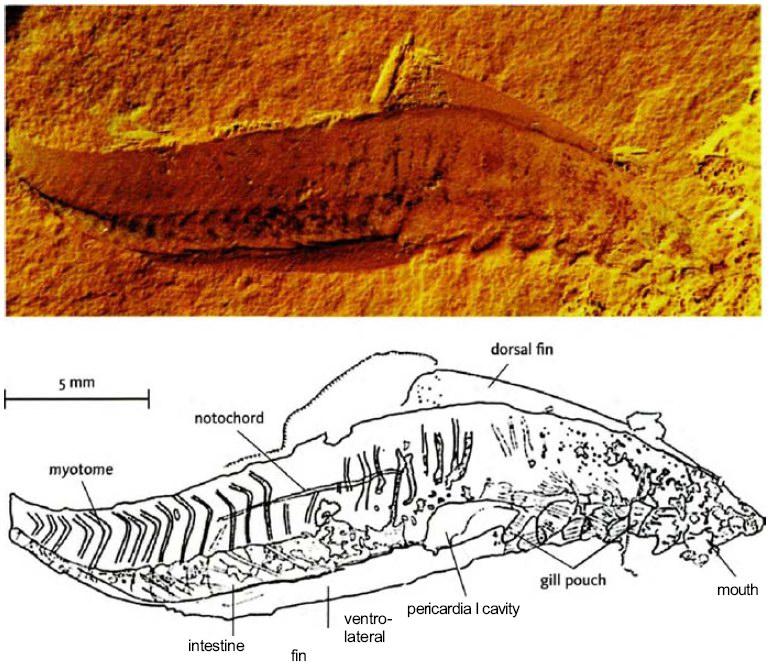
Позвоночные животные, как предполагалось, не были столь древними.
Ископаемая Myllokunmingia fengjiaoa, Чэнцзян. От D-G Shu и др. [264]
Как мы видели на Свидании 22, в Чэнцзян есть ископаемые, которые, кажется, являются настоящими позвоночными, предшествующими ланцетникоподобной пикайе из сланца Бёрджесс и другим кембрийским хордовым животным. Согласно традиционной зоологической мудрости, никогда не было позвоночных, возникших столь рано. Все же, миллокунмингия, больше чем 500 экземпляров которой были теперь обнаружены в Чэнцзян, в значительной степени похожа на хорошую бесчелюстную рыбу, возникшую, как ранее думали, 50 миллионов лет спустя, в середине ордовика. Сначала были описаны два новых рода – миллокунмингия, которая была охарактеризована как относительно близкая к миногам, и хайкоуихтис (Haikouichthys) (названный, увы, не в честь японской формы стиха), у которого, как полагали, было сходство с миксинами. Некоторые таксономисты-ревизионисты теперь помещают обоих в один вид, Myllokunmingia fengjiaoa. Это спорное обновление статуса хайкоуихтиса красноречиво говорит о том, как трудно различать детали очень древних ископаемых. Выше – фотография ископаемой особи миллокунмингии, вместе с рисунком, сделанным с помощью камеры-люциды. Я полон восхищения терпением, которое необходимо для реконструкции таких древних животных, как эти.
Смещение позвоночных назад, в середину кембрийского периода, только укрепляет идею о внезапном взрыве, которая является основой мифа. Действительно, похоже, что большинство нынешних главных типов животных сначала появляются как ископаемые в узком промежутке в кембрии. Это не означает, что не было никаких представителей этих типов до кембрийскиого периода. Но они, главным образом, не превращались в окаменелости. Как мы должны это истолковать? Мы можем выделить различные комбинации трех главных гипотез, скорее похожих на гипотезы взрыва млекопитающих после исчезновения динозавров.
Настоящего взрыва не было. Согласно этой точке зрения был только взрыв способности превращаться в окаменелость, а не подлинной эволюции. Типы, на самом деле, берут начало задолго до кембрийского периода, с копредками, разбросанными на сотни миллионов лет в докембрии. Это представление поддерживается некоторыми молекулярными биологами, которые использовали метод молекулярных часов для датирования основных копредков. Например, Г. Э. Рэй, Дж.С.Левинтон и Л.Х.Шапиро (G.A. Wray, J. S. Levinton, L. H. Shapiro) в известной статье в 1996 году оценили, что копредок, объединяющий позвоночных и иглокожих, жил приблизительно миллиард лет назад, а копредок, объединяющий позвоночных и моллюсков, еще на 200 миллионов лет раньше, более чем в два раза старше так называемого Кембрийского взрыва. Оценки молекулярных часов вообще имеют свойство отодвигать эти глубоко ветвящиеся пути назад в докембрий, намного дальше, чем это устраивает большинство палеонтологов. Согласно этой точке зрения, ископаемые по неизвестным причинам были не полностью сформированы до кембрия. Возможно, им не хватало годных к окаменению твердых частей, таких как раковины, панцири и кости. В конце концов, сланец Бёрджесс и залежи Чэнцзян крайне необычны среди всех геологических слоев, благодаря тому, что в них зарегистрированы окаменелые мягкие части. Возможно, докембрийские животные, несмотря на длительное существование в широком диапазоне сложно организованных тел, были просто слишком маленькими, чтобы превратиться в окаменелость. В пользу этой идеи есть некоторые маленькие типы животных, которые не оставили окаменелостей после кембрия вообще, пока не появились сегодня как живые «сироты». Почему, тогда, мы должны чувствовать себя вправе ожидать окаменелостей до кембрия? В любом случае, некоторые из найденных докембрийских ископаемых, включая фауну эдиакария и окаменелые отпечатки следов и нор, указывают на присутствие реального докембрийского многоклеточного.
Взрыв средней силы. Копредки, объединяющие различные типы, действительно жили довольно близко друг от друга по времени, но все еще были разбросаны более чем на несколько десятков миллионов лет до наблюдаемого взрыва окаменелостей. Далеко удаленный от современности Чэнцзян, возрастом 525 миллионов лет, на первый взгляд кажется довольно близким к предполагаемому копредку, которому, скажем, 590 миллионов лет. Но их разделяют целые 65 миллионов лет, такое же время, как прошло до сегодня со смерти динозавров – все время, в течение которого современные млекопитающие распространялись все шире, производя захватывающе разнообразные группы, которые мы видим сейчас. Даже 10 миллионов лет – длительное время, с учетом быстрых эволюционных взрывов «Рассказа Галапагосского Вьюрка» и «Рассказа Цихлиды». Слишком просто, глядя в прошлое, думать, что, поскольку мы признаем два древних ископаемых как принадлежащие различным современным типам, эти два ископаемых, должно быть, столь же отличаются друг от друга, как современные представители двух типов. Слишком легко забыть, что у современных представителей было пятьсот миллионов лет, чтобы разойтись. Нет никакой веской причины полагать, что кембрийский таксономист, счастливо лишенный богатства 500-миллионолетних зоологических знаний, поместил бы эти два ископаемых в отдельные типы. Он, возможно, разместил бы их только в отдельные отряды, несмотря на непостижимый в то время факт, что их потомкам было суждено, в конечном счете, разойтись насколько, что это послужит основанием для статуса отдельных типов.
Взрыв с коротким запалом. Это третья точка зрения, по моему мнению, безумная. Или, используя более парламентский язык, бесконтрольно и легкомысленно нереалистичная. Но я должен уделить ей некоторое время, потому что она недавно стала необъяснимо популярной, после риторики, которую я приписывал перегретым романтикой зоологам.
Согласно третьей точке зрения, новые типы возникли внезапно, в единственном скачке макромутации. Вот некоторые цитаты в остальном уважаемых ученых, которые я использовал прежде в «Расплетая радугу».
Как будто легкость, с которой делались эволюционные скачки, создавшие главные конструктивные новшества – основу новых типов – была почему-то утрачена, когда кембрийский период подошел к концу. Как будто главный двигатель эволюции потерял часть своей силы... В результате эволюция кембрийских организмов могла совершать большие скачки, включая скачки на уровне типов, в то время как позже она была более ограничена, делая только скромные скачки на уровне классов.
Или вот, тот же выдающийся ученый, о котором мы слышали в начале рассказа.
Вначале, в процессе ветвления, мы обнаруживаем множество длинных скачков мутаций, которые весьма значительно отличаются от основы и друг от друга. Эти виды имеют достаточно морфологических различий, чтобы быть классифицированными как основатели отдельных типов. Эти основатели также ветвятся, но делают это с помощью немного более близких вариантов длинных скачков, производя ветви от каждого основателя типа к различным дочерним видам, основателям классов. По мере продолжения процесса, обнаруживаются более пригодные варианты все более близкого соседства, так по очереди появляются основатели отрядов, семей и родов.
Эти цитаты побудили меня парировать, что это – как если бы садовник посмотрел на старый дуб и озадаченно заметил:
Не странно ли, что ни одна новая толстая ветвь не появились на этом дереве за много лет? В эти дни все новые побеги, кажется, на уровне тонких веточек!
Вот другая цитата, для которой, на сей раз, я укажу автора, потому что она была напечатана после «Расплетая Радугу», и я, поэтому, не использовал ее прежде. Эндрю Паркер (Andrew Parker) в «In the blink of an eye» главным образом увлечен защитой своей интересной и оригинальной теории, что Кембрийский взрыв был вызван внезапным приобретением животными глаз. Но прежде, чем подойти к своей теории непосредственно, Паркер начинает, принимая за чистую монету «бесконтрольную и легкомысленную» версию мифа о Кембрийском взрыве. Сначала он формулирует сам миф в наиболее откровенно «взрывной» версии, которую мне доводилось читать:
544 миллиона лет назад было действительно три типа животных с разнообразными внешними формами, но 538 миллионов лет назад их было тридцать восемь, такое же количество, которое существует сегодня.
Он продолжает пояснять, что не говорит о чрезвычайно быстрой градуалистической эволюции, сжатой в промежуток 6 миллионов лет, которая была бы крайней версией нашей второй гипотезы, и лишь с трудом приемлемой. И при этом он не говорит, как я, что вблизи изначального расхождения пары (будущих) типов они не могут быть совсем разными и должны действительно пройти через последовательные стадии, будучи парой видов, затем родов, и так далее до того, пока, в конечном счете, их разделение не послужило бы основанием признания их на уровне типов. Нет, Паркер расценивает каждый из своих появившихся за 538 миллионов лет 38 типов как полностью оперенный тип, который возник внезапно, в два макромутационных счета:
38 типов животных развилось на Земле. Таким образом, имели место только 38 монументальных генетических события, приведя к 38 различным внутренним организациям.
Монументальные генетические события не могут совершенно не обсуждаться. Контролирующие гены различных семейств Hox, которые мы встретили в «Рассказе Плодовой Мушки», могут, конечно, видоизменяться разительным образом. Но монументальное монументальному рознь. Плодовая мушка с парой ног на месте антенн – монументальнее не бывает, и даже тогда есть большой вопросительный знак относительно выживания. Для этого есть серьезная общая причина, которую я кратко объясню.
Для мутантного животного существует определенная вероятность стать лучше вследствие его новой мутации. «Лучше» означает по сравнению с родительским типом до мутации. Родитель должен быть, по крайней мере, настолько хорош, чтобы выжить и воспроизвести потомка, иначе это был бы не родитель. Легко заметить, что чем меньше мутация, тем больше вероятность, что она должна быть усовершенствованием. «Легко заметить» было любимой фразой великого статистика и биолога Р. Э. Фишера, и он иногда использовал ее, когда было совсем нелегко для обычных смертных заметить. В данном случае, однако, я думаю, что на самом деле легко следовать за аргументами Фишера в случае простой метрической особенности – например такой, как длина бедра, которая изменяется в одном измерении: некоторое количество миллиметров, которое могло стать большим или меньшими.
Представьте себе ряд мутаций увеличения размера. В одной крайности мутация нулевого размера, по определению – практически точная копия родительского гена, который, как мы видели, должен быть достаточно хорош, чтобы, по крайней мере, пережить детство и воспроизвести потомка. Теперь представьте себе случайную мутацию маленькой величины: нога, скажем, становится на один миллиметр длиннее или на один миллиметр короче. Полагая, что родительский ген не безупречен, у мутации, которая бесконечно мало отличается от родительской версии, есть 50-процентный шанс стать лучше и 50-процентный шанс стать хуже: будет лучше, если это будет шаг в правильном направлении, хуже, если это – шаг в противоположном направлении, относительно родительского положения. Но очень большая мутация, вероятно, будет хуже, чем родительская версия, даже если это будет шаг в правильном направлении, потому что она проскочит мимо. Чтобы дойти до крайности, вообразите нормального во всем человека с бедрами двухметровой длины.
Доводы Фишера имели более общий характер. Когда мы говорим о макромутационных скачках на территорию нового типа, мы больше не имеем дело с простыми метрическими признаками, такими как длина ног, и мы нуждаемся в другой версии доказательств. Существенным моментом, как я выразился прежде, является то, что существует гораздо больше способов быть мертвым, чем быть живым. Вообразите математический пейзаж всех возможных животных. Я должен назвать его математическим, потому что это – пейзаж в сотнях измерений, и он включает почти бесконечно большой диапазон мыслимых монстров, так же как (относительно) небольшое количество животных, которые действительно когда-либо жили. То, что Паркер называет «монументальным генетическим событием», было бы эквивалентно макромутации огромного действия, не только в одном измерении, как с нашим примером бедра, но в сотнях измерений одновременно. Таков масштаб изменений, о которых мы говорим, если, подобно Паркеру, вообразить резкое и немедленное превращение из одного типа в другой.
В многомерном пейзаже всех возможных животных живущие существа являются островками жизнеспособности, отделенными от других островов гигантскими океанами гротескного уродства. Начиная с любого острова, Вы можете эволюционировать далеко за его пределы, делая один шаг за раз, здесь медленно перемещаем ногу, там обрезаем кончик рога или закрашиваем перо. Эволюция – это траектория через многомерное пространство, в котором каждый шаг пути должен представлять собой тело, способное к выживанию и размножению, так же как родительский образец, достигнутый предыдущим шагом. Учитывая достаточное количество времени, довольно длинная траектория ведет от жизнеспособной отправной точки до жизнеспособного конечного пункта, настолько отдаленного, что мы признаем его как отдельный тип, скажем, моллюсков. А другая пошаговая траектория из одной и той же отправной точки может привести, через непрерывно жизнеспособные промежуточные звенья, к другому жизнеспособному конечному пункту, который мы признаем как уже другой тип, скажем, кольчатых червей. Нечто подобное, должно быть, случилось для каждой из вилок, приводящих к каждой паре типов животных от их соответствующего копредка.
Суть, к которой мы ведем, в следующем. Случайное изменение достаточной величины, чтобы породить новый тип одним махом, будет настолько большим в сотнях измерений одновременно, что нужно быть несообразно везучим, чтобы приземлиться на другом острове жизнеспособности. Почти неизбежно мегамутация такой величины приземлится в середине океана нежизнеспособности: вероятно, неопознаваемое как животное вообще.
Креационисты глупо уподобляют дарвиновский естественный отбор урагану, дующему через кладбище старых автомобилей, и которому посчастливилось собрать Боинг 747. Они, конечно, неправы, поскольку полностью упускают постепенную, кумулятивную природу естественного отбора. Но метафора кладбища старых автомобилей полностью подходит к гипотетическому неожиданному изобретению нового типа. Эволюционный шаг такой же величины, как, скажем, неожиданный переход от земляного червя к улитке, действительно должен был быть столь же везучим, как ураган на кладбище старых автомобилей.
Мы можем теперь с полной уверенностью отклонить третью из наших трех гипотез, безумную. Это оставляет другие две или некоторый компромисс между ними, и здесь я склонен быть агностиком, стремящийся к большему количеству данных. Как мы увидим в эпилоге к этому рассказу, кажется все более признанным, что ранние оценки молекулярных часов были преувеличены, когда отодвигали главные пункты ветвления на сотни миллионов лет назад в докембрий. С другой стороны, простой факт, что существует мало окаменелостей, если таковые вообще имеются, большинства типов животных до кембрийского периода, не должен заставлять нас предполагать, что эти типы, должно быть, эволюционировали чрезвычайно быстро. Аргументация урагана на кладбище старых автомобилей говорит нам, что у всех этих кембрийских ископаемых, по-видимому, были непрерывно эволюционировавшие предшественники. Эти предшественники должны были быть там, но они не были обнаружены. Какие бы ни были причины, и какой бы ни была шкала времени, они оказались не в состоянии фоссилизировать, но они должны были быть там. На первый взгляд, тяжелее поверить, что многие животные могли быть невидимыми в течение 100 миллионов лет, чем в течение всего лишь 10 миллионов лет. Это вынуждает некоторых людей предпочитать теорию кембрийского взрыва с коротким запалом. С другой стороны, чем короче Вы делаете запал, тем тяжелее поверить, что все это многообразие могло быть втиснуто в имеющееся в наличии время. Таким образом, этот довод урезает оба пути и не делает окончательного выбора между нашими двумя уцелевшими гипотезами.
Летопись окаменелостей не полностью лишена многоклеточной жизни перед Чэнцзян и Сириус Пассетом. Приблизительно на 20 миллионов лет ранее, почти ровно на кембрийско-докембрийской границе, начинает появляться множество микроскопических ископаемых, которые скорее похожи на крошечные раковины – вместе они известны как «маленькая раковинная фауна». Для большинства палеонтологов стало неожиданностью, когда некоторые из них были опознаны как бронированная обшивка лопастеногих – родственников бархатного червя. Это означает, что расхождение между различными группами первичноротых должно было произойти в докембрии, до видимого «взрыва».

Как мы должны это понимать? Dickinsonia costatt часть эдиакарской фауны.
И есть намеки на более древнее разнообразие животных. За двадцать миллионов лет до начала кембрийского периода, в эдиакарии позднего докембрия, отмечается всемирное процветание таинственной группы животных, названных эдиакарской фауной в честь Эдиакара Хиллз в Южной Австралии, где они были впервые найдены. Трудно сказать наверняка, чем являлось большинство из них, но они были среди первых окаменелых крупных животных. Некоторые из них – вероятно, губки. Некоторые немного похожи на медуз. Другие несколько напоминают актиний или морские перья (похожих на перья родственников актиний). Некоторые выглядят немного похожими на червей или слизняков и могли бы, очевидно, представлять настоящих двусторонне-симметричных животных. Другие же явно таинственны. Как мы должны понимать это существо, дикинсонию? Действительно ли это – коралл? Или червь? Или гриб? Или нечто совершенно отличающееся от всего того, что сохранилось сегодня? Есть даже одно похожее на головастика ископаемое из Австралии, все еще формально не описанное, которое подозревается в том, что оно – хордовое (тип, напомню, к которому принадлежат позвоночные). Если бы это оказалось правдой, это было бы очень волнующе, но поживем – увидим. Несмотря на такие дразнящие намеки, зоологи пришли к согласию в том, что эдиакарская фауна, хотя и интригующа, не очень помогает нам каким-то образом рассмотреть родословную большинства современных животных.
Есть также отпечатки окаменелостей, которые, кажется, являются следами или норами докембрийских животных. Эти следы говорят нам о раннем существовании ползающих животных, достаточно больших, чтобы их сделать. К сожалению, они не многое говорят нам о том, на что были похожи эти животные. Есть также некоторые еще более древние, главным образом микроскопические окаменелости, найденные в Доушаньто, в Китае, которые представляются эмбрионами, хотя не ясно, в какое животное они, возможно, превратились. Еще более древние являются маленькими, дискообразными отпечатками из Северо-Западной Канады, датированными приблизительно между 600 и 610 миллионами лет назад, но эти животные, пожалуй, еще более загадочны, чем эдиакарские формы.
Эта книга опирается на ряд из 39 пунктов свиданий, и хотелось бы высказать своего рода предположения относительно возраста каждого. Большинство пунктов свиданий может теперь быть датировано с определенной уверенностью, используя сочетание поддающихся датировке окаменелостей и молекулярных часов, калиброванных такими окаменелостями. Не удивительно, что окаменелости начинают подводить нас, когда мы достигаем более древних пунктов свиданий. Это означает, что молекулярные методы больше не могут быть надежно калиброваны, и мы входим в дебри недатируемости. Для полноты я заставил себя проставить своего рода даты для этих первобытных копредков, примерно от Копредка 23 до 39. Самое последнее доступное свидетельство, мне кажется, поддерживает, даже если только слегка, точку зрения, близкую к взрыву с запалом средней длины. Это противоречит моему более раннему убеждению в пользу отсутствия реального взрыва. При поступлении дополнительных данных, что, надеюсь, будет происходить, я не буду сколь-нибудь удивлен, если мы продвинемся другим путем дальше в глубокий докембрий в наших поисках копредков современных типов животных. Или мы можем быть отодвинуты назад к впечатляюще короткому взрыву, в котором копредки большинства больших типов животных уплотнены во временном отрезке 20 или даже 10 миллионов лет около начала кембрия. В этом случае я серьезно ожидаю, что, даже если мы правильно помещаем двух кембрийских животных в различные типы на основании их подобия современным животным, ранее в кембрийском периоде они были бы намного ближе друг к другу, чем современные потомки каждого к современным потомкам другого. Кембрийские зоологи не поместили бы их в отдельные типы, а только, скажем, в отдельные подклассы.
Я не удивился бы, увидев любую из первых двух гипотез доказанной. Я не принимаю риск на себя. Но я съем свою шляпу, если будет когда-либо найдено какое-нибудь доказательство в пользу третьей гипотезы. Есть все причины полагать, что эволюция в кембрийском периоде была по существу таким же процессом, как эволюция сегодня. Вся эта взволнованная сверх меры риторика о главной движущей силе эволюции, ослабленной после кембрия, все эти эйфорические возгласы о диких, беззаботных танцах сумасбродных изобретений, с новыми типами, прыгнувшими в жизнь во время блаженного рассвета зоологической легкомысленности – из-за всего этого я готов принять риск на себя: все это просто явная глупость.
Я спешу сказать, что не имею ничего против поэзии и прозы кембрия. Но дайте мне вариант Ричарда Форти (Richard Fortey) на странице 120 его красивой книги «Life: an unauthorised biography»:
Я могу представить себя стоящим на кембрийском берегу вечером, почти так же, как я стоял на берегу архипелага Шпицберген и впервые задавался вопросом о биографии жизни. Морской прибой у моих ног выглядел и ощущался бы почти так же. Там, где море встречается с сушей, есть полоса немного липких, скругленных строматолитных подушек, оставшихся от обширных рощ докембрия. Ветер со свистом проносится через красные равнины позади меня, где нет никакой видимой жизни, и я могу почувствовать острые уколы переносимого ветром песка на задней стороне моих ног. Но в грязном песке у моих ног я могу увидеть отпечатки червя, небольшие извивающиеся бороздки, выглядящие так знакомо. Я могу увидеть следы отпечатков, оставленных скрывшимися ракообразными животными... Кроме свиста бриза и ударов прибоя, совсем тихо, и никто не кричит среди ветра...
Эпилог к «Рассказу Бархатного Червя»
В большей части этой книги я беззаботно разбрасывался датами свиданий, и даже был достаточно опрометчив, представляя многих копредков, присваивая им определенное число «пра-» перед «прародителем». Мои даты главным образом были основаны на окаменелостях, которые, как мы увидим в «Рассказе Секвойи», могут быть датированы с точностью, соизмеримой с широко используемой шкалой времени. Но окаменелости никогда сильно не помогали нам при рассмотрении родословной мягкотелых животных, таких как бархатный червь. Целаканты пропали из летописи в течение прошлых 70 миллионов лет, вот почему открытие живого целаканта в 1938 году было таким волнующим сюрпризом. Летопись окаменелостей даже в лучшие времена может быть ненадежным свидетельством. И теперь, достигнув кембрийского периода, мы, к сожалению, остаемся без окаменелостей. Независимо от того, как мы интерпретируем «взрыв», все сходятся во мнениях, что почти все предшественники большой кембрийской фауны по неясным причинам были не в состоянии фоссилизировать. Поскольку мы ищем копредков, предшествующих кембрию, мы не находим больше помощи в горных породах. К счастью, окаменелости – не единственная наша помощь. В «Рассказе Слоновой Птицы», «Рассказе Розогуба» и других местах мы использовали изобретательную технику, известную как молекулярные часы. Настало время объяснить ее должным образом.
Разве не было бы замечательно, если бы измеряемые или исчисляемые эволюционные изменения случались с постоянной скоростью? Мы могли бы тогда использовать саму эволюцию как ее собственные часы. И это не должно приводить к логическому кругу, потому что мы могли бы калибровать эволюционные часы на участках эволюции, где летопись окаменелостей оправданна, а затем экстраполировать на участки, где она не годится. Но как мы измерим скорость эволюции? И, даже если мы могли бы ее измерить, с какой стати мы должны ожидать, что какой-нибудь аспект эволюционных изменений должен идти с постоянной скоростью, как часы?
Нет ни малейшей надежды, что длина ног, или размер мозга, или число усов будут эволюционировать с постоянной скоростью. Такие особенности важны для выживания, и их скорости эволюции, конечно, будут ужасно непостоянны. Как часы, они обречены самими принципами своей собственной эволюции. В любом случае, трудно вообразить согласованный стандарт для измерения скорости видимой эволюции. Вы измеряете эволюцию длины ноги в миллиметрах за миллион лет как процентное изменение за миллион лет, или как? Дж. Б. С. Холдейн (J. B. S. Haldane) предложил единицу скорости эволюции, дарвин, который основан на пропорциональном изменении за поколение. Везде, где он использовался на реальных ископаемых, результаты варьируют от миллидарвинов до килодарвинов и мегадарвинов, и никто не удивлен.
Молекулярные изменения похожи на намного более обнадеживающие часы. Во-первых, потому что ясно, что Вы должны измерить. Так как ДНК является текстовой информацией, написанной в четырехбуквенном алфавите, есть вполне естественный способ измерить скорость ее эволюции. Вы только подсчитайте буквенные различия. Или, если хотите, Вы можете прибегнуть к помощи белковых продуктов кода ДНК и подсчитать замены аминокислот (Когда молекулярные часы были впервые предложены Эмилем Цукеркандлем (Emile Zuckerkandl) и великим Лайнусом Полингом (Linus Pauling), это было единственным доступным методом.). Есть причины надеяться, что большинство эволюционных изменений на молекулярном уровне являются нейтральными, а не управляются естественным отбором. Нейтральные – не то же самое, что бесполезные или нефункциональные – это лишь означает, что различные версии гена одинаково хороши, поэтому изменение от одного к другому не замечается естественным отбором. Это хорошо для часов.
Вопреки моей довольно смешной репутации «ультрадарвиниста» (клевета, против которой я выступил бы более решительно, если бы название не звучало для меня столь лестно), я не думаю, что большинство эволюционных изменений на молекулярном уровне было одобрено естественным отбором. Напротив, я всегда уделял много времени так называемой нейтральной теории, связанной с великим японским генетиком Мотоо Кимурой (Motoo Kimura), или ее продолжению, «почти нейтральной» теории его сотрудницы Томоко Охты (Tomoko Ohta). Реальный мир, конечно, не интересуют пристрастия человека, но оказалось, что я, безусловно, хотел бы, чтобы такие теории были верными. Потому, что они дают нам отдельную, независимую хронику эволюции, не связанную с видимыми особенностями окружающих нас существ, и они дают надежду, что нечто вроде молекулярных часов могло бы действительно работать.
На случай, если смысл был неправильно понят, я должен подчеркнуть, что нейтральная теория ни в коем случае не преуменьшает важность отбора в природе. Естественный отбор всесилен относительно тех видимых изменений, которые затрагивают выживание и размножение. Естественный отбор – единственное объяснение, которое мы знаем для функциональной красоты и будто бы «спланированной» сложности живых существ. Но если есть какие-нибудь изменения, которые не имеют никакого видимого результата – изменения, которые проходят незамеченными прямо перед радаром естественного отбора – они могут накапливаться в генофонде безнаказанно и могут дать именно то, что нам нужно для эволюционных часов.
Как всегда, Чарльз Дарвин опередил свое время в отношении нейтральных вариаций. В первом издании «Происхождения видов», почти в начале 4 главы, он написал:
Сохранение благоприятных вариаций и отклонение вредных вариаций я назвал естественным отбором. Вариации бесполезные и безвредные не подвергаются действию естественного отбора; они сохраняются как колеблющийся элемент, как это наблюдается у некоторых видов, названных полиморфными.
В шестом и последнем издании во втором предложении было даже больше современно звучащих дополнений:
...как это наблюдается у некоторых полиморфных видов, либо же, в конце концов, становятся фиксированными...
«Фиксированный» – генетический специальный термин, и Дарвин, конечно, не мог иметь его в виду в полностью современном смысле, но он дает мне прекрасное вступление к следующему вопросу. Новая мутация, частота которой в популяции вначале близка к нулю по определению, как говорят, становится «фиксированной», когда она достигает 100 процентов в популяции. Скорость эволюции, которую мы стремимся измерить для молекулярных часов, является скоростью, с которой последовательность мутаций в одном и том же генетическом локусе фиксируется в популяции. Оказалось, что очевидный путь для фиксации – если естественный отбор одобряет новую мутацию относительно предыдущего аллеля «дикого типа», и поэтому ведет к его закреплению: он становится нормой, критерием ритма часов. Но новая мутация может также закрепиться, даже если она ровно настолько же хороша, как ее предшественница – истинная нейтральность. Отбор ничего не делает: все происходит абсолютно случайно. Вы можете моделировать процесс, бросая монету, и можете вычислить скорость, с которой это случится. Как только нейтральная мутация сдрейфует к 100 процентам, она станет нормой, так называемым «диким типом» в этом локусе, пока другой мутации не посчастливится сдрейфовать к фиксации.
Если есть сильный компонент нейтральности, у нас потенциально могли бы быть изумительные часы. Сам Кимура не был очень увлечен идеей молекулярных часов. Но он верил – теперь это кажется справедливым – что большинство мутаций в ДНК действительно являются нейтральными – «ни полезными, ни вредными». И, в замечательно ясном и простом алгебраическом примере, который я не буду здесь подробно выписывать, он вычислил, что, если это верно, то скорость, с которой подлинно нейтральные гены должны «в конечном счете, стать фиксированными», точно равна скорости, с которой вариации образуются изначально: скорости мутации.
Вы видите, насколько прекрасно это для каждого, кто хочет датировать пункты развилок («свидания»), используя молекулярные часы. Пока скорость мутации в нейтральном генетическом локусе остается постоянной в течение долгого времени, скорость фиксации также будет постоянной. Вы можете теперь сравнить один и тот же ген у двух различных животных, скажем, ящера и морской звезды, последним общим предком которых был Копредок 25. Теперь подсчитайте число букв, в которых ген морской звезды отличается от гена ящера. Предположим, что половина различий накопилась в линии, ведущей от копредка к морской звезде, а другая половина в линии, ведущей от копредка к ящеру. Это дает Вам количество тиканий часов, начиная со Свидания 25.
Но все не так просто, и осложнения интересны. Во-первых, если бы Вы слушали тиканье молекулярных часов, то оно не было бы регулярным, как у маятниковых или наручных пружинных часов; оно было бы похоже на счетчик Гейгера возле источника радиации. Совершенно беспорядочное! Каждое тиканье – фиксация еще одной мутации. Согласно нейтральной теории, интервал между следующими друг за другом тиканиями может быть длинным, или он может быть коротким, случайным «генетическим дрейфом». В счетчике Гейгера время следующего тиканья непредсказуемо. Но – и это действительно важно – средний интервал в большом количестве тиканий весьма предсказуем. Надежда на то, что молекулярные часы предсказуемы, так же как счетчик Гейгера, и как правило, это верно.
Во-вторых, скорость тиканья варьирует от гена к гену в геноме. Это было замечено давно, когда генетики могли видеть только белковые продукты ДНК, а не саму ДНК. Цитохром-С эволюционировал со своей собственной характерной скоростью, которая быстрее, чем у гистонов, но медленнее, чем у глобинов, у которых в свою очередь медленнее, чем у фибринопептидов. Таким же образом, когда счетчик Гейгера подвергается воздействию очень слабого источника радиации, такого как глыба гранита, в сравнении с очень сильным радиоактивным источником, таким как глыба радия, время следующего тиканья всегда непредсказуемо, но средняя скорость тиканья предсказуема и кардинально различна, если Вы двигаетесь от гранита к радию. Гистоны похожи на гранит, тикающий с очень медленной скоростью; фибринопептиды похожи на радий, жужжащий как случайная пчела. Другие белки, такие как цитохром-С (или, скорее, гены, которые их создают) являются промежуточными. Существует спектр генных часов, каждые идут со своей собственной скоростью, и каждые полезны для различных целей датирования и для перекрестного контроля друг друга.
Почему различные гены бегут с различными скоростями? Что отличает «гранитные» гены от «радиевых» генов? Вспомните, что нейтральные не означает бесполезные, это означает одинаково хорошие. Гранитные гены и гены радия – оба полезны. Просто радиевые гены могут изменяться во многих местах на своем протяжении и все еще быть полезными. Из-за способа работы гена, участки на его протяжении могут изменяться безнаказанно, не затрагивая его функций. Другие части того же гена очень чувствительны к мутации, и его функции нарушаются, если эти части поражены мутацией. Возможно, у всех генов есть гранитная часть, которая не должна сильно изменяться, если ген должен продолжать работать, и радиевые части, которые могут переключаться беспрепятственно, пока гранитная часть не затронута. Возможно, у гена цитохрома-С есть смесь гранитных и радиевых кусочков; у генов фибринопептида – более высокая доля радиевых кусочков, в то время как у генов гистона – более высокая доля гранитных кусочков. Есть некоторые проблемы, или, по крайней мере, затруднения с тем, как объяснить различия в скорости тиканья между генами. Но вся суть для нас в том, что скорости тиканья действительно варьируют от гена к гену, в то время как скорость для любого конкретного гена является довольно постоянной даже у очень отдаленных видов.
Однако не совсем постоянной, и это приносит нам следующую проблему, которая является серьезной. Скорости тиканья не просто неопределенны и нестабильны. Для любого конкретного гена они могут быть систематически больше у некоторых видов существ, чем у других, и это вносит реальную погрешность. У бактерий намного менее эффективная система репарации ДНК, чем сложное «исправление ошибок» нашей ДНК, таким образом, их гены видоизменяются с более высокой скоростью, и их молекулярные часы тикают быстрее. У грызунов также немного неряшливые репаративные ферменты, чем объясняется, почему молекулярная эволюция быстрее у грызунов, чем у других млекопитающих. Главные изменения в эволюции, как внесенные «сгоряча», имеют возможность изменять скорость мутации, что могло бы причинить вред нашими оценками дат ветвления. Сейчас развиваются сложные методы, которые могут учесть изменяющиеся скорости мутации в различных линиях, но они находятся в стадии становления.
Еще более тревожно, что время размножения, казалось бы, предполагает максимальную возможность для мутации. Таким образом, виды с короткими циклами жизни, такие как плодовые мушки, ускоряют мутации до более высокой нормы за миллион лет, чем, скажем, слоны с их длинными интервалами между поколениями. Это предполагало бы, что молекулярные часы могли бы быть рассчитанными в поколениях, а не в реальном времени. Фактически, однако, когда молекулярные биологи изучили скорости изменений в последовательностях, используя линии, у которых оказались хорошие летописи окаменелостей для калибровки, они этого не обнаружили. Казалось, действительно молекулярные часы измеряли время в годах, а не в поколениях. Это было мило, но как это объяснить?
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Рассказ Евы
Рассказ Евы Есть впечатляющая разница между «родословными деревьями генов» и «родословными деревьями людей». В отличие от человека, который происходит от двух родителей, у гена есть только один родитель. Каждый из Ваших генов должен был быть получен или от Вашей матери,
Рассказ Секвойи
Рассказ Секвойи Люди спорят о том, какое одно место в мире Вы должны посетить прежде, чем умрете. Мой кандидат – лес Muir Woods, несколько севернее моста «Золотые Ворота». Или, если Вы считаете, что слишком поздно, я не могу вообразить лучшего места, чтобы быть похороненным (вот
Рассказ Taq
Рассказ Taq Достигнув нашего самого древнего свидания, собрав в нашем странствии всю жизнь, которую знаем, мы имеем возможность рассмотреть ее разнообразие. На самом глубинном уровне разнообразие жизни является химическим. Профессии, которыми заняты наши
Рассказ Stw 573
Рассказ Stw 573 Не думаю, что есть смысл придумывать причины, в силу которых ходить на двух ногах – это здорово. Будь так, шимпанзе делали бы то же самое, не говоря уже о других животных. Нет причины, по которой бег на двух или четырех конечностях быстрее или удобнее.
Рассказ Гориллы
Рассказ Гориллы Становление дарвинизма, которое пришлось на XIX век, привело к появлению двух противоположных взглядов на человекообразных обезьян. Противники Дарвина, хотя и согласившиеся принять идею эволюции, были в ужасе от возможного родства с грубыми,
Рассказ Орангутана
Рассказ Орангутана Возможно, заявление о наших давних связях с Африкой было поспешным. Что если наши предки покинули Африку около 20 млн лет назад и поселились в Азии, а 10 млн лет назад вернулись в Африку?Если так, то современные человекообразные обезьяны, включая тех,
Рассказ Гиббона[12]
Рассказ Гиббона[12] На рандеву № 4 мы встречаем крупную группу пилигримов. И теперь могут возникнуть проблемы с установлением родства. (Чем дальше, тем затруднительнее это сделать.) Существует двенадцать видов гиббонов, принадлежащих к четырем основным группам. Это Bunopithecus
Рассказ Айе-айе
Рассказ Айе-айе Один британский политик как-то описал конкурента (который позднее стал лидером партии) как человека, “в котором есть что-то ночное”. При взгляде на айе-айе складывается именно такое впечатление, и это не случайно: она ведет целиком ночной образ жизни. Из
Рассказ Шерстокрыла
Рассказ Шерстокрыла Шерстокрыл из Юго-Восточной Азии мог бы рассказать нам, каково это – парить во воздуху ночного леса. Но для нас, пилигримов, у него приготовлена гораздо более приземленная история. Мораль ее в том, что составленная нами схема с сопредками, рандеву и
Рассказ Мыши
Рассказ Мыши Из тысяч грызунов домашняя мышь (Mus musculus) может поведать кое-что особенное: ни один другой вид млекопитающих, кроме нашего собственного, мы не изучали так интенсивно. Именно на мышах, а не на пресловутых морских свинках держатся медицинские, физиологические и
Рассказ Бобра
Рассказ Бобра “Фенотип” – это то, что находится под влиянием генов. В целом под фенотипом понимают все, что относится к телу. Но есть тонкости, вытекающие из этимологии. Phamo по-гречески означает “показывать”, “обнаруживать”, “выявлять”, “выражать”, “раскрывать”.
Рассказ Гиппопотама
Рассказ Гиппопотама В школе, изучая греческий, я узнал, что “гиппос” означает “лошадь”, а “потамос” – “река”. Выходило, что гиппопотамы – “речные лошади”. Позднее, забросив греческий и занявшись зоологией, я узнал, что гиппопотамы не имеют отношения к лошадям. В
Рассказ Тюленя
Рассказ Тюленя В большинстве естественных популяций число самцов и самок примерно одинаково. На то есть эволюционные причины, и о них рассказал замечательный специалист по статистике и эволюционной генетике Рональд Э. Фишер. Представьте себе популяцию, количество в
Рассказ Броненосца
Рассказ Броненосца С зоологической точки зрения Южная Америка сродни Мадагаскару: как и Мадагаскар, она откололась от Африки – но не с восточной, а с западной стороны. Произошло это примерно тогда же или чуть позднее. Как и Мадагаскар, в течение почти всего времени
Рассказ Утконоса
Рассказ Утконоса Старое латинское название утконоса – Ornithorhynchus paradoxus [“парадоксальный птицеклюв”]. Когда его обнаружили, он показался ученым настолько нелепым, что присланный в музей экземпляр сочли чучелом из сшитых вместе частей тела млекопитающего и птицы. Кое-кто
Рассказ Taq
Рассказ Taq Итак, мы встретили практически все существующие формы жизни. Теперь можно окинуть взглядом открывшееся разнообразие. На самом глубоком уровне разнообразие жизни является химическим. Профессии, которыми владеют наши пилигримы, охватывают широкий диапазон