5. 3. 4. Гомологи катехоламиновых нейронов педальных ганглиев
Во всех разобранных до сих пор примерах клеточных гомологий имеющиеся материалы не давали возможности обсуждать вопрос о том, каким способом специфические нейроны появились в ганглии. Напомним, что априорно можно представить два пути их появления: либо клетки с разной химической специфичностью дифференцировались в процессе эволюции исходной однородной клеточной популяции ганглия, либо они были особенными ещё до ганглионизации. Клетки, о которых пойдёт речь теперь, позволяют исследовать этот вопрос.
Сравнительные данные дают возможность проследить историю катехоламиновых нейронов, имеющихся в некотором числе в составе педальных ганглиев виноградной улитки. Из детального описания её моноаминергических нейронов, данного в разделе 4.3, извлечём несколько положений, важных для последующих сопоставлений. Во-первых, педальные ганглии, связанные с иннервацией ноги, богаче катехоламиновыми элементами, чем все другие ганглии, кроме, может быть, церебральных. Во-вторых, катехоламиновые нейроны составляют лишь малую часть нейронов педальных ганглиев. В-третьих, они расположены в основном группами и в значительной степени у корешков педальных нервов, а иногда в самих этих нервах. В-четвёртых, катехоламиновые аксоны, приходящие по педальным нервам, богато представлены в мышце ноги (их много и в дериватах педальной мускулатуры — в стенке пениса и в ретракторе пениса).
Педальный отдел нервной системы претерпевает в пределах класса гастропод особенно сильную эволюцию, что делает его удобным объектом для анализа эволюционных закономерностей. Среди ныне живущих гастропод имеются представители древних групп, у которых педальный отдел находится на чрезвычайно примитивном уровне организации: ганглии отсутствуют, нейроны образуют в ноге род диффузной сети, несколько усложнённой наличием внутримышечных нервных тяжей. Таковы
Docoglossa, представитель которых, акмея, был взят для данного сравнительного исследования. У форм с более сложной организацией этого отдела нервной системы значительная часть педальных нейронов ганглионизируется, и образующиеся при этом парные педальные ганглии выносятся за пределы ноги, а затем вместе с другими ганглиями ЦНС подвергаются концентрации, образуя компактный мозг. Одновременно идёт процесс гистологической дифференциации педальных ганглиев: в них появляются гигантские нейроны, стратифицируется синаптический нейропиль. В таком продвинутом виде педальные отделы представлены не только у виноградной улитки и других стебельчатоглазых, но и у ряда заднежаберных моллюсков, например у голожаберных (хотя у всех, по-видимому, гастропод какая-то часть педальных нейронов остаётся на периферии, в педальном нервном сплетении). Представитель подкласса заднежаберных — дендронот также был взят для данного сравнительного исследования. Кроме этих форм, представляющих «низший» и «высший» пределы эволюции педального отдела, исследованию были подвергнуты несколько других представителей класса.
Катехоламинергические клетки и волокна выявлялись водным формальдегидным методом. Работа проводилась совместно с А. В. Сахаровой. Чтобы не потерять из виду интересующие нас клетки, мы двигались небольшими шагами, с каждым шагом раздвигая на один таксономический ранг круг охваченных объектов.
Кроме представителей рода Helix, о которых шла речь в главе 5, мы исследовали некоторых других хелицид: Eobania vermiculata и Caucasotachea sp. Анатомическая организация педальной нервной системы у них такая же, как у виноградной улитки. Картина катехоламиновых элементов также сходна: богатый синаптический нейропиль в педальных ганглиях; группы преимущественно мелких нейронов, расположенные так же, как у виноградной улитки; большое число аксонов в мышце ноги, где наблюдаются островки синаптического нейропиля рядом с группками нейронов, не содержащих биогенных аминов.
Вслед за хелицидами были исследованы представители трёх других семейств отряда стебельчатоглазых.
Xeropicta candaharica (сем. Hygromiidae). Небольшое отличие от хелицид заключается лишь в том, что люминесцирующие катехоламиновые аксоны образуют в ноге заметное терминальное сплетение непосредственно под эпителием. Впрочем Роджерс [275а] отмечает то же самое у садовой улитки, но у виноградной выходы неварикозных аксонов под эпителий, хотя и имеют место, не носят такого выраженного характера.
Bradybaena fruticum (сем. Bradybaenidae). Заметных отличий от хелицид в картине катехоламиновых элементов педальной системы не наблюдается.
Limax maximus (см. Limacidae). Хорошо выражены неварикозные, довольно толстые терминальные ветвления катехоламиновых аксонов под эпителием подошвы ноги; варикозный синаптический нейропиль организован в ноге не беспорядочными островками, как у рассмотренных выше видов, а строго ориентированными двумя продольными рядами таких островков. Эти наши наблюдения не соответствуют данным Осборна и Коттрелла, которые, изучив тот же вид наземных слизней, пишут, что в ноге «наблюдается очень мало моноаминовых волокон» [262]. Тела катехоламиновых нейронов расположены в педальных ганглиях примерно в той же позиции, что у виноградной улитки. Размеры мелких, расположенных группами, клеток, по измерениям Осборна и Коттрелла, около 15 мк, тогда как большая непарная клетка, вероятно соответствующая непарной клетке виноградной улитки, достигает у слизня 100 мк.
Литературные данные имеются, кроме того, для Strophocheilus oblongus (сем. Strophocheilidae). Эта гигантская тропическая улитка изучена гистохимически бразильскими авторами [195] и полученные результаты очень интересны: в педальных ганглиях катехоламиновые нейроны отсутствуют. Факт этот тем более примечателен, что в церебральных ганглиях картина серотониновых и катехоламиновых нейронов оказалась типичной для стебельчатоглазых пульмонат, в частности, как уже отмечалось, бразильская улитка имеет пару гигантских нейронов — гомологов ПЦ1 и ЛЦ1. Сообщается, что нейропиль всех подглоточных ганглиев содержит катехоламиновые аксоны, которые предполагаются отростками клеток, лежащих на периферии. К сожалению, авторы не исследовали ногу бразильской улитки.
Кроме отряда стебельчатоглазых, в подкласс лёгочных входит отряд сидячеглазых улиток. Мы исследовали два вида.
У прудовика, Lymnaea stagnalis, имеющего хорошо выраженные педальные ганглии, катехоламиновые нейроны педального отдела отличаются от соответствующих клеток виноградной улитки только, пожалуй, значительно более крупными размерами. Группы этих нейронов расположены в основном в латеральных частях обоих ганглиев. Непарная правая клетка, расположенная, как и у виноградной улитки, дорзально, может быть отнесена к гигантским нейронам. Нейропиль богат катехоламиновыми контактами. В ноге — только аксоны, они выходят к эпителию только у края подошвы. В пенисе, стенка которого имеет педальное происхождение, много варикозных волокон на мышечных клетках субэпителиального слоя мышц.
У американской катушки Helisoma (из планорбид) нами найдена примерно такая же, как у прудовика, картина в педальных ганглиях и несколько отличная в ноге. Здесь совсем отсутствуют островки синаптического нейропиля (у прудовика их было очень мало); гладкие, неварикозные катехоламиновые аксоны, прослеживаемые вдоль педальных нервов от самых педальных ганглиев, проходят через толщу ноги и достигают субэпителиальной области, где далее следуют параллельно поверхности под эпителиальными клетками.
Литературные данные имеются для педальных ганглиев катушки Planorbarius corneus; здесь, как и у прудовика, отмечен непарный гигантский катехоламиновый нейрон, достигающий 280 — 300 мк [238], который, как показали последующие исследования, является интернейроном, возбуждающим одни и тормозящим другие центральные нейроны [92].
Рассмотрим теперь несколько форм, наиболее далеких от виноградной улитки и представляющих два других подкласса — переднежаберных и заднежаберных.
Из заднежаберных имеются данные для двух видов голожаберных слизней — тритонии и дендронота. В педальных ганглиях тритонии М. С. Манохина и Л. В. Кузьмина не нашли катехоламиновых нейронов [37]. Также не нашли их и мы в педальных ганглиях дендронота. В отличие от тритонии, у дендронота мы смогли исследовать не только ЦНС, но и периферию. Как в ноге, так и в её деривате — стенке пениса имеется чрезвычайно богатая сеть варикозных катехоламиновых волокон, контактирующих с мышечными клетками. Эти волокна являются отростками клеток, которые расположены в самих этих органах. Каждая катехоламиновая клетка посылает также гладкий, неварикозный отросток в ЦНС, где такие отростки дают начало варикозному синаптическому нейропилю.
Лежащие на периферии педальные катехоламиновые нейроны дендронота были подвергнуты дополнительному электронномикроскопическому исследованию. В подтверждение данных люминесцентной гистохимии было найдено, что в пенисе эти клетки всегда расположены у внутреннего края мышечной стенки, они мелкие, биполярные. Один отросток идет между мышечными клетками в периферическом направлении. Он древовидно ветвится; по ходу ветвей имеются типичные пресинаптические аксональные расширения, контактирующие с клетками второго (снаружи) мышечного слоя. Эти расширения наполнены секреторными гранулами, которые (типично для катехоламинов) имеют плотное зерно при глутаральдегидной префиксации, но лишены его при фиксации одной лишь четырёхокисью осмия. Никогда не наблюдаются контакты между этими аксонами и мышечными клетками наружного слоя, которые вообще не иннервированы. С противоположной стороны от тела нейрона отходит другой отросток, который вступает в состав нервного стволика, идущего между стенкой пениса и семепроводом. Очевидно, это нерв, следующий в ЦНС. На теле катехоламинового нейрона нередко можно видеть синаптические окончания.
Обратимся теперь к переднежаберным моллюскам. Литературных данных о педальных катехоламиновых нейронах для этого подкласса нет, нами исследованы три вида.
У примитивной акмеи Testudinalia (Acmaea) tessellata, не имеющей педальный ганглиев, катехоламиновые нейроны лежат в ноге на небольшом расстоянии от эпителия. Эти клетки биполярные: наружный отросток, лишённый варикозных утолщений, ветвится под клетками эпителия; внутренний направляется к находящемуся в толще ноги педальному ганглиозному стволу, вступая по пути и в самом стволе в состав варикозного катехоламинового синаптического нейропиля. В самих парных ганглиозных педальных стволах, богатых катехоламиновыми нервными окончаниями, отсутствуют нейроны, содержащие катехоламины.
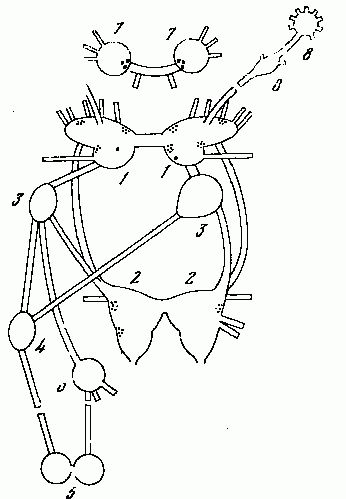
Рис. 14. Центральная нервная система переднежаберного моллюска Pomatiaselegansи позиции центральных нейронов, содержащих биогенные амины. Обозначение ганглиев: 1 — церебральный, 2 — педальный, 3 — плевральный, 4 — супраинтестинальный, 5 — висцеральный, 6 — субинтестинальный, 7 — буккальный, 8 — тентакулярный. В ганглиях 4 — 6 выявление биогенных аминов не проводили. В плевральных и тентакулярных ганглиях моноаминергические нейроны не обнаружены.
Другой исследованный вид — аквариальная «ампуллярия», которая, по сообщению А. А. Шилейко, представляет собой Pila (globosa?) из сем. Pilidae. Улитки этого семейства относятся к группе Architaenioglossa, для которой также характерно отсутствие педальных ганглиев. Парные педальные ганглиозные тяжи связаны многочисленными комиссурами. Такое строение педальной нервной системы рассматривается как древний признак.
В главных педальных стволах и в дочерних тяжах, доходящих почти до подошвы, богато представлены участки синаптического нейропиля с варикозными катехоламиновыми волокнами. Тела катехоламиновых нейронов встречаются в составе ганглиозных стволов. В стенке пениса тела этих нейронов расположены более периферически. Нейроны биполярны, причём периферический отросток не только доходит до эпителия, но и даёт веточки между эпителиальными клетками. Внутренний отросток следует к островкам синаптического нейропиля, расположенным тут же, в стенке органа.
Наконец, третий вид — Pomatias elegans (сем. Pomatiasidae) относится к группе Taenioglossa, представители которой имеют компактные педальные ганглии. В составе этих ганглиев, наряду с большинством нейронов, лишённых биогенных аминов, имеются латерально расположенные группки мелких катехоламиновых клеток (рис. 14). Нейропиль ганглиев богат варикозными катехоламиновыми волокнами. В ноге тоже имеются мелкие катехоламиновые нейроны. Их отростки не подходят к эпителию подошвы. В стенке пениса, которая у этой улитки не является дериватом педальной мышцы, а вырастает из спинной области, катехоламиновых волокон нет совсем.
Эти сравнительные данные позволяют реконструировать картину эволюции катехоламиновых нейронов, входящих в состав педальной нервной системы. Вряд ли можно сомневаться, что их исходная позиция представлена у примитивной акмеи, где эти клетки занимают субэпителиальное положение в подошве ноги. Биполярные эпителиальные или субэпителиальные нейроны обычно рассматриваются как нейроны с сензорной функцией, которую выполняет периферический отросток. Отростки педальных катехоламиновых нейронов проявляют, однако, три рода функциональных отношений. Неварикозный отросток, расположенный под эпителием или входящий в него, очевидно, выполняет сензорные функции; варикозный периферический или центральный отросток, контактирующий с мышечными клетками, мы вправе считать моторным; наконец, варикозный отросток, контактирующий с другими нейронами, служит для осуществления межнейронных связей. Все три рода отношений можно найти в разных группах гастропод. По-видимому, педальные катехоламиновые нейроны, исходно мультифункциональные, в ходе эволюции приобретали разную специализацию, став сензорными в одних случаях, моторными — в других (например, в пенисе дендронота), интернейронами — в третьих.
Непарный дофаминергический интернейрон заслуживает особого упоминания. Берри и Коттрелл [92] исследовали синаптические эффекты этого гигантского нейрона в ЦНС катушки, где он расположен в левом педальном ганглии (ганглий ошибочно обозначен правым в работе [238]). Сравнительно-гистохимические данные, изложенные выше, показывают, что гомологи этой клетки легко прослеживаются в ряду пульмонат. По моим наблюдениям, у прудовика этот нейрон особенно велик и его легко найти в правом педальном ганглии (гомологичном левому ганглию планорбид) рядом с плевро-педальным коннективом; крупные размеры нейрон имеет у слизня, но у хелицид он невелик.
В разных подклассах гастропод педальные катехоламиновые нейроны по-разному вовлечены в процесс ганглионизации. Налицо какая-то избирательность этого процесса, но природа её неясна, хотя общая тенденция прослеживается чётко. Так, среди лёгочных только в примитивном семействе Strophocheilidae нейроны этого типа не обнаружены в составе педальных ганглиев. То, что такие нейроны остались на периферии и у голожаберных, возможно, объясняется их моторными функциями.
Теперь можно ответить на вопрос о том, каким способом катехоламиновые нейроны появились в составе нейронной популяции педальных ганглиев виноградной улитки. Сравнительные данные позволяют с уверенностью утверждать, что клетки с этим типом химической специфичности не возникли в процессе эволюционного дифференцирования исходно однородной нейронной популяции ганглиев: они как специфичная группа существовали до возникновения ганглиев. В процессе дивергентной эволюции гастропод эти клетки ганглионизировались или оставались на периферии; одновременно они приобретали разную функциональную специализацию, что, однако, не сказывалось на их медиаторном химизме.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК