Порочные круги в физиологических системах как основа клинической предпатологии
Порочные круги в физиологических системах как основа клинической предпатологии
Первый порочный круг. Ведущие процессы: уменьшение эффективности действия гормонов и недостаток гормонов. Имеются данные о возрастном повышении порога чувствительности гипоталамуса и желез внутренней секреции к соединениям (глюкоза, кортизон, эстрогены, гонадотропины), являющимся звеньями функциональной межсистемной регуляции по принципу прямых и обратных связей. Имеются и данные, свидетельствующие о возможном пострадиационном снижении чувствительности тканей к гормонам некоторых периферических эндокринных желез (тиреоидным, кортикостероидным). Показано также возрастное снижение чувствительности семенников к гонадотропной стимуляции и понижение в старости эффекта действия тестостерона на обменные процессы и морфологию тканей.
Ускоренное снижение чувствительности тканей к медиаторам межсистемных связей может происходить и при воздействии различных внешних факторов и при различных заболеваниях.
Снижение эффективности действия гормонов (в общем случае — медиаторов межсистемных связей) может происходить по трем причинам: 1) ухудшение качества гормонов; 2) изменения в акцепции гормонов; 3) снижение реактивности эффекторного органа в ответ на гормональный сигнал.
В большинстве случаев нет оснований предполагать ухудшение качества гормонов в начальный период зарождения и развития порочных кругов. Усиление выработки собственной железой гормонов (того же качества) успешно компенсирует снижение их сродства с акцептором.
Нарушения в акцепции гормонов могут зависеть от изменения качества и количества гормональных рецепторов. В обоих случаях необходимо анализировать состояния прежде всего плазматической мембраны клеток. Здесь только укажем, что нарушения в акцепции гормонов в начальный период зарождения порочных кругов наиболее вероятны и они связаны, по-видимому, с тонкими изменениями поверхности плазматической мембраны. Получены прямые экспериментальные доказательства возможности изменять порог чувствительности к гормонам (в общем виде — к медиаторам межсистемных связей) воспринимающих их тканей, воздействуя на состояние плазматической мембраны.
Снижение реактивности эффекторного органа в ответ на гормональный стимул, т. е. снижение способности воспринимающих их тканей ответить адекватной реакцией, характерно для возрастных изменений.
При снижении эффективности действия гормонов для выполнения их регулирующей роли организму требуется все большее количество гормонов. Создается состояние повышенных потребностей организма в гормонах. Соответственно стимулируется гормонообразование для осуществления нормальных регуляторных функций со стороны эндокринной системы.
Происходит постоянно нарастающее повышение гормональной активности основных периферических эндокринных желез (щитовидной, коры надпочечников, половой). Так, женские яичники могут увеличивать в 2 раза выработку стероидных гормонов каждые 10 лет. Радиационное воздействие и другие факторы могут ускорять эти процессы. При достаточных физиологических резервах это не приводит к истощению эндокринных функций.
Однако в отягчающих условиях длительное напряжение эндокринных функций может приводить к начальным явлениям истощения их и может отмечаться недостаточное количество гормонов в организме. Это вызывает новую стимуляцию гормонообразования. Так замкнулся первый порочный потенциально патогенетический круг (рис. 7).
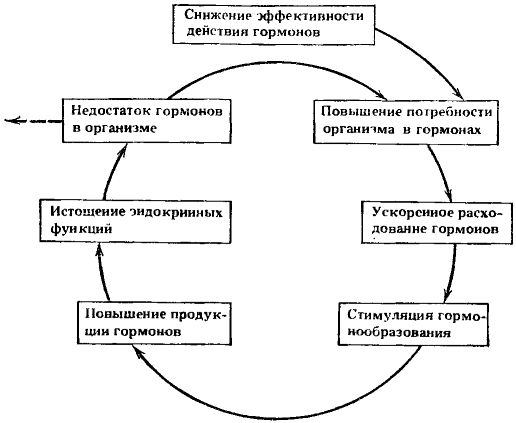
Рис. 7. Схема первого порочного потенциально патогенетического круга
Ведущие процессы: уменьшение эффективности действия гормонов и недостаток в организме гормонов
Длительное функционирование первого порочного круга может приводить к различным последствиям.
1. В сложившихся условиях крайне затруднена оценка функционального состояния эндокринной функции путем измерения концентрации гормонов в крови (тем более в моче). В крови, как в проточной системе, при ускоренном расходовании гормонов и напряжении в функционировании эндокринной железы реальная концентрация гормонов будет сильно флюктуировать и не отражать состояние железы.
2. Недостаток в организме гормона определяется разницей в скоростях его продукции и расходования и непосредственно не связан с состоянием гормонообразования. Недостаток гормона в организме может быть и при повышенном уровне гормонообразования, и при увеличении в размерах железы. Следовательно, нормальные показатели функционирования железы или признаки усиления ее деятельности могут скрывать реальный дефицит гормона в организме.
3. В рассматриваемых условиях могут быть расхождения в оценке состояния железы по гормональной продуктивности и по морфологическим признакам на основе биопсии. В условиях длительного напряжения функций железы общий уровень гормонообразования может складываться за счет работы разных участков железы, находящихся в различных функциональных состояниях (от гипофункциональных до состояний сильной гиперфункции и явного истощения). И наоборот, при сниженном уровне гормонообразования отдельные участки железы могут находиться в состоянии высокой функциональной активности, а отдельные узелки — в состоянии высокой пролиферативной активности. Указанное особенно ярко проявляется при развитии возрастной патологии.
Свидетельством недостатка в организме гормонов, несмотря на усиленное гормонообразование, является отсутствие признаков гормонального токсикоза в этот период, в том числе и при дополнительном введении в организм гормонов. Наоборот, в этих случаях введение в организм гормонов оказывало благотворное действие по целому ряду показателей.
Основной результат функционирования первого порочного круга, имеющий важнейшее значение для создания последующих потенциально патогенетических кругов, — это формирование состояния скрытой эндокринной недостаточности в организме.
Второй порочный круг. Ведущие процессы: усиленная гемоклазия и увеличение массы кроветворной ткани. При длительном функционировании первого порочного круга и при дальнейшем увеличении недостатка гормонов в организме может развиться общая неспецифическая реакция неблагополучия в ответ на скрытую эндокринную недостаточность. Она сопровождается активизацией защитно-охранительных систем организма, в том числе систем клеточного фагоцитоза. Нас интересуют в первую очередь процессы деструкции и фагоцитоза (пожирания) клеток крови, в первую очередь эритроцитов, при эндокринной недостаточности.
В принципе полезная гемокластическая (кроверазрушительная) функция селезенки и других органов, участвующих в деструкции и фагоцитозе эритроцитов, направлена на очищение кровотока от устаревших и поврежденных клеток. Однако при длительной стимуляции ее происходит ускоренная убыль клеток из русла крови. Известно, что в физиологических условиях у человека лишь 10% старых эритроцитов разрушается в кровяном русле. Основная масса эритроцитов разрушается внутриклеточно после захвата их макрофагами печени, селезенки, костного мозга и легких. Кроме того, в эритрофагоцитозе могут участвовать ретикулярные клетки, гистиоциты и полинуклеарные лейкоциты. Селезенка не является основным местом деструкции эритроцитов в норме. В ней разрушается лишь 13—23% этих эритроцитов.
Однако в определенных и патологических условиях селезенка становится настоящим кладбищем эритроцитов. Так, достаточно перевязать селезеночную вену или денервировать ее, как у экспериментальных животных развивается анемия.
Мы приводили уже данные о синдроме гиперспленизма и о прямой зависимости продолжительности жизни эритроцитов от размеров селезенки при ряде заболеваний. Всякое замедление кровотока в селезенке и усиление секвестрации крови (т. е. задержки ее в полостях селезенки) ведут к увеличению вероятности контакта с макрофагами и другими эритрофагоцитирующими клетками, что в условиях селезенки нормальных размеров способствует первичной деструкции эритроцитов. Даже отсутствие первичной усиленной деструкции эритроцитов в селезенке в этих условиях еще не говорит о том, что она не оказывает на них повреждающего действия. В период замедленного прохождения эритроцитов по пульпе селезенки в эритроцитах могут возникать неблагоприятные изменения, которые лишь в дальнейшем приведут их к ускоренному разрушению. При этом прежде всего происходят изменения в гликопротеидах, белках и липидах поверхностных слоев мембраны, облегчающих контакт эритроцитов с макрофагами и другими эритрофагоцитирующими клетками.
Еще раз напомним, что указанные изменения происходят в нормальном организме лишь с целью удаления старых и поврежденных клеток крови. Удаление селезенки не увеличивает продолжительности жизни всей популяции эритроцитов. Однако при стимуляции этих функций в условиях неспецифических реакций неблагополучия организма происходит ускорение разрушения эритроцитов.
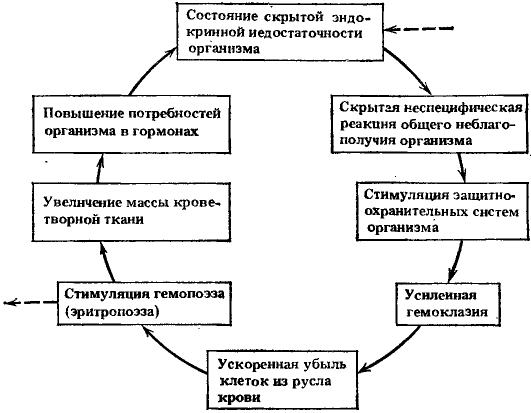
Рис. 8. Схема второго порочного потенциально патогенетического круга
Ведущие процессы: усиленная гемоклазия и увеличение массы кроветворной ткани
Вызывать такие реакции могут разные причины, в том числе и скрытая или явная недостаточность эндокринной системы. Имеются прямые доказательства этому. Ранее постулированное нами состояние скрытой (для клинициста) неспецифической реакции общего неблагополучия организма, ведущее через стимуляцию гемоклазии к ускоренной убыли клеток из русла крови, получило прямое экспериментальное доказательство.
Ускоренная убыль клеток из русла крови вызывает соответствующую стимуляцию гемопоэза. В случаях несильной, но длительной по времени стимуляции гемопоэза, длящейся месяцами, в начале увеличивается продуктивность костного мозга за счет увеличения массы кроветворной ткани. «Желтый» жировой костный мозг трубчатых костей заменяется «красным» костным мозгом, активно продуцирующим эритроциты. В этот период показатели периферической крови и данные миелограммы остаются в пределах нормы. Такие состояния могут продолжаться длительное время и оставаться скрытыми для гематолога, использующего традиционные клинико-лабораторные методы исследования крови. Поскольку эти изменения вначале выражены не резко, они могут быть выявлены лишь специальными изотопными методами, позволяющими количественно определять суммарную продуктивность всего костного мозга организма за период времени, превышающий несколько суток.
Увеличение массы гемопоэтической ткани повышает ее потребность в пластических веществах и в регуляторах обмена, в том числе и в гормонах. Это вновь способствует повышению потребностей организма в гормонах и обостряет уже имеющееся состояние скрытой эндокринной недостаточности.
Так сформировался второй порочный круг, схема которого приведена на рис. 8. Одновременное функционирование обоих порочных кругов может приводить к более явным явлениям недостаточности эндокринной системы и напряжения гемопоэза.
Третий порочный круг. Ведущие процессы: недостаточность лейкопоэза и снижение иммунобиологической реактивности. Длительное функционирование первых двух порочных кругов в конце концов может приводить в глубоко зашедших случаях к зарождению нового, третьего порочного круга. В основе начала его лежит продолжающаяся стимуляция гемопоэза (эритропоэза). Организм уже не может поддерживать нормальный цитологический состав крови путем повышения продуктивности костного мозга за счет увеличения массы кроветворной ткани. Исчерпан также резерв репопуляции полипотентных стволовых клеток при нормальном распределении их дифференцировок в различные ростки кроветворения. В таких случаях в соотношении продукции клеток разных ростков происходит смещение в сторону клеток красного ряда и осуществляется примат продукции клеток этого ростка, так как он обеспечивает главную, т. е. дыхательную, функцию крови. Обоснования этого уже рассматривали.
Уменьшение производства клеток белого ряда приводит к развитию недостаточности лейкопоэза (прежде всего лимфопоэза) со всеми вытекающими из этого последствиями, особенно со стороны иммунитета. Выраженность этих последствий может быть различной.
На первом этапе нормальное количество эритроцитов и гемоглобина в крови обеспечивается за счет несущественного перераспределения пролиферирующих и дифференцирующихся клеток костного мозга в сторону производства эритроцитов с некоторым ущербом для белого ростка. В этот период в периферической крови число лейкоцитов неустойчиво находится в границах, близких к норме. При различных нагрузках и функциональных пробах на лейкопоэз происходят хорошо известные извращения лейкоцитарных реакций (вместо лейкоцитоза наблюдается ареактивность или кратковременное уменьшение их числа).
На втором этапе при более значительном перераспределении пролиферирующих и дифференцирующихся клеток костного мозга в сторону производства эритроцитов продолжает поддерживаться норма функций красной крови при снижении числа лейкоцитов (гранулоцитов и лимфоцитов).
Формы проявления недостаточности лейкопоэза могут быть разными. Это зависит от степени недостаточности поступления ранних пролиферирующих предшественников в различные ростки кроветворения. Если следовать по пути возможных направлений дифференцировок полипотентной стволовой клетки, то на первой развилке выбора направлений дифференцировок уменьшается поток клеток в лимфопозз и, следовательно, в первую очередь будет ослаблена система иммунной защиты организма. Имеются прямые клинические и экспериментальные доказательства этого положения. Наиболее чувствительным показателем недостаточности иммунной защиты организма является снижение устойчивости его к инфекционным факторам, бактериальным и другим токсинам, действию радиации; ослабляется и противораковая устойчивость организма.
Недостаточность иммунной защиты организма может вызывать неспецифическую скрытую реакцию общего неблагополучия организма, вновь стимулирующую защитно-охранительную систему организма, включая гемоклазию и эритрофагоцитоз. Это, в свою очередь, дает дополнительный стимул к дальнейшей активизации гемопоэза. В условиях стимулированного эритропоэза такой стимул означает дальнейшую активизацию его и еще большее преобладание эритропоэза над другими ростками костного мозга.
Замкнулся новый, уже третий порочный потенциально патогенетический круг, представленный в упрощенном виде на рис. 9.
В этих условиях можно предполагать усиление потока клеток-предшественников в сторону миелопоэза, обеспечивающего эритроцитарный, грануло-макрофагальный и тромбоцитарный ростки гемопоэза. На уровне предшественника миелопоэза идет новый выбор направления дальнейшей дифференцировки. Необходимость примата производства клеток красного ростка приводит к развитию недостаточности грануло-макрофагального и тромбоцитарного ростков. Здесь могут складываться многообразные соотношения между этими ростками. По-видимому, меньше должен страдать или совсем не снижать своей активности макрофагальный росток, поскольку активация его эритрофагоцитарной активности происходила раньше в ответ на недостаток в организме гормонов и предшествовала активации эритропоэза.
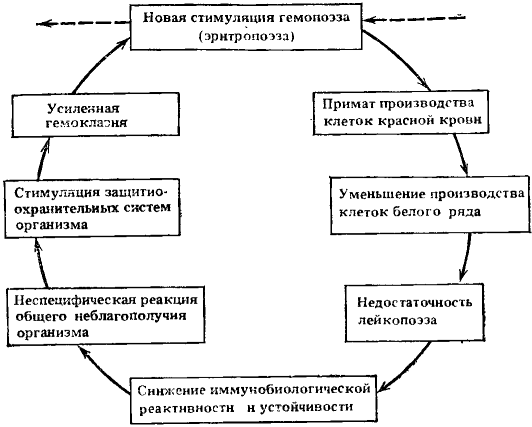
Рис. 9. Схема третьего порочного потенциально патогенетического круга
Ведущие процессы: недостаточность лейкопоэза и снижение иммунобиологической реактивности
Ослабление иммунологической защиты организма должно также стимулировать усиление процессов гуморального и клеточного иммунитета и вызывать опять состояние скрытого неблагополучия организма. Однако в сложившихся условиях (активированный эритропоэз, активированная функция эритрофагоцитоза), по-видимому, всякое иммунологическое обострение уже не будет давать нормально выраженной реакции во всех звеньях гуморального и клеточного иммунитета. Будут прежде всего сказываться недостаточное поступление ранних предшественников в лимфопоэз и примат производства клеток эритроцитарного ряда. Поэтому в ответ на стимуляцию иммунной защиты организма система иммунитета, как мы полагаем, сможет ответить прежде всего теми реакциями, которые были до этого стимулированы, т. е. реакциями усиления гемоклазии, эритрофагоцитоза (см. рис. 9).
Все события, охваченные первыми тремя порочными кругами, еще не дают ярких клинических изменений. При использовании традиционных клинико-лабораторных исследований, обычно проводимых при массовых профилактических осмотрах населения и отсутствии жалоб на здоровье, указанные изменения остаются вне внимания врача. Четкие отклонения от нормы не обнаруживаются: они являются клинически скрытыми и могут быть выявлены лишь при специальных нагрузочных тестах и при использовании особых методов исследования или при обращении к врачу по поводу какой-либо травмы или острой болезни, что вызвало более длительное обследование больного.
Однако длительное функционирование описанных трех порочных кругов может приводить к более глубоким изменениям, обращающим на себя внимание врача и при традиционных методах клинико-лабораторного обследования. Возникает следующий, четвертый порочный потенциально патогенетический круг.
Четвертый порочный круг. Ведущие процессы: дальнейшая интенсификация эритропоэза и ухудшение качества эритроцитов. Длительное и одновременное функционирование трех порочных потенциально патогенетических кругов может в неблагоприятном исходе привести к тому, что показатели красной крови уже не смогут поддерживаться в пределах нормы за счет увеличении массы кроветворной ткани и примата производства клеток красного ряда. Появляются клинически заметные изменения как в системе крови, так и в эндокринной и других системах. Развитие указанных изменений можно разбить на несколько периодов или этапов.
1. В первый период еще не происходит ухудшение качества эритроцитов. Для сохранения дыхательной функции крови в норме начинают использоваться такие возможности, как ускоренный выброс из костного мозга недозревших эритроцитов (ретикулоцитов) в периферическую кровь и увеличение содержания гемоглобина в эритроцитах (увеличение цветного показателя). Общее число эритроцитов в пределах нормы, но число ретикулоцитов увеличено или значительно колеблется. Цветной показатель часто может быть выше нормы. Содержание лейкоцитов — на уровне нижней границы нормы или ниже ее (часто за счет лимфоцитов), может наблюдаться сдвиг лейкоцитарной формулы влево. Эритрофагоцитоз облигатными и факультативными системами клеточного фагоцитоза заметно усилен. В эритрофагоцитозе начинают участвовать ретикулярные и эндотелиальные клетки: их фагоцитоз не связан с состоянием иммунитета. Продолжительность жизни эритроцитов сокращена, иногда весьма значительно. Перекрестные переливания крови показывают, что это сокращение пребывания эритроцитов в русле крови происходит за счет внеклеточных для них факторов.
В этот период устойчивость организма к инфекции, токсинам и радиации снижена. При возникновении травматических ран репаративная реакция вялая и скорость их заживления снижена.
При исследовании периферических эндокринных желез выявляется повышенная функциональная активность щитовидной железы, коры надпочечников и эндокринной половой железы без явлений гормонального токсикоза. При морфологических исследованиях в щитовидной железе и коре надпочечников выявляются очаговые узлы функционального истощения наряду с очагами активной пролиферации.
Изложенное состояние наиболее подробно описано применительно к периоду отдаленных последствий радиационного воздействия после состояния полного клинического выздоровления или к периоду формирования последствий слабых хронических воздействий радиации. Однако они не являются специфическими и возникают при различных иных воздействиях, а также могут быть элементами возрастной патологии.
Часто наиболее чувствительным тестом развития указанных состояний выступает сниженная устойчивость к инфекции и бактериальным токсинам — проявления слабости иммунной защиты организма. Эксперименты показали, что восстановление такой устойчивости может затягиваться на чрезвычайно длительные сроки, составляющие значительную часть всей продолжительности жизни. Это происходит потому, что функции иммунитета являются второстепенными после дыхательной функции крови. Привлекает внимание в этих состояниях и замедление посттравматической репарации. Нами это обнаружено при травмах мягких тканей и костей. Это может быть также следствием перераспределения потоков клеток в различных ростках кроветворения, в том числе и в макрофагально-моноцитарном ростке. Изменения дифференцировок моноцитов в направлении тканевых макрофагов, участвующих в защитно-охранительных реакциях организма в ответ на недостаточность эндокринной системы, в ущерб гистиоцитам соединительной ткани могут объяснить феномен замедления посттравматической репарации.
Важно подчеркнуть, что при возникновении описанных состояний длительное введение в организм в субфизиологических дозах гормональных препаратов щитовидной железы, коры надпочечников и реже половой железы в определенных условиях снимает или существенно ослабляет все перечисленные характерные явления. Наоборот, разнообразные дополнительные нагрузки (например, инфекция, токсины, антигормоны) утяжеляют эти явления и способствуют дальнейшему развитию порочных кругов.
2. Возможным вторым этапом функционирования четвертого порочного потенциально патогенетического круга является состояние с ухудшением качества эритроцитов. С целью сохранения жизненно важной дыхательной функции крови организм вынужден использовать новые резервы компенсации. Повышается пролиферативная активность клеток эритропоэза еще и за счет сокращения продолжительности клеточного цикла эритробластов и возможно более ранних предшественников. Возможность сокращения продолжительности цикла бластных клеток красного и особенно белого ряда костного мозга экспериментально доказана. Для ряда пролиферирующих клеток других тканей также имеются доказательства или это можно предполагать по аналогии.
Имеются общие закономерности для всех клеточных популяций, реагирующих на внешние сигналы и повреждения изменением продолжительности клеточного цикла. Продолжительность собственно митоза, т. е. самого процесса деления клеток, практически не изменяется. Изменяется продолжительность интерфазы, т. е. периодов G1, S и G2.
Наибольшее значение имеет продолжительность периода G1. На этой стадии идут массовый рибосомальный синтез белка, основной рост массы клетки, накопление структурно-мембранных компонентов клетки, все подготовительные мероприятия, необходимые для последующего удвоения молекулы ДНК. Наиболее значительные изменения в своей продолжительности имеет период G1.
Продолжительность периодов G1, S и G2 при различных повреждениях клеток и в ответ на сигналы тревоги и необходимости подготовки к репарации предстоящих повреждений (при стрессовых ситуациях, при введении в организм радиопрофилактических средств и т. п.) увеличивается за счет блоковых задержек. При этом длительность блоковых задержек пропорциональна глубине повреждения клеток. Чем сильнее повреждение, тем больше времени необходимо на их репарацию, которая должна завершиться до перехода клетки в следующую стадию своего развития. Играет роль и изменение скорости репарации повреждений.
Следует сказать, что имеются общие закономерности для гомойотермных организмов, определяющие эффективность восстановления, т. е. полноту устранения возникших спонтанных и индуцированных внешним воздействием повреждений. Под повреждениями здесь понимается вся совокупность возможных разнообразных повреждений, ошибок обмена и синтеза. Эти закономерности представлены на рис. 10. Эффективность работы восстановительных механизмов сложных биологических систем зависит в основном от двух факторов: скорости восстановительных процессов и продолжительности времени, в течение которого могут эффективно осуществляться восстановительные процессы.
Детально изучались возможные границы изменения этих факторов у млекопитающих. Здесь только укажем, что в условиях гомойотермного организма млекопитающих из двух указанных факторов, определяющих эффективность и полноту восстановления, в организме более оперативно и в течение более длительных периодов используется фактор времени. Так, блоковые задержки на стадиях клеточного цикла могут наступать немедленно вслед за повреждающим воздействием. В то же время клетки могут в определенных условиях уходить в период покоя G0 практически на неограниченное время.
Установлено, что в активно пролиферирующих реальных популяциях организма по мере усиления повреждения увеличивается продолжительность блоковой задержки. Однако она происходит лишь до определенного предела. Пока не достигнут предел продолжительности блоковой задержки, клетки, облученные в этой стадии, не имеют гибели ни интерфазного, ни митотического типа (в первом и последующих митозах). При отсутствии у клеток возможностей к дальнейшему увеличению продолжительности блоковой задержки в связи с усилением их повреждения (увеличение дозы облучения) они начинают гибнуть до митоза или во время последующих митозов.
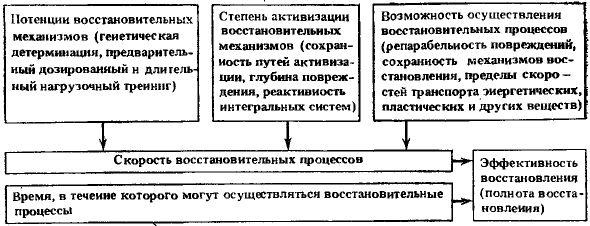
Рис. 10. Факторы, определяющие эффективность и полноту восстановления спонтанных и индуцированных внешним воздействием повреждений
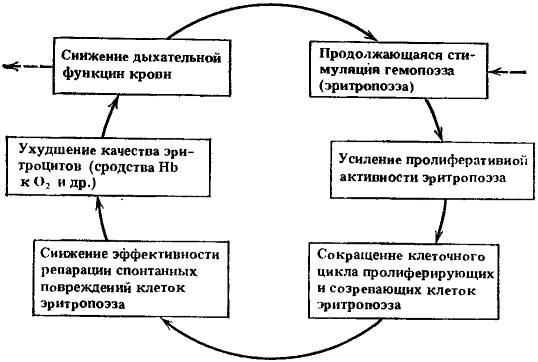
Рис. 11. Схема четвертого порочного потенциально патогенетического круга
Ведущие процессы: дальнейшая интенсификация эритропоэза и ухудшение качества эритроцитов
Следовательно, недостаток времени для осуществления восстановления клетки на данной стадии клеточного цикла от индуцированных повреждений заставляет клетку переходить в следующую стадию с сохранением части повреждения. Остаточные повреждения часто не мешают проходить даже очередной митоз, но они могут кумулироваться до уровня, несовместимого с жизнью клетки, и она погибает через один или несколько митозов.
При значительном усилении пролиферативной активности не всегда адекватный ответ ткани достигается только за счет вовлечения в делящийся пул клеток из покоящегося резерва. Происходит и сокращение продолжительности генеративного цикла клеток, в основном за счет периода G1. При этом повышается чувствительность клеток к повреждающим воздействиям, например к радиации. Увеличивается и вероятность неполного восстановления спонтанных и индуцированных повреждений из-за недостатка времени на их репарацию. Клетки могут переходить из стадии в стадию и вступать в митоз с остаточными повреждениями, способными к кумуляции.
В этом мы видим одну из главных причин появления морфологических изменений (включений), совместимых с жизнью клетки и не имеющих определенного функционального значения. Вторая принципиально возможная причина появления таких изменений может заключаться в отставании поступления пластических и энергетических веществ от значительно ускоренных процессов роста и созревания клетки.
Выше были специально рассмотрены биохимические и биофизические изменения, сопровождающие ускорение процессов пролиферации и созревания клеток. Закономерные изменения, происходящие при значительном усилении пролиферативной активности, могут быть связаны с процессами генетической регуляции и могут носить внегеномный характер.
Наше внимание привлекли ошибки рибосомального синтеза белка, зависящие от изменения рибосомального окружения (ионный гомеостаз, pH и др.) и уменьшения времени на перебор кодон—антикодонных соотношений, происходящих, как предполагается, при сокращении продолжительности периода G1. В этих случаях может включаться ошибочная аминокислота, близкая по своей кодовой специфичности «правильной» аминокислоте, информацию о которой несет матричная РНК. В таких случаях, несмотря на то что ни в ДНК, ни в матричной РНК, ни в рибосоме никаких изменений не произошло, происходит синтез белка с ошибками. Сократилось лишь время на один цикл работы рибосомы. Одна или несколько таких ошибок не могут существенно повлиять на активный центр фермента, но могут изменять антигенные свойства белка и модифицировать ряд физических характеристик белка и белоксодержащих структур (плазматических мембран и др.). Анализ всего пути следования информации от ДНК до функционирующего белка показал, что имеются и другие, но менее вероятные возможности внесения ошибок, влекущих за собой включение ошибочных аминокислот в пептидную цепь белка. Они во многом зависят от изменения окружения информационных молекул, от ионного гомеостаза, от pH и других локальных изменений, которые происходят при значительном усилении пролиферативной активности клеточной популяции и значительном ускорении клеточного деления.
При накоплении проходящих через митоз изменений могут уже ухудшаться качества белковой части гемоглобина, теряться сродство гемоглобина к кислороду, в связи с чем будет снижаться дыхательная функция крови. Это, в свою очередь, будет вновь усиливать стимуляцию эритропоэза.
Таким образом, сформировался новый порочный патогенетический круг, схема которого в упрощенном виде представлена на рис. 11.
Пятый порочный круг. Ведущие процессы: истощение эритропоэза и стимуляция лейкопоэза. Длительное функционирование четырех описанных выше порочных потенциально патогенетических кругов создает все возрастающую стимуляцию функций красной крови по изложенным уже механизмам межсистемных связей. В конце концов это может приводить к начинающемуся истощению эритропоэтической функции и явлениям стойкой анемии различной выраженности. Эта анемия некурабельна при применении гемостимуляторов.
Уменьшение числа эритроцитов и гемоглобина в периферической крови, снижение дыхательной функции крови приводят к мобилизации всех возможных резервов кроветворения для улучшения дыхательной функции. Продукция других ростков кроветворения снижена, начинает проявляться недостаточность лимфопоэза, гранулопоэза и тромбопоэза. В далеко зашедших случаях в крови может отмечаться снижение числа гранулоцитов, лимфоцитов, включая иммунокомпетентные клетки, и реже тромбоцитов. Картина изменений цитологического состава может быть весьма разнообразной. Чаще преобладает снижение зрелых клеточных форм одного из указанных ростков, но могут наблюдаться и сложные отношения, включая панцитопению. Проявления подобных состояний достаточно хорошо известны клиницистам.
Указанные процессы развиваются на фоне скрытой или уже явной эндокринной недостаточности, скрытой или уже ставшей явной недостаточности иммунологической защиты организма. Начинает проявляться накопление модифицированного белка в разных белоксодержащих структурах, и в первую очередь в мембранах, в том числе в наружной клеточной мембране, что может отражаться на акцепторных свойствах ее по отношению к медиаторам межклеточных и межсистемных связей.
Эндокринная недостаточность начинает усугубляться и ухудшением качества гормонов в связи с невозможностью в течение очень длительных сроков отвечать на постоянный стимуляционный сигнал увеличенной продукцией полнокачественных гормонов. Железа начинает выделять недозревшие и некачественные гормоны или их малоактивные аналоги. Эти процессы наиболее подробно изучены применительно к продукции гормонов женским яичником. Яичник начинает вместе с эстрадиолом вырабатывать и неклассические фенолстероиды, уже не обладающие всеми свойствами женского полового гормона. По каналам обратной связи это приводит к активизации выделения гипофизом гонадотропного гормона, что вновь стимулирует яичник к усиленной выработке гормона. Возникают стойкие очаги активной клеточной пролиферации в паренхиме железы.
Сложные отношения возникают и в системе иммунитета. Известно определенное противостояние клеточного и гуморального иммунитета, особенно в условиях недостаточного поступления клеток — ранних предшественников в лимфопоэз. При усилении клеточного иммунитета может ослабляться гуморальный иммунитет и наоборот. При значительном примате клеточной продукции миелопоэза могут быть ослаблены все иммунные функции и клеточный и гуморальный иммунитет. В условиях функционирования описанных порочных кругов, по-видимому, клеточный иммунитет может быть ослаблен меньше, чем гуморальный, поскольку первый в определенной мере участвует в усиленной эритроклазии и эритрофагоцитозе. Недостаток иммунных функций создает также состояние, вероятно, общего неблагополучия организма и вызывает по каналам обратной связи стимул к усилению их.
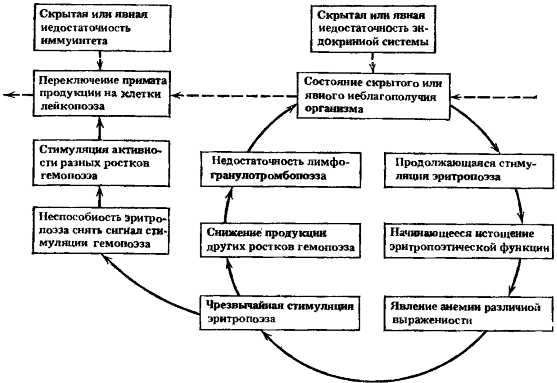
Рис. 12. Схема пятого порочного потенциально патогенетического круга
Ведущие процессы: истощение эритропоэза и стимуляция лейкопоэза
Усугубление всех указанных процессов при возникшей неспособности эритропоэза снять или существенно ослабить стимуляционный сигнал к активации гемопоэза выправлением дыхательной емкости крови приводит к стимуляции разных ростков кроветворения. В одних случаях появляется полицитемия, в других — начинает преобладать один из неэритроцитарных ростков. Чаще происходит переключение примата производства клеток в направлении лейкопоэза (лимфопоэза или гранулопоэза), но возможно и в направлении тромбопоэза и моноцитопоэза. Все это происходит в условиях реакции скрытого или явного неблагополучия организма при определенных нарушениях межсистемных взаимодействий. Схема основных событий пятого порочного круга представлена на рис. 12.
Шестой порочный круг. Ведущие процессы: примат лейкопоэза и нарушения качества и количества лейкоцитов. Шестой порочный потенциально патогенетический круг рассмотрен на примере клеток белого ряда. Значительная стимуляция пролиферативной активности клеток белого ряда, продолжающаяся длительные сроки, может приводить к сокращению продолжительности клеточного цикла делящихся и созревающих клеток, участвующих в лейкопоэзе (полипотентная стволовая клетка, коммитированная в сторону лимфопоэза или миелопоэза с дальнейшим направлением дифференцировки в сторону гранулоцитарного или моноцитарного ростков, миелобласты и миелоциты или монобласты и моноциты). Известно, что в случае такой стимуляции клетки белого ряда реагируют более значительным укорочением продолжительности клеточного цикла, чем клетки красного ряда. Возможность снижения эффективности и полноты репарации спонтанных и индуцированных повреждений, ошибок обмена и синтеза мы также уже обсуждали. По-видимому, клетки белого ряда, как выполняющие менее значимую функцию, чем клетки красного ряда, не имеют столь мощных систем репарации. Известно, что эритропоэз может увеличивать свою продуктивность в 6—8 раз после массивных кровопусканий, сокращать при этом продолжительность клеточного цикла для клеток делящегося и делящегося—созревающего пула. Тем не менее очень часто не наблюдаются какие-либо морфологические изменения клеток или включения в них. Для клеток белого ряда характерно появление разнообразных морфологических отклонений или включений при стимулированном лейкопоэзе.
Поэтому имеются основания считать, что в клетках белого ряда в отличие от клеток красного ряда быстрее будут накапливаться недорепарированные повреждения, ошибки обмена и синтеза, чем в клетках красного ряда. Имеются в виду повреждения, переходящие из одной стадии в следующую и через митоз. Быстрее будут ухудшаться качество лейкоцитов, их функциональные свойства. Изложенное приводит к снижению иммунологической защиты организма и к новой стимуляции лейкопоэза.
Указанные события замкнули новый, уже шестой порочный патогенетический круг (рис. 13), функционирование которого может наращивать силу стимуляционного сигнала к дальнейшей активации лейкопоэза. Продолжающаяся стимуляция лейкопоэза, ускоренное деление клеток, накопление в них повреждений, сохраняющиеся во многих поколениях клеток биофизические и биохимические изменения, характерные для усиленно пролиферирующих клеток, в конце концов могут приводить к автономности ритма ускоренного клеточного деления, к редуцированному клеточному метаболизму и дедифференцированному состоянию. Возникают порочные потенциально патогенетические круги во внутриклеточных процессах, которые будут рассмотрены ниже. Все изложенное формирует состояния, способствующие малигнизации клеток.
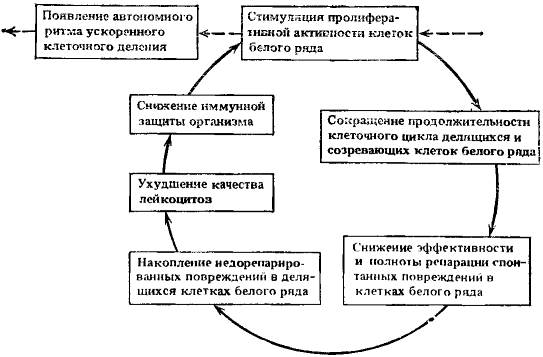
Рис. 13. Схема шестого порочного потенциально патогенетического круга
Ведущие процессы: примат лейкопоэза и нарушения качества и числа лейкоцитов
Рассмотренные порочные потенциально патогенетические круги в физиологических системах не исчерпывают всех возможных взаимосвязей, последовательно вовлекающих новые процессы и новые физиологические структуры, способствующие переходу предпатологии в клинически выраженные формы. Внутри каждого порочного круга также не исчерпаны все активно действующие взаимосвязи. Тем не менее сделанные упрощения позволили описать шесть последовательно возникающих порочных кругов в физиологических системах.
Для нас было важно получить такую последовательность событий в результате функционирования порочных кругов, которая соответствовала бы: 1) последовательности клинических проявлений в период между воздействием радиации или другого лейкозогенного агента и заболеванием лейкозом; 2) возможной последовательности развития клинических синдромов общей гемобластозной болезни; 3) последовательности изменений в различных системах (кроветворения, эндокринная, иммунная и др.), предшествующих появлению отдаленной радиационной патологии, и могла их объяснить.
Кроме того, порочные круги должны были иметь определенную связь с возрастными изменениями, поскольку частота возникновения лейкозов, так же как и других злокачественных заболеваний, зависит от возраста организма. Следовательно, процессы и события, ведущие к предлейкозу, должны быть неспецифическими и общими с целым рядом других предпатологических состояний.
Описанные взаимосвязи, создающие порочные потенциально патогенетические круги, отвечают указанным требованиям и хорошо объясняют всю последовательность событии. Возможность диагностического использования этой последовательности событий будет рассмотрена в последних разделах.
Изложение порочных кругов мы начали с изменений в эндокринной системе, в которой раньше, чем в других системах нормального человека, выявляются значимые для организма возрастные изменения. Это не значит, что развитие порочных кругов всегда должно начинаться с изменений в этой системе. Источником зарождения порочных кругов могут быть и другие самые разнообразные причины. Они могут начинать порочные круги в различных пунктах. Например, хронические и токсические воздействия — с реакций общего неблагополучия организма и стимуляции защитно-охранительных систем организма; хронические нейрогенные и некоторые лекарственные — с явлений стойкой гиперсеквестрации крови в селезенке и ускоренного разрушения клеток крови; различные иммуннопатологические факторы — также с ускоренной гемоклазии; онкогенные вирусы, по-видимому, будут ускорять накопление повреждений в бластных клетках белой крови и их предшественниках.
Вероятность прогрессивного развития каждого патогенетического круга зависит от наступающего истощения репаративных и компенсаторно-восстановительных возможностей системы. Последние генетически предопределены. Различная предопределенность их обусловливает индивидуальные отличия в формировании порочных кругов и в вероятности завершения их переходом от состояния предпатологии к клинически выраженной патологии. Многое зависит от процессов, происходящих на субклеточном уровне, которые также могут формировать свои порочные патогенетические круги.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Восковая основа
Восковая основа Если бы попробовать изобразить внутренность современного рамочного улья в увеличенных, «человеческих» масштабах, пришлось бы нарисовать запрятанное в грандиозный глухой куб поселение, опрокинутое над ровной и пустой квадратной площадью размером
ДНК — МОЛЕКУЛЯРНАЯ ОСНОВА ГЕНОМА
ДНК — МОЛЕКУЛЯРНАЯ ОСНОВА ГЕНОМА Сколько истин, признаваемых нами в настоящее время бесспорными, в момент провозглашения их казались лишь парадоксами или даже ересями! Екатерина II Что мыслимо — то возможно, что возможно — то мыслимо. Г. Лейбниц Тонкое устройство
ДНК — МОЛЕКУЛЯРНАЯ ОСНОВА ГЕНОМА
ДНК — МОЛЕКУЛЯРНАЯ ОСНОВА ГЕНОМА Сколько истин, признаваемых нами в настоящее время бесспорными, в момент провозглашения их казались лишь парадоксами или даже ересями! Екатерина II Что мыслимо — то возможно, что возможно — то мыслимо. Г. Лейбниц Тонкое устройство
НА КРУГИ СВОЯ
НА КРУГИ СВОЯ Продолжим нашу беседу об отношениях между насекомыми и растениями. Сначала скажем несколько добрых слов о растениях вообще, потому что они — травы, кустарники и деревья, — изобретя хлорофилл, взяли на себя величайшую ответственность за судьбу всех
Общие черты предпатологии радиационной и нерадиационной природы
Общие черты предпатологии радиационной и нерадиационной природы Наличие длительного скрытого периода развития радиационного лейкоза, так же как и других отдаленных радиационных последствий, заставило обратить наше внимание на более тщательное изучение того внешне
Порочные потенциально патогенетические круги в физиологических и внутриклеточных системах
Порочные потенциально патогенетические круги в физиологических и внутриклеточных системах Биофизический анализ совокупности экспериментальных и клинических материалов о некоторых пострадиационных, возрастных и предлейкозных изменениях позволил обнаружить и
Порочные круги во внутриклеточных системах как основа клеточной патологии
Порочные круги во внутриклеточных системах как основа клеточной патологии Вводные замечания можно распространить и на порочные потенциально патогенетические круги во внутриклеточных системах. Следует сказать, что в клетке труднее, чем в физиологических системах,
Основа
Основа Прежде чем мы начнем что-то мастерить, следует в немногих словах обрисовать основные положения метода.Все дело заключается в том, чтобы, используя подходящую основу, прочно удерживать в вертикальном положении влагоемкий органический субстрат, который должен
Применение витаминов в клинической практике
Применение витаминов в клинической практике Применение витаминов в профилактических и лечебных целях можно систематизировать следующим образом.В профилактических целях:1. Профилактика первичных гипо-авитаминозов, обусловленных:• недостаточным поступлением
Восковая основа
Восковая основа Если бы попробовать изобразить внутренность современного рамочного улья в увеличенных, «человеческих» масштабах, пришлось бы нарисовать запрятанное в грандиозный глухой куб поселение, опрокинутое над ровной и пустой квадратной площадью размером
4.5. Морфогенетические зародыши в биологических системах
4.5. Морфогенетические зародыши в биологических системах На клеточном уровне зародыши морфогенетических трансформаций могут быть морфическими единицами низшего уровня в клетках, которые присутствуют как в начале, так и в конце процесса клеточной дифференциации.
1. Анатомическая основа рефлекса
1. Анатомическая основа рефлекса Практическое использование собаки возможно только при правильном направлении ее поведения в желательную для человека сторону. Это достигается соответствующей дрессировкой собаки. Для того чтобы правильно дрессировать и использовать
Глава 2 Йога -основа пранаямы
Глава 2 Йога -основа пранаямы Корни пранаямы базируются на учении йоги. Поэтому сначала мы предпримем несколько вводных шагов в учении и философии йоги, для того, чтобы войти в ее основы. За несколько веков до Христа великий индийский мудрец и философ Патанджали впервые
Анализ физиологических механизмов восходящих активирующих влияний на кору мозга у голодных животных методом аппликации различных фармакологических веществ А. И. ТУРЕНКО (Москва)
Анализ физиологических механизмов восходящих активирующих влияний на кору мозга у голодных животных методом аппликации различных фармакологических веществ А. И. ТУРЕНКО (Москва) Многочисленные исследования показали, что основным регулятором потребления пищи является