Глава 496. Почему кодируемых аминокислот двадцать? (XII)
Глава 496.
Почему кодируемых аминокислот двадцать? (XII)
Неискушенному Читателю может показаться, что элементы машины генетического кодирования описаны в предыдущей главе настолько детально, что к концу чтения он стал даже как-то утомляться, чувствуя, что несколько заинтриговавшее его начало книжки оборачивается страницами из учебника для старших классов, способными привести в уныние любого, кто вспомнит родную школу. Искушенному же Читателю, напротив, все рассказанное хорошо известно, и он, грешным делом, подумывает, не написать ли самому учебник посвежее – для тех же старших классов. Не мысля гордый свет забавить – другими словами, не имея намерения вогнать в скуку того и другого, Автор хотел бы подчеркнуть, что понимает: дьявол скрывается в деталях. Но их так много в молекулярной биологии, что любая формализация кажется возмутительным упрощением. Однако, часто бывает, что соблазн формализации неодолим, и тут Автор не может отказать себе в удовольствии еще раз процитировать испанского философа Хосе Ортегу-и-Гассета:
«Серый цвет аскетичен. Такова его символика в обыденном языке, на этот символ и намекает Гете: „Теория, мой друг, суха, но зеленеет жизни древо“. Самое большее, на что способен цвет, не желающий быть цветом, – стать серым; зато жизнь представляется зеленым деревом – какая экстравагантность!.. Элегантное желание предпочесть серый цвет чудесной и противоречивой цветовой экстравагантности жизни приводит нас к теоретизированию. В теории мы обмениваем реальность на тот ее аспект, каким являются понятия. Вместо того чтобы в ней жить, мы о ней размышляем. Но как знать, не скрывается ли за этим явным аскетизмом и удалением от жизни, каким является чистое мышление, наиболее полная форма жизненности, ее высшая роскошь?»
– Браво, Хосе! Именно так я и думаю – даже убежден в этом.
Формализации, теоретизированию, схемам, дизайну генетического кода посвящена основная, хотя и меньшая по объему, оставшаяся часть книги, к которой Автор сейчас переходит. Первая формальная гипотеза структуры генетического кода представляет собой возможный ответ на вопрос, почему кодируемых аминокислот именно двадцать.
В 1954 году Гамов первым показал, что «при сочетании 4 нуклеотидов тройками получаются 64 комбинации, чего вполне достаточно для записи наследственной информации». Он был первым, кто предположил кодирование аминокислот триплетами нуклеотидов и выразил надежду, что «кто-нибудь из более молодых уч?ных дожив?т до его [генетического кода] расшифровки». В 1968 году американцы Роберт Холли, Хар Корана и Маршалл Ниренберг получили Нобелевскую премию за расшифровку генетического кода. Премия была присуждена уже после смерти Георгия Гамова в том же году четырьмя месяцами ранее.
Числа 64 (теоретическая емкость кода) и 20 (фактическая кодирующая емкость, то есть количество кодируемых аминокислот) составляют соотношение правил комбинаторики для размещений и сочетаний с повторами: число А размещений (упорядоченных наборов) с повторами из r (r = 3; размер кодона) элементов множества М, содержащего k (k = 4; число оснований) элементов, равно
Akr = kr= A43 = 64,
а число С сочетаний с повторами из k элементов по r, т. е. любое подмножество из 3 элементов множества, содержащего 4 элемента, равно:
Сkr= [(k+r-1)!] : [r! (k-1)!] = С43= 20.
Это немедленно подводит к мысли о том, что эволюция генетического кода могла начаться с этапа «наборного» кодирования, когда продукт кодировался не последовательностью оснований триплета, а их набором, то есть две такие группы кодонов, как например, САА, АСА, ААС или TGC, TCG, GCT, GTC, CTG, CGT были функционально равнозначны (внутри группы) и направляли синтез одной и той же аминокислоты каждый. Подобные соображения приходят в голову при чтении работ Ишигами и Нагано (1975) – с их идеей о том, что каждая первичная аминокислота могла соответствовать широкому набору кодонов, а также Фолсома (1977) и Трейнора (1984) – с их идеей пермутации оснований в рамках триплета. Очевидно, что меньшее число кодонов не обеспечивало необходимого разнообразия продуктов, а большее было избыточно и, по крайней мере, не соответствовало числу известных сегодня аминокислот. В свое время мы также внесли (очень) скромную лепту в эти идеи, отметив, что число сочетаний из 4 по 3 с повторами иллюстрируется числом квантовых состояний Бозе-газа из трех частиц с четырьмя вероятными собственными квантовыми состояниями54.
Позднее Гамов предложил схему реализации генетического кода, которая предусматривала сборку полипептида непосредственно на молекуле ДНК. По этой модели, каждая аминокислота помещается в ромбической выемке между четырьмя нуклеотидами, по два от каждой из комплементарных цепей. Хотя такой ромб состоит из четыр?х нуклеотидов и, следовательно, число сочетаний равно 256, из-за ограничений, связанных с водородными связями нуклеотидных остатков, возможными оказываются как раз 20 вариантов таких ромбов. Эта схема, получившая название бубнового кода, предполагает корреляцию между последовательными аминокислотными остатками, так как два нуклеотида всегда входят в два соседних ромба (перекрывающийся код). Дальнейшие исследования показали, однако, что эта модель Гамова также не согласуется с опытными данными.
Если бы емкость генетического кода использовалась без остатка, то есть каждому триплету соответствовала бы только одна аминокислота, его защищенность была бы весьма сомнительна: любая нуклеотидная мутация могла оказаться катастрофической. В случае же действующей версии треть случайных точечных мутаций приходится на последние буквы кодонов, половина которых (кодоны октета I) к мутациям не чувствительна вовсе: третья буква кодона может быть любой из четырех – T, C, A или G. Устойчивость к точечным мутациям кодонов октета II в значительной степени определяется двумя факторами – (1) возможностью произвольной замены третьего основания (правда, уже при выборе только из двух – либо пуринов, либо пиримидинов), не меняющей кодируемой аминокислоты вовсе, и (2) возможностью замены пуринов на пиримидины и наоборот, которая сохраняет близкую гидрофильность/гидрофобность продуктов, хотя и не сохраняет их массы. Таким образом, Природа использует чрезвычайно удачный «люфт», называемый вырожденностью кода, когда кодируемому знаку соответствует более, чем один кодирующий.
Эволюция последовательно уточняла функции каждого из трех оснований кодона, что, в конечном счете, привело строгой триплетности только двух кодонов: ATG – для M (метионина) и TTG – для W (триптофана). По способности триплета кодировать только одну аминокислоту отнесем эти два к группе вырожденности I. Когда продукт кодируется фиксированным дублетом оснований, а третье может быть любым из четырех возможных и фактически служит разделителем между функциональными дублетами, говорят об аминокислотах группы вырожденности IV; таких аминокислот восемь: аланин, A, аргинин, R, валин, V, глицин, G, лейцин, L, пролин, P, серин, S, треонин, T. Обобщенный кодон для каждой аминокислоты этой группы, например, лейцина, записывается так: СТN (N—произвольное основание).
Двенадцать кодируемых продуктов относятся к группе вырожденности II; в этой группе третье основание – одно из двух (а не из четырех, как в предыдущем случае): это пурин (R), то есть, либо аденин, А, либо гуанин, G, – или пиримидин (Y), то есть, либо цитозин, С, либо тимидин, Т. К этой группе относятся три аминокислоты, знакомые нам по четвертой группе вырожденности, – аргинин, лейцин и серин, но кодируемые здесь другими дублетами, две пары – аспарагин/аспарагиновая кислота (N/D), и глутамин/глутаминовая кислота (Q/E), а также гистидин H, лизин K, и тирозин Y. Универсальный генетический код относит к этой группе также цистеин С, с его двумя кодирующими триплетами – TGC и TGT, то есть, с третьим пиримидином, а также три стоп-кодона, TAG, TAA и TGA, которые работают только как пунктуационные знаки, фиксирующие окончание гена, но не кодирующие никакой аминокислоты. Обобщенный кодон для аминокислот этой группы, например, аспарагина, записывается так: AAY, а аспарагиновой кислоты – GAR.
Наконец, группа вырожденности III содержит изолейцин, кодируемый тремя триплетами ATA, ATC и ATT. Основания А, С и Т, третьи в кодонах для I, имеют общий символ Н, поэтому обобщенный изолейциновый кодон записывается так: АТН. Все эти особенности кода хорошо иллюстрирует приведенная выше его таблица.
Любопытно, что молекулярная масса кодируемой аминокислоты находится в обратной зависимости от номера группы вырожденности, к которой она относится (В. Щербак). Это первое, отмечаемое здесь, свидетельство очевидной причастности молекулярной массы компонентов генетического кода к его рациональной организации.
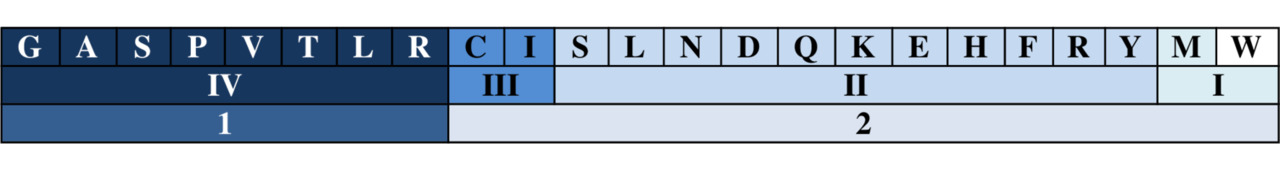
В приведенной табличке упорядоченность по нарастанию молекулярной массы относится к аминокислотам в составе упорядоченных по номерам групп вырожденности (римские цифры), сгруппированным в два октета (арабские цифры). При этом позиция цистеина С скорректирована, о чем речь будет идти в следующей главе; там же мы расскажем и об октетах.
Возвращаясь к выбору именно двадцати аминокислот для кодирования, стоит отметить еще одно интересное обстоятельство: этот выбор мог определяться также квантовой теорией информации, которая предлагает оптимальный алгоритм (алгоритм Гровера) упаковки и чтения информационного содержания ДНК (Апурва Патель, 2001). Такой алгоритм определяет число объектов N, различаемое числом ответов да/нет на вопросы Q, следующим образом:
(2Q +1) sin-1 (1 / ?N) = ? /2.
Решения этого уравнения для малых значений Q весьма характерны:
Q = 1lnN = 04.0
Q = 2lnN = 10.5
Q = 3lnN = 20.2.
Теоретически эти значения не обязательно должны быть целыми числами. Любопытно, что в первом приближении они соответствуют последовательности тетраэдрических чисел, а также эволюции функционального размера кодона от синглетного к триплетному. Другими словами, тетраэдр также можно построить из десяти и из четырех мономеров; эти числа и отмечены в решениях приведенного уравнения. Позднее мы покажем, что комбинация размерных параметров аминокислот и нуклеотидов, базирующаяся на предложенных нами правилах, приводит к пространственному равновесию тетраэдра из двадцати мономеров, соответствующих этим аминокислотам. Здесь же стоит, пожалуй, вспомнить актуальные до сих пор слова В?зе (1973): «Представляется почти жестокой шуткой, что Природа выбрала такое число [кодируемых] аминокислот, какое легко получается в результате множества
математических операций». Но, так или иначе, двадцати альфа-аминокислот (из сотен, встречающихся в природе) оказалось довольно для обеспечения необходимого разнообразия белков.
…………………
Число 496, которым обозначена эта глава, интересно тем, что оно относится к классу так называемых совершенных чисел, и это единственное трехзначное совершенное число. Совершенным называют натуральное число, равное сумме всех своих собственных делителей (т. е. всех положительных делителей, отличных от самого? числа). Сумма всех делителей числа 496, то есть, 1+2+4+8+16+31+62+124+248, равна ему самому. Мы вспомнили о совершенных числах и отмечаем уникальность именно этого числа, потому что оно, во-первых, трехзначно – как трехзначны кодирующие элементы, о которых мы говорим, а во-вторых, как и все предыдущие упомянутые здесь числа, оно – случайно или нет – характеризует один из формальных параметров генетического кода, о которых мы будем говорить дальше. Терпение читателя небезгранично, и Автор вспоминает в связи с этим выдержку из письма одного из читателей известному популяризатору математики Мартину Гарднеру: Перестаньте отыскивать интересные числа! Оставьте для интереса хотя бы одно неинтересное число! Но соблазн велик, и трудно удержаться.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Двадцать пять миллионов лет назад
Двадцать пять миллионов лет назад Жарко в июльский полдень в казахстанской степи. Все залито солнечным блеском: холмистая равнина, расположенные во впадинах и окаймленные камышом озера, желтые участки песков, поросшие саксаулом.Холмы обрываются в сторону озер крутыми
Глава 496. Почему кодируемых аминокислот двадцать? (XII)
Глава 496. Почему кодируемых аминокислот двадцать? (XII) Неискушенному Читателю может показаться, что элементы машины генетического кодирования описаны в предыдущей главе настолько детально, что к концу чтения он стал даже как-то утомляться, чувствуя, что несколько
Глава 23. Обмен аминокислот. Динамическое состояние белков организма
Глава 23. Обмен аминокислот. Динамическое состояние белков организма Значение аминокислот для организма в первую очередь заключается в том, что они используются для синтеза белков, метаболизм которых занимает особое место в процессах обмена веществ между организмом и
Всасывание аминокислот.
Всасывание аминокислот. Происходит путем активного транспорта с участием переносчиков. Максимальная концентрация аминокислот в крови достигается через 30–50 мин после приема белковой пищи. Перенос через щеточную каемку осуществляется целым рядом переносчиков, многие
Наследственные нарушения транспорта аминокислот
Наследственные нарушения транспорта аминокислот Болезнь Хартнупа – нарушение всасывания триптофана в кишечнике и его реабсорбции в почечных канальцах. Так как триптофан служит исходным продуктом для синтеза витамина РР, то основные проявления болезни Хартнупа –
Пути обмена аминокислот в тканях
Пути обмена аминокислот в тканях Аминокислоты – это бифункциональные соединения, содержащие аминную и карбоксильную группу. Реакции по этим группам являются общими для различных аминокислот. К ним относят:1. по аминной группе – реакции дезаминирования и
Трансаминирование аминокислот
Трансаминирование аминокислот Трансаминирование – реакции переноса a-аминогруппы с аминокислоты на a-кетокислоту, в результате чего образуются новая кетокислота и новая аминонокислота. Реакции катализируют ферменты аминотрансферазы. Это сложные ферменты, коферментом
Дезаминирование аминокислот
Дезаминирование аминокислот Дезаминирование аминокислот – реакция отщепления a-аминогруппы от аминокислоты с выделением аммиака. Различают два типа реакций дезаминирования: прямое и непрямое.Прямое дезаминирование – непосредственное отщепление аминогруппы от
Непрямое дезаминирование аминокислот
Непрямое дезаминирование аминокислот Большинство аминокислот не способно дезаминироваться в одну стадию, подобно глутамату. Аминогруппы таких аминокислот перносятся на ?-кетоглутарат с образованием глутаминовой кислоты, которая затем подвергается прямому
Декарбоксилирование аминокислот
Декарбоксилирование аминокислот Некоторые аминокислоты и их производные могут подвергаться декарбоксилированию. Реакции декарбоксилирования необратимы и катализируются ферментами декарбоксилазами, нуждающимися в пиридоксальфосфате в качестве кофермента.
Глава 25. Метаболизм отдельных аминокислот
Глава 25. Метаболизм отдельных аминокислот Метаболизм метионина Метионин – незаменимая аминокислота. Метильная группа метионина – мобильный одноуглеродный фрагмент, используемый для синтеза ряда соединений. Перенос метильной группы метионина на соответствующий
Что означает выражение «двадцать пятый кадр»?
Что означает выражение «двадцать пятый кадр»? Впервые этот термин появился в середине минувшего столетия в США и относился к кино. Дело в том, что киносъемочная камера и соответственно проекционный киноаппарат продвигают пленку со скоростью 24 кадра в секунду. Но в 1957
Двадцать основных причин освобождения от армейской службы во всех призывных пунктах за 1986 г.
Двадцать основных причин освобождения от армейской службы во всех призывных пунктах за 1986 г. №п/п Заболевания и патологические состояния Число лиц, освобожденных от армейской службы Отношение лиц, освобожденных от армейской службы, — к лицам, годным к службе,
ДВАДЦАТЬ ПЯТЬ ЛЕТ СПУСТЯ
ДВАДЦАТЬ ПЯТЬ ЛЕТ СПУСТЯ ПРОХОДЯТ годы, и крохотная, величиной с наперсток, затерянная в почве зародышевая камера разрастается, становится заметным холмиком. Он наглухо облицован сверху глиной, песком, цементом, и в этой мертвой снаружи и немой, как камень, глыбе течет