Глава 25. Метаболизм отдельных аминокислот
Глава 25. Метаболизм отдельных аминокислот
Метаболизм метионина
Метионин – незаменимая аминокислота. Метильная группа метионина – мобильный одноуглеродный фрагмент, используемый для синтеза ряда соединений. Перенос метильной группы метионина на соответствующий акцептор называют трансметилированием, имеющим важное метаболическое значение. Метильная группа в молекуле метионина прочно связана с атомом серы, поэтому непосредственным донором одноуглеродного фрагмента служит активная форма аминокислоты.
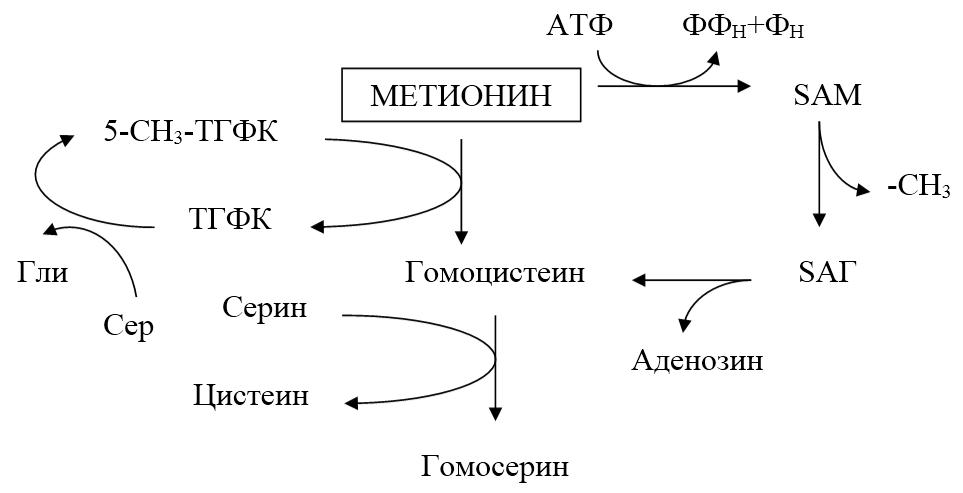
Рисунок 25.1. Обмен метионина.
Реакция активации метионина
Активной формой метионина является S-аденозилметионин (SAM), образующийся в результате присоединения метионина к молекуле аденозина. Аденозин образуется при гидролизе АТФ. Эту реакцию катализирует фермент метионинаденозинтрансфераза, присутствующий во всех типах клеток. Она уникальна для биологических систем, так как является единственной реакцией, в результате которой освобождаются все три фосфатных остатка АТФ. Отщепление метильной группы от SAM и перенос ее на соединение-акцептор катализируют ферменты метилтрансферазы. SAM в ходе реакции превращается в S-аденозилгомоцистеин (SAГ).
Реакции метилирования играют важную роль в организме и протекают очень интесивно. Они используются для синтеза:
1. фосфатидилхолина из фосфатидилэтаноламина;
2. карнитина;
3. креатина;
4. адреналина из норадреналина;
5. метилировании азотистых оснований в нуклеотидах;
6. инактивации метаболитов (гормонов, медиаторов) и обезвреживании чужеродных соединений.
Все эти реакции вызывают большой расход метионина, так как он является незаменимой аминокислотой. В связи с этим играет большое значение возможность регенерации метионина. В результате отщепления метильной группы SAM превращается в SAГ, который при действии гидролазы расщепляется на аденозин и гомоцистеин. Гомоцистеин может снова превращаться в метионин под действием гомоцистеинметилтрансферазы. Донором метильной группы в этом случае служит 5-метилтетрагидрофолиевая кислота (5-метил-ТГФК), которая превращается в ТГФК. Промежуточным переносчиком метильной группы в этой реакции служит производное витамина B12-метилкобаламин, выполняющий роль кофермента. Поставщиком одноуглеродных фрагментов для регенерации 5-метил-ТГФК служит серин, который превращается в глицин.
Синтез креатина
Креатин необходим для образования в мышцах макроэргического соединения креатинфосфата. Синтез креатина идет в 2 стадии с использованием 3 аминокислот: аргинина, глицина и метионина. В почках образуется гуанидинацетат при действии глицинамидинотрансферазы. Затем гуанидинацетат транспортируется в печень, где происходит реакция его метилирования с образованием креатина. Креатин с током крови переносится в мышцы и клетки мозга, где из него под действием креатинкиназы (реакция легко обратима) образуется креатинфосфат – своеобразное депо энергии.
Метаболизм фенилаланина и тирозина
Фенилаланин – незаменимая аминокислота, так как в клетках животных не синтезируется ее бензольное кольцо. Метаболизм метионина осуществляется по 2-м путям: включается в белки или превращается в тирозин под действием специфической монооксигеназы – фенилаланингидроксилазы. Данная реакция необратима и играет важную роль в удалении избытка фенилаланина, так как высокие концентрации его токсичны для клеток.
Обмен тирозина значительно сложнее. Кроме использования в синтезе белков, тирозин в разных тканях выступает предшественником таких соединений как катехоламины, тироксин, меланин и др.
В печени происходит катаболизм тирозина до конечных продуктов фумарата и ацетоацетата. Фумарат может окислятся до СО2 и Н2О или использоваться для глюконеогенеза.
Превращение тирозина в меланоцитах. Он является предшественником меланинов. Синтез меланинов – сложный многоступенчатый процесс, первую реакцию – превращение тирозина в ДОФА – катализирует тирозиназа, использующая в качестве кофактора ионы меди.
В щитовидной железе из тирозина синтезируются гормоны тироксин и трийодтиронин.
В мозговом веществе надпочечников и нервной ткани тирозин является предшественником катехоламинов. Промежуточным продуктом их синтеза является ДОФА. Однако в отличие от меланоцитов, гидроксилирование тирозина осуществляется под действием тирозингидроксилазы, которая является Fe2+-зависимым ферментом, и его активность регулирует скорость синтеза катехоламинов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
3. Перемещения отдельных клеток
3. Перемещения отдельных клеток Многие процессы формообразования происходят не путем перемещения клеточных пластов, а путем перемещения отдельных клеток. Иногда движение клеточного пласта происходит при частичной потере его эпителиальной структуры. Так, например,
Глава 12 Метаболизм
Глава 12 Метаболизм ХимиотерапияБорьба с бактериальными заболеваниями во многом проще, чем с вирусными. Как уже было показано, бактерии проще размножаются в культуре. Бактерии более уязвимы. Живя вне клетки, они производят ущерб организму, отнимая у него питание либо
Глава 496. Почему кодируемых аминокислот двадцать? (XII)
Глава 496. Почему кодируемых аминокислот двадцать? (XII) Неискушенному Читателю может показаться, что элементы машины генетического кодирования описаны в предыдущей главе настолько детально, что к концу чтения он стал даже как-то утомляться, чувствуя, что несколько
Глава 8. Введение в метаболизм
Глава 8. Введение в метаболизм Обмен веществ или метаболизм – это совокупность химических реакций в организме, которые обеспечивают его веществами и энергией, необходимыми для жизнедеятельности. Процесс метаболизма, сопровождающийся образованием более простых
Метаболизм фруктозы
Метаболизм фруктозы Значительное количество фруктозы, образующее при расщеплении сахарозы, прежде чем поступить в систему воротной вены, превращается в глюкозу уже в клетках кишечника. Другая часть фруктозы всасывается с помощью белка-переносчика, т.е. путем
Метаболизм галактозы
Метаболизм галактозы Галактоза образуется в кишечнике в результате гидролиза лактозы.Нарушение метаболизма галактозы проявляется при наследственном заболевании – галактоземии. Оно является следствием врожденного дефекта фермента
Метаболизм лактозы
Метаболизм лактозы Лактоза, дисахарид содержится только в молоке и состоит из галактозы и глюкозы. Лактоза синтезируется только секреторными клетками желез млекопитающих в период лактации. Она присутствует в молоке в количестве от 2 % до 6 % в зависимости от вида
Глава 22. Метаболизм холестерола. Биохимия атеросклероза
Глава 22. Метаболизм холестерола. Биохимия атеросклероза Холестерол – стероид, характерный только для животных организмов. Основное место его образования в организме человека – печень, где синтезируется 50% холестерола, в тонком кишечнике его образуется 15–20%, остальное
Глава 23. Обмен аминокислот. Динамическое состояние белков организма
Глава 23. Обмен аминокислот. Динамическое состояние белков организма Значение аминокислот для организма в первую очередь заключается в том, что они используются для синтеза белков, метаболизм которых занимает особое место в процессах обмена веществ между организмом и
Метаболизм метионина
Метаболизм метионина Метионин – незаменимая аминокислота. Метильная группа метионина – мобильный одноуглеродный фрагмент, используемый для синтеза ряда соединений. Перенос метильной группы метионина на соответствующий акцептор называют трансметилированием,
Концентрация врачей в отдельных регионах
Концентрация врачей в отдельных регионах Если проанализировать карту, характеризующую плотность врачей частной практики, то можно составить следующее представление о 134 существующих департаментах:на юге Франции на одного врача приходится 360 жителей;на севере
Практика отдельных лабораторий
Практика отдельных лабораторий В качестве примера следует привести коммерческие методы, используемые лабораториями Siba Geigy, с помощью которых производят не только лекарства, но и продукцию, предназначенную для сельского хозяйства, в частности пестициды. Эти лаборатории
Мафиозная практика отдельных лабораторий
Мафиозная практика отдельных лабораторий В соответствии с расследованием, проведенным Мишелем Редиа, многонациональное общество Merek, центральное бюро которого находится в штате Нью-Джерси в США, начало заниматься мошеннической деятельностью. Мошенничество
Глава 496. Почему кодируемых аминокислот двадцать? (XII)
Глава 496. Почему кодируемых аминокислот двадцать? (XII) Неискушенному Читателю может показаться, что элементы машины генетического кодирования описаны в предыдущей главе настолько детально, что к концу чтения он стал даже как-то утомляться, чувствуя, что несколько
Принцип отдельных признаков
Принцип отдельных признаков У Г. Менделя был один удивительный предшественник, почти вскрывший основные закономерности наследования признаков за 35 лет до работ Менделя. Это О. Сажре — французский растениевод (1763–1851), автор замечательных работ по гибридизации тыквенных,
От отдельных признаков до фена
От отдельных признаков до фена Английский биолог В. Бэтсон был одним из тех, кто формулировал целые научные направления, прозорливо отличал «существенное от несущественного». Он предложил термин «генетика» в 1906 г. и был организатором первых конференций, по гибридизации,