Регуляция размножения и дифференцировки кроветворных клеток
Регуляция размножения и дифференцировки кроветворных клеток
В связи с успехами исследования процессов кроветворения и функциональных возможностей стволовых клеток при использовании метода селезеночных колоний и культур на полутвердых средах и других методов представления о регуляции путей их пролиферации и дифференцировки существенно изменились.
Экспериментально подтвердилось существование единой полипотентной стволовой клетки, способной к дифференцировкам по всем направлениям гемопоэза, включая лимфопоэз (рис. 3). Казалось бы, это непосредственно доказывает правильность исходного положения о конкуренции различных ростков кроветворения за направление дифференцировки стволовой клетки в соответствии с потребностями организма. Однако вопрос оказался значительно сложнее.
Наряду с исходной полипотентной стволовой кроветворной клеткой могут существовать и обладающие стволовыми свойствами предшественники, коммитированные (т. е. ограниченные к выбору направления дифференцировки) предшественники двух типов — миелопоэза и лимфопоэза. Это первое и главное разделение направлений возможной дифференцировки единой полипотентной стволовой клетки. Как осуществляется регуляция выбора дифференцировки — в сторону миелопоэза или лимфопоэза, не очень ясно.
Стволовые клетки, коммитированные в сторону лимфопоэза, дифференцируются сначала в общих, а затем в раздельных предшественников Т- и В-лимфоцитов, из которых через ряд промежуточных стадий образуются Т-лимфоциты (приобретая определенные клеточные рецепторы) и В-лимфоциты (приобретая рецепторы к комплементу, к фрагментам и комплексам иммуноглобулинов и комплексам антиген—антитело). Кроме того, из них образуются плазматические клетки. Лимфопоэз наименее изучен. До сих пор неясно, имеются ли в лимфоцитарном ряду клетки, не способные к пролиферации. Среди лимфоидных клеток трудно определить принадлежность их к пролиферирующему, созревающему или функциональному пулам, так как возможны взаимные перекрытия.
Стволовые клетки, коммитированные в сторону миелопоэза, в дальнейшем через поэтинчувствительные предшественники разделяются на три ростка: эритроцитарный, гранулоцито-макрофагальный и тромбоцитарный.
В эритроцитарном направлении выявлено несколько промежуточных клеток (в том числе и бурстообразующая клетка) между общим предшественником миелопоэза и эритропоэтинчувствительными клетками. Последующие морфологически различаемые клетки относятся к проэритробласту, трем видам эритробластов, ретикулоциту и зрелому эритроциту. Обычно происходит 5—6 митозов в морфологически различимых клетках, и каждый эритробласт дает 30—60 эритроцитов. Эритробласты теряют способность к делению, как только в клетке накопится 27 нг гемоглобина. В случае замедления синтеза гемоглобина увеличивается число делений эритробластов. Из одной полипотентной стволовой клетки, начавшей дифференцировку в сторону миелопоэза, может образоваться около 1 млн эритропоэтинчувствительных клеток и около 0,1 млн клеток — предшественников гранулоцитов и макрофагов.
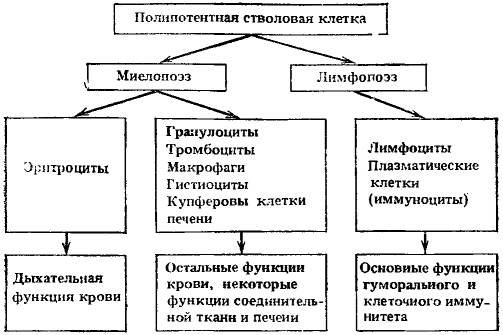
Рис. 3. Основные пути дифференцировки полипотентной стволовой клетки, клеточные формы и их главные функции
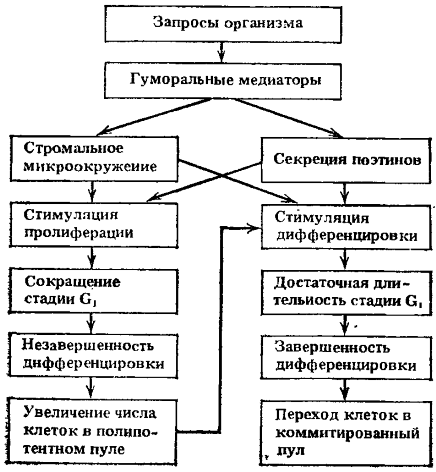
Рис. 4. Возможные пути регуляции пролиферации и дифференцировки полипотентных стволовых клеток в соответствии с запросами организма
Развитие гранулоцито-макрофагального направления миелопоэза проходит на стадии образования клеток, дающих колонии на полутвердых средах (KOEK), кластерообразующих клеток и предшественников гранулоцитов и моноцитов-макрофагов. Последующие клетки морфологически различимы. Миелобласт нейтрофильный далее дает клетки гранулоцитарного ряда (эезинофил), нейтрофильного ряда (нейтрофил) и базофильного ряда (базофил и тучная клетка).
Монобласт дает моноциты, макрофаги, гистиоциты, купферовы клетки печени и остеокласты. Клетки моноцитопоэтического ряда объединены в систему фагоцитирующих мононуклеаров как способные к фагоцитозу и осуществляющие в норме функции клеточного фагоцитоза. Ранее входившие вместе с указанными клетками в ретикуло-эндотелиальную систему ретикулярные клетки и эндотелиальные клетки исключены из системы фагоцитирующих мононуклеаров как являющиеся лишь факультативными фагоцитами, не имеющими рецепторов для иммуноглобулинов и не способными к иммунному фагоцитозу. Клетки же системы фагоцитирующих мононуклеаров в процессе дифференцировки приобретают рецепторы для иммуноглобулинов и комплемента и становятся способными к активному иммунному фагоцитозу.
Тромбопоэтинчувствительные клетки формируют морфологически различимые клетки: мегакариобласт, промегакариоцит и тромбоциты. Для предшественников характерны 4—5 эндомитозов.
Таким образом, по современным данным, клетки костного мозга, способные к делению, можно условно разделить на морфологически различимые клетки, как правило способные к очень ограниченному числу делений (4—8 делений), и на морфологически неразличимые клетки, выявляемые лишь по способности их давать колонии в селезенке облученных мышей или пролиферировать на полутвердых средах (и другими методами), способные к чрезвычайно активной пролиферации. Число делений, которые может совершить полипотентная стволовая клетка, на порядок больше, чем для морфологически различимых клеток. В норме их в костном мозге лишь около 0,5% от числа всех клеток. Однако достаточно сохраниться 0,1% стволовых клеток после сильного радиационного воздействия, чтобы обеспечить восстановление нормального пула стволовых клеток и резкое повышение дифференцировки клеток в последующих отделах.
При нормальном кроветворении абсолютное большинство стволовых клеток находится вне клеточного цикла (период G0 или длительный период G1). Стабильное поддержание величины отдела стволовых клеток обеспечивается небольшой долей клеток, вступивших в митотический цикл. В случае уменьшения отдела стволовых клеток в пролиферацию может вовлекаться вся популяция сохранившихся стволовых клеток. При этом время удвоения популяции стволовых клеток оказывается очень коротким, порядка 15—20 ч. По данным ряда авторов, укорочение или удлинение стадии G1 стволовых клеток соответственно сопровождается пролиферацией (т. е. оставлением в пуле стволовых клеток) или дифференцировкой (т. е. уходом в последующие отделы).
Следовательно, основной вклад в восполнение дефицита клеточности костного мозга вносит пролиферация морфологически неразличимых клеток и потому исследование регуляции их численности и регуляции выбора направления дифференцировки является чрезвычайно важным. К сожалению, в этих вопросах еще очень много неясного.
Как решается для стволовых клеток первый выбор направления дифференцировки (в сторону лимфопоэза или миелопоэза), тоже неясно. По многочисленным данным, в опытах на собаках и обезьянах аллогенная кроветворная ткань вызывала появление огромного количества лимфоцитов, специфически иммунных против реципиента, при полной или частичной блокаде остального кроветворения. Следовательно, возможно переключение части стволовых клеток с эритроидного на лимфоидный путь дифференцировки. Характер дифференцировки кроветворной ткани в условиях трансплантации указывает на конкуренцию миелоидных и лимфоидных клеток за общий предшественник. У полицитемических радиохимер, у которых снижен эритропоэз, ускоряется восстановление лимфопоэза. Приведенные нами данные о существенной задержке пострадиационного восстановления числа лимфоцитов в крови человека в период, когда эритропоэз напряжен, также свидетельствуют о конкурентных отношениях ростков кроветворения. Об этом непосредственно говорит и проведенное нами изучение соотношений в морфологически различаемых клетках разных ростков костного мозга у облучавшихся собак.
Тем не менее некоторыми учеными предполагается, что направление дифференцировки стволовых клеток или закреплено генетически постоянно, или осуществляется стохастически, в обоих случаях — независимо от запросов организма. Возможным доказательством этого может служить одно и то же распределение маркерных клеток по разным росткам гемопоэза при изменении общих запросов организма.
В связи с изложенным, вероятно, более правильно считать существование конкурентных отношений между ростками кроветворения за преимущественную продукцию конечных зрелых форм клеток, а не примат дифференцировки полипотентной стволовой клетки в тот или иной росток гемопоэза. При этом влияние дальнодействующей для гемопоэза регуляции с целью реакций на запросы организма в экстремальных ситуациях может осуществляться не только через поэтинподобные регуляторы, но и через микроокружение в локальных участках гемопоэтических тканей, которое может зависеть также от запросов организма. Эритропоэтин для красного ростка и колониестимулирующий фактор для гранулоцитарного ростка стимулируют пролиферацию клеток и укорачивают средний период генерации клеток. При этом удлинение или укорочение стадии G1 клеточного цикла может определять уход клеток в пролиферацию или дифференцировку (рис. 4). В случае вступления клетки в деление сокращается генерационный цикл и клетка не успевает осуществить дифференцировку. Пролиферация прекращается, как только весь объем микроокружения окажется занятым стволовыми клетками.
Таким образом, исследования реакции отдела стволовых клеток на экстремальные воздействия и пострадиационные изменения гемопоэза позволили показать ряд общих закономерностей в реакции этой системы на экстремальные воздействия и в процессах последующего восстановления.
Влияние запросов организма на гемопоэз осуществляется дальнодействующей регуляцией посредством гуморальных факторов. Поэтому следует кратко рассмотреть отдельные радиационные изменения со стороны эндокринной и других систем организма.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
1. Стабильность дифференцировки и эпигенетическая наследственность
1. Стабильность дифференцировки и эпигенетическая наследственность Факторы, вызывающие дифференцировку, обычно действуют короткое время, в то время как само дифференцированное состояние может сохраняться гораздо дольше. Например, нервная ткань у позвоночных
5. Программы размножения и гибели клеток
5. Программы размножения и гибели клеток По мере развития количество клеток, из которых состоит зародыш, увеличивается. Деления клеток (дробление яйца) на самых ранних стадиях развития происходят равномерно (синхронно). Ho у одних видов раньше, у других позже эта
4. Формы размножения организмов
4. Формы размножения организмов Преемственность поколений организмов в природе осуществляется за счет воспроизведения. Размножение — это способность организма воспроизводить себе подобных. В природе существует два типа размножения: бесполое и половое.Виды бесполого
8.7. Регуляция процессов размножения и полового поведения
8.7. Регуляция процессов размножения и полового поведения Большой сложностью отличается нейрогуморальная регуляция процессов размножения и полового поведения. Роль гипоталамо-гипофизарной системы в этой регуляции особенно наглядна. Гипоталамус, посредством секреции
Долгожительство зависит от размера, размножения и еды
Долгожительство зависит от размера, размножения и еды В последнее время известный берлинский профессор Рубнер сделал попытку определить количество энергии, потребляемой во время роста и в продолжение жизни, думая найти в этом основание для решения вопроса о
Глава 2. БИОТЕХНИКА РАЗМНОЖЕНИЯ
Глава 2. БИОТЕХНИКА РАЗМНОЖЕНИЯ 2.1. ЕСТЕСТВЕННОЕ ОСЕМЕНЕНИЕ Вольное спаривание — естественный способ размножения собак. У самок могут быть моно- и полигамные половые акты. При моногамном спаривании собаки совершают по одному-два коитуса ежедневно с одним самцом на
Способы размножения
Способы размножения Размножение — важнейший биологический процесс, обеспечивающий поддержание и увеличение численности вида, возможность его расселения и, в конечном итоге, успех борьбы за существование. В животном мире существует целый ряд способов размножения,
4.1. Виды размножения
4.1. Виды размножения В процессе эволюции живых организмов происходила и эволюция способов размножения, разнообразие которых наблюдаются у ныне живущих видов. Все варианты размножения можно разделить на два принципиально отличающихся типа – бесполое и
Органы размножения кобеля
Органы размножения кобеля То, о чем я собираюсь здесь рассказать, не содержит ничего нового для серьезного собаковода, тем не менее, краткое описание анатомии племенного кобеля может быть для кого-то полезным.Предстательная железаНепосредственно под мочевым пузырем
Органы размножения суки
Органы размножения суки Женские половые клетки — яйцеклетки — производятся в яичниках. Влагалище, матка и маточные трубы — это те пути, по которым проходят сперматозоиды до оплодотворения яйцеклетки.ЯичникиЭтот парный орган находится в брюшной полости суки
Органы размножения кобеля
Органы размножения кобеля То, о чем я собираюсь здесь рассказать, не содержит ничего нового для серьезного собаковода, тем не менее, краткое описание анатомии племенного кобеля может быть для кого-то полезным.Предстательная железаНепосредственно под мочевым пузырем
Органы размножения суки
Органы размножения суки Женские половые клетки — яйцеклетки — производятся в яичниках. Влагалище, матка и маточные трубы — это те пути, по которым проходят сперматозоиды до оплодотворения яйцеклетки.ЯичникиЭтот парный орган находится в брюшной полости суки
7. Система органов размножения
7. Система органов размножения Размножение относится к важнейшим отправлениям организма и обеспечивает продолжение рода. Для выполнения функций, связанных с размножением, у собак служит половой аппарат.Половой аппарат кобеля. Половой аппарат кобеля состоит из