4. 2. 5. Идентифицированные клетки
ЛПл1 и ППл1. Эти симметрично расположенные гигантские клетки проявляют сходные физиологические свойства. Размеры в фиксированном материале до 150 мк, живые клетки немного крупнее. Чаще всего расположены на дорзальной поверхности ганглиев, но иногда позиция тела клетки смещается. Клетки пигментированные, каждая из них выделяется на фоне окружающих мелких нейронов. Визуальная идентификация проста. Обе клетки склонны молчать или генерируют редкие и нерегулярные потенциалы действия в ответ на спонтанные ВПСП. При пропускании слабого деполяризующего тока легко выявляются осцилляторные свойства: появляется регулярная активность мономодального характера без аккомодации к току. ТПСП в спонтанном синаптическом притоке никогда не наблюдались.
Потенциал покоя 45-50, у молчащих нейронов до 60 мВ; потенциал действия 2-компонентный — первый, меньший компонент соответствует, по-видимому, аксонной триггерной зоне, тогда как второй длительностью около 10 мсек и амплитудой около 65-70 мВ, т. е. с небольшим овершутом, является соматическим. Следовая гиперполяризация, как уже отмечалось, бывает значительной величины [283].
Ацетилхолин и серотонин оказывают умеренное деполяризующее действие с выраженной десенситизацией; дофамин как будто неэффективен (эти наблюдения сделаны только на правой клетке).
ЛПа1. Единственный гигантский нейрон, расположенный на заднем конце левого париетального ганглия, у выхода левого мантийного нерва. Чаще всего клетка лежит несколько правее нерва, иногда над ним. В этой области граница между левым париетальным и висцеральным ганглиями изменчива, так что в редких случаях клетка оказывается по другую сторону щели, т. е. в висцеральном ганглии. Таким образом, клетку очень просто идентифицировать визуально. Когда позиция этого нейрона нетипична и он переходит в висцеральный ганглий, его легко отличить от «собственных» нейронов висцерального ганглия по отсутствию активности. При прямом возбуждении пропусканием деполяризующего тока клетка генерирует, как правило, псевдоспайки, при этом залп быстро прекращается вследствие аккомодации к току. В спонтанном синаптическом притоке наблюдаются только мелкие ВПСП. Действие медиаторов не изучено.
Клетка ЛПа1, вместе с ЛПа3 и ППа3, была объектом исследования французских авторов, применявших, в частности, метод фиксации потенциала. Эти авторы дают следующие, средние для всех трёх нейронов, цифры: потенциал покоя — 54±6 мВ (при 23-25°), потенциал действия 92 мВ, трансмембранное сопротивление 1 — 2,5 МОм, постоянная времени 40-50 мсек, откуда входная ёмкость равна около 25 нФ [171].
ЛПа2. Гигантская клетка, расположенная на переднем конце ганглия. В этой области картина крупных нейронов отличается значительным непостоянством: по данным Кунце, на дорзальной стороне в разных препаратах можно видеть до четырёх и на вентральной до трёх гигантских нейронов. Одна из этих клеток белая, что облегчает её индентификацию; белая клетка чаще всего лежит на вентральной стороне ганглия, но иногда заходит на дорзальную, располагаясь медиально. Остальные, пигментированные, гигантские нейроны приходится различать, учитывая их относительную позицию и физиологические свойства. Клетка ЛПа2 всегда расположена латеральнее «молчащего» нейрона ЛПа3.
ЛПа2 проявляет редкую активность или молчит, но, в отличие от ЛПа3, хорошо поддерживает активность при пропускании деполяризующего тока и нередко генерирует в ответ на спонтанные залпы ВПСП. Другое отличие от ЛПа3 — наличие в спонтанном синаптическом притоке как возбуждающих, так и тормозных ПСП. Залпы ТПСП кумулируются в торможение большой длительности. Потенциал покоя около 40- 50 мВ, потенциалы действия с овершутом, без задержки на нисходящей фазе. Ацетилхолин на уровне генерации потенциалов действия гиперполяризует, этот эффект извращается после инъекции в клетку ионов хлора.
ЛПа3 и ППа3. А. В. Карякин, изучавший эти клетки [20], считает их парными нейронами, обладающими сходным и симметричным ветвлением аксонов. Приведём сообщённые этим автором характеристики клеток: размеры около 120 мк; потенциал покоя 56,7±4,7 мВ (температура не указана); потенциал действия 94,8±8,3 мВ, длительностью 5 мсек, однокомпонентный при прямом раздражении и двухкомпонентный при антидромном; действие ацетилхолина деполяризующее; синаптический приток только возбуждающий, ВПСП мелкие, амплитудой 2-3 мВ, длительностью 40-50 мсек, амплитуда ВПСП линейно зависит от величины мембранного потенциала, рассчитанный равновесный потенциал для ВПСП равен -30 мВ.
Мы можем к этому добавить, что обе клетки легко аккомодируются к деполяризующему току и быстро перестают генерировать, так что называть их нейронами пейсмекерного типа, как это делает А. В. Карякин, на наш взгляд, неверно. Нуждаются в проверке и данные о действии ацетилхолина, полученные А. В. Карякиным только в условиях перфузии всего ганглия. В немногих опытах с ионофоретической аппликацией мы наблюдали, что деполяризация, вызванная ацетилхолином, не достигает уровня генерации потенциалов действия; будучи апплицированным на уровне генерации, ацетилхолин вызывал в наших экспериментах гиперполяризацию мембраны (сделано только на клетке ЛПа3). (См. также об этих клетках в работах 120 и 171].
ППа1. В заднемедиальной доле правого париетального ганглия обычно имеется несколько гигантских нейронов, число и позиции которых варьируют. С полной уверенностью поэтому клетку ППа1 визуально идентифицировать нельзя, но всё же при достаточном опыте её можно узнать, руководствуясь следующими признаками: во-первых, это одна из крупнейших, а иногда крупнейшая клетка доли; во-вторых, она всегда примыкает к правой границе доли, т. е. к складке, отделяющей долю от латеральной области ганглия; в-третьих, клетка обычно выглядит более бледной и прозрачной, чем другие гигантские нейроны доли, хорошо пигментированные; обычно оранжеватая клетка ППа1 бывает белесоватого оттенка.
Я остановлюсь на этой клетке несколько подробнее, так как она понадобится нам в следующей главе.
Физиологические свойства клетки ППа1 настолько своеобразны, что позволяют не спутать её ни с каким другим нейроном. Она имеет совершенно особый тип залповой активности, подробно описанный выше (4.2.3.).
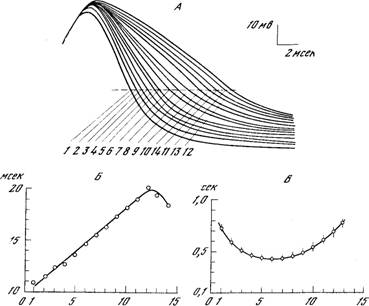
Рис. 9. Характеристики эндогенной залповой активности нейрона ППа1 [Из 289].
А - изменение формы потенциалов действия в течение залпа (цифрой указан номер потенциала в залпе). Б — изменение длительности потенциала действия в течение залпа (показателем длительности служит интервал между восходящим и нисходящим фронтом потенциала действия при 50% его амплитуды). На абсциссе — номер потенциала, на ординате — время, мсек. В — изменение межспайкового интервала в течение залпа. На абсциссе — номер интервала, на ординате — время, сек.
Повторим ещё раз, что потенциалы действия крупные, до 100 мВ, с большим овершутом и задержкой на нисходящей фазе, причём длительность задержки меняется в ходе залпа. Потенциалы генерируются сомой и не имеют аксонного компонента, что также довольно редкое свойство [283].
Клетка ППа1 сильно деполяризуется ацетилхолином, серотонин тоже оказывает на неё возбуждающее действие. Оба медиатора становятся неэффективными в безнатриевой среде, тогда как способность генерировать потенциалы действия в этих условиях сохраняется (прослежено в течение не менее получаса после исчезновения реакции на возбуждающие медиаторы). Удаление ионов кальция из безнатриевого раствора вызывает падение амплитуды потенциалов действия и вскоре полное подавление возбудимости [24]. Дофамин гиперполяризует клетку, вызывая торможение активности.
Необычный тип эндогенной активности, проявляемый нейроном ППа1, привлекает к этому нейрону интерес многих физиологов. Из публикаций последнего времени отметим работу Христофферсена, изучавшего на этом нейроне (а также на ЛПа3) вклад электрогенного насоса в мембранный потенциал [120], и исследование Шаланки с соавторами, которые детально изучили параметры залповой активности [289]. Эти авторы нашли, что при 22° залп состоит из 10 - 20 спайков, причем межспайковые интервалы находятся в пределах 0,3-1,5 сек., длительность залпов составляет 4,0+0,3 сек., а межзалповых пауз — 5,7± 1,5 сек. Величина мембранного потенциала перед спайком составляет 55±5 мв, сразу после залпа до 64+3 мв, амплитуда потенциала действия около 75 мв (рис. 9).
Последняя величина меньше наблюдавшейся нами, что, возможно, обусловлено различиями в составе раствора Рингера. Также, в противоположность нашему опыту, Шаланки с соавторами отмечают, что залповая активность клетки ППа1 обычно представлена в чистом виде, не замаскированная искажениями, которые создаются синаптическим притоком (этими авторами отмечались лишь ВПСП, и то в небольшом количестве и не всегда). Мы, напротив, всегда наблюдаем богатый синаптический приток, включающий, как уже отмечалось выше, не только ВПСП, но и разные ТПСП, в том числе «гигантские», которые в некоторых препаратах бывают двуфазными (первая фаза деполяризующая, вторая гиперполяризующая, причем в ряду таких ПСП кумулируется гиперполяризация). Эта синаптическая бомбардировка в наших экспериментах, как правило, лишает активность того регулярного рисунка, который наблюдается у этой клетки после снятия ПСП. Поскольку несомненно, что Шаланки и соавторы работали с той же клеткой, которую ранее мы с Шаланки картировали, присвоив ей индекс ППа1, причину различий нужно искать в каких-то условиях эксперимента или препаровки.
Ещё одно расхождение касается хода отростка клетки ППа1. Основываясь на результатах исследования срезов после инъекции в ППа1 проционового желтого, Шаланки с соавторами заключают, что отросток не выходит за пределы правого париетального ганглия. По моим наблюдениям, аксон ППа1 покидает этот ганглий и переходит по комиссуре в висцеральный ганглий. Более точные сведения, полученные на других хелицидах, показывают, что из висцерального ганглия аксон затем выходит в составе интестинального нерва [162б, 210]. Гайнер, нашедший, что аксон этой клетки доходит до предсердия, приводит веские физиологические и биохимические доводы в пользу предположения, что ППа1 — пептидергическая нейросекреторная клетка, для которой предсердие служит местом выделения особого, синтезируемого только этим нейроном нейрогормона в кровоток [162а, б].
ППа2. Клетка, по размерам сравнимая с ППа1, т. е. гигантская, оранжевато-прозрачная, всегда лишённая белого материала, часто пигментированная. В отличие от ППа1, которая обладает уникальным набором характеристик, ППа2 как будто является представителем группы нейронов, проявляющих сходные свойства и расположенных по обе стороны щели, разделяющей правый париетальный и висцеральный ганглии. Другие, неидентифицированные члены этой группы, имеют меньшие размеры. В висцеральном ганглии к этой группе, возможно, относится клетка В5.
Для ППа2 и сходных с ним нейронов характерны потенциал покоя величиной 35-45 мВ, потенциалы действия с аксонным и соматическим компонентами и без задержки на нисходящей фазе (амплитуда соматического потенциала около 70- 80 мВ) отсутствие чувствительности к ацетилхолину. Фоновая активность ППа2 изменчива, на инъекцию деполяризующего тока клетка отвечает активностью бимодального характера, тип которой описан в разделе 4.2.3. Серотонин возбуждает эту клетку, дофамин тормозит её. В спонтанном синаптическом притоке видны отдельные и залповые ВПСП, имеются также ТПСП.
ППа3 - описана вместе с ЛПа3.
ППа4. Как правило, эта клетка имеет меньшие размеры, чем ППа1, ППа2 и ППа3. Её, однако, обычно легко узнать благодаря тому, что у медиальной границы скопления бледных клеток группы D эта клетка наиболее богата пигментом.
Как правило, клетка активна, потенциалы действия генерируются с частотой около 1 в сек с очень нерегулярными интервалами. Они имеют довольно характерный вид: спайк возникает без предшествующей фазы медленной деполяризации, он быстр (не более 10 мсек), иногда генерируются сдвоенные спайки, разделенные короткой паузой. Спайки однокомпонентные, амплитудой до 80 мВ, при гиперполяризации амплитуда уменьшается. По-видимому, генерация имеет место только в аксоне, а сома этой клетки невозбудима; при генерации сдвоенных спайков, вероятно, в активность вовлекается вторая аксонная ветвь. Эти признаки активности, наряду с очень крупными ВПСП, резко отличают клетку ППа4 от лежащих поблизости нейронов группы D и клетки ППа1.
Упомянем другие, выявленные нами свойства клетки [24]. Иногда, хотя и редко, в ней наблюдаются залпы гигантских ТПСП, вызывающие остановку активности. При пропускании деполяризующего тока клетка ведёт себя как мономодальный осциллятор. Ацетилхолин вызывает очень сильную деполяризацию, которая сменяется фазой гиперполяризации; интересно отметить, что такой тормозной фазы не бывает после деполяризации, вызванной инъекцией тока, но она бывает после деполяризации, вызванной залпом ВПСП. Возможно, клетка снабжена двумя родами холинорецепторов. Атропин не снимает действия ацетилхолина. Серотонин деполяризует и возбуждает эту клетку.
Исключение ионов натрия из омывающей среды приводит к прекращению генерации потенциалов действия, а также к снятию деполяризующих эффектов ацетилхолина и серотонина. Весьма характерно, что такое влияние безнатриевой среды развивается очень медленно, в течение около 1,5 час. протока, а при остановке протока клетка быстро восстанавливает способность генерировать потенциалы действия. Такой тип поведения некоторых неидентифицированных нейронов подробно исследовал на виноградной улитке В. Д. Герасимов [10]. При исключении ионов кальция генерируется учащённая регулярная активность, амплитуда потенциалов действия уменьшается.
ППа5. Эта крупная пигментированная клетка представлена не во всех препаратах. Она расположена у медиальной границы ганглия на уровне вершины висцерального ганглия и отличается от остальных идентифицируемых и картированных нейронов ганглия своей тормозной реакцией на ацетилхолин, который гиперполяризует эту клетку. Исследована она слабо. Потенциал покоя до 50 мВ, потенциалы действия более 75 мВ, обычно имеется активность, иногда довольно регулярная, но чаще изменённая синаптическим притоком, в котором различимы ВПСП (они растут при гиперполяризации) и залпы гигантских ТПСП. На фоне торможения, вызванного последними, действие ацетилхолина инвертируется.
В1. Очень крупная, прозрачная, умеренно пигментированная клетка, которая хорошо видна, если висцеральный ганглий повернут к наблюдателю задне-правой стороной, где она занимает крайнее угловое положение. В некоторых препаратах это угловое положение занимает крупная белая клетка, которая чаще лежит на вентральной стороне ганглия, и тогда В1 несколько смещена. Клетка не проявляет спонтанной активности, а её реакция на деполяризацию бывает двух форм, что, возможно, связано с разными состояниями клетки. В одних случаях в ответ на инъекцию деполяризующего тока генерируется небольшое число потенциалов действия, их следовые гиперполяризации, суммируясь, гасят генерацию. В других препаратах, встречающихся чаще, в течение и после такого залпа мембранный потенциал непрерывно и медленно уменьшается. На некоторой фазе этого падения пауза, вызванная прекращением генерации потенциалов действия, вдруг сменяется серией быстрых осцилляций, амплитуда которых в разных случаях бывает различной. Эта реакция, найденная нами в клетке В1, не наблюдалась более ни у какого другого нейрона улитки; её подробно исследовал Э. Лабош, посвятивший этому вопросу специальную статью [228].
В остальных отношениях о клетке В1 известно довольно мало. Величина потенциала покоя 45-50 мВ, потенциалы действия около 80 мВ длительностью 15 мсек. В спонтанном синаптическом притоке мы наблюдали ВПСП, амплитуда которых росла при увеличении мембранного потенциала. Ацетилхолин вызывает эффекты, подобные эффектам деполяризующего тока.
В2. Прозрачная, непигментированная клетка средних размеров, лежащая поверх корешка интестинальнго нерва и окружённая мелкими пигментированными нейронами. Единственная в своей области, эта клетка проявляет частую спонтанную активность, что позволяет идентифицировать её с большой уверенностью. Активность нерегулярная, с перебоями, с меняющейся в ходе залпа амплитудой потенциалов действия.
В3. Крупная белая клетка, представленная не во всех препаратах и, по-видимому, являющаяся одним из нейронов белой группы А. Чаще всего нерегулярно активна, потенциалы действия двухкомпонентные, с большим овершутом, с задержкой нанисходящей фазе соматического компонента. Имеются ВПСП иТПСП. Ацетилхолин гиперполяризует, серотонин стимулирует.
В4. Клетка, отличающаяся очень крупными размерами, имеется в указанном на схеме месте у многих, но не у всех особей. Трудно с полной достоверностью утверждать, что это всегда одна и та же клетка, так как её физиологические свойства характерны для целой группы крупных нейронов, имеющихся в этой области (группа F).
В5. Очень крупная клетка, встречающаяся во многих препаратах. По своим физиологическим характеристикам соответствует клетке ППа2.
В6. Зрительная идентификация этой клетки трудна, хотя в общем можно сказать, что клетка занимает позицию, ближайшую к левому париетальному ганглию, на краю выпуклой, нечётко обозначенной дольки в задней части висцерального ганглия. Физиологически она легко отличается от окружающих нейронов, в большинстве своём относящихся к группе F.
Клетку В6 характеризует меняющийся рисунок активности и богатство синаптического притока, в котором выделяются крупные ВПСП и приходящие сериями крупные ТПСП. Иногда возбуждающий приток настолько силен, что он создает постоянную деполяризацию, достаточную для частой и регулярной авторитмичности. Такие периоды могут сменяться периодами полного молчания, вызываемыми залпами ТПСП, или же периодами, когда отдельные нерегулярные спайки генерируются в ответ на отдельные ВПСП. Для этой клетки, как и для В2, к которой она близка физиологически, характерна изменчивость амплитуды потенциалов действия, что, по-видимому, указывает на их аксонное происхождение.
Группы А и В. Белые клетки, расположенные в области переднего конца висцерального ганглия так, что часть их (группа А) попадает в состав висцерального, а часть (группа В) — в состав левого париетального ганглия. Это нейроны, имеющие потенциал действия с аксонным и соматическим компонентами, причем у последнего большой овершут и задержка на нисходящей фазе. К ним приложимы характеристики, указанные Пархоменко для нейронов типа «а» (см. 4.2.3.). Размеры клеток изменчивые, но за редким исключением не очень большие (не более 50-60 мк).
Судя по описанию, именно с этими клетками работала на садовой улитке Ф. Вальд [325а], нашедшая, что натрий-зависимый аксонный потенциал действия отличается у них от преимущественно кальциевого соматического потенциала,- полное повторение вывода, сделанного до этого Н. Т. Пархоменко [46].
Клетки этих групп обычно имеют нерегулярную активность. При деполяризации ведут себя как мономодальные осцилляторы. Имеют ВПСП и отдельные некрупные ТПСП. Ацетилхолин гиперполяризует, это действие извращается после инъекции ионов хлора. Серотонин, деполяризуя, стимулирует пейсмекерный механизм.
Группа С. Небольшое число крупных белых клеток, расположенных вдоль медиальной границы левого париетального ганглия. Исследованы слабо и как будто соответствуют по своим характеристикам клеткам групп А и В.
Группа D. Бледные, реже белые клетки размером до 150 мк, образующие выпуклую долю в правом париетальном ганглии. Размеры и позиция доли изменчивы (особенно в передне-заднем направлении), иногда складка делит её на две доли. Но при всей изменчивости позиций эти клетки всегда лежат латеральнее области, занятой клетками ППа1, ППа2 и ППа4. Число клеток этой группы — около 30.
Эта группа изучалась нами довольно детально как электрофизиологически, так и электронно-микроскопически. Вкратце, физиологические свойства нейронов следующие. По мембранным характеристикам эти клетки соответствуют типу «а», по Н. Т. Пархоменко (см. 4.2.3.). Потенциал действия двухкомпонентный, причём в активности иногда представлен только аксонный компонент («псевдоспайк»), но чаще имеется и соматический компонент с большим овершутом и задержкой на нисходящей фазе. Длительность этого потенциала изменчива, но как будто не менее 30 мсек. Клетки молчат или имеют редкую активность. После залпа, вызванного деполяризацией мембраны, в течение нескольких минут клетка может развить глубокую, до -70 мВ, гиперполяризацию, на фоне которой извращается действие ацетилхолина (он гиперполяризует и тормозит клетку на уровне генерации активности). Серотонин деполяризует и снимает способность гиперполяризоваться после активности; на фоне серотонина клетка ведёт себя как мономодальный осциллятор. Дофамин гиперполяризует.
Клетки группы D обладают отчетливо выраженной способностью генерировать потенциалы действия в безнатриевой среде. Нужно заметить, что в протоке безнатриевого Трис-рингера возбуждающее действие серотонина снимается за несколько минут, т. е. ионы натрия действительно вымываются из околоклеточной среды. Между тем потенциалы действия продолжают генерироваться (проверено в течение более двух часов непрерывного протока, а также в ганглиях, выдержанных в течение 4-6 час. в сменяемом безнатриевом рингере). В этих условиях исключение из раствора ионов кальция или добавление 20 мМ Со2+, блокирующего кальциевые каналы, приводит к быстрому уменьшению потенциалов действия и затем подавлению их генерации. Гиперполяризующее действие ацетилхолина связано с повышением проницаемости для ионов хлора.
Группа Е. Некомпактное скопление относительно небольших, пигментированных клеток, лежащее в правом париетальном ганглии позади коннектива, связывающего этот ганглий с висцеральным. Отчасти лежат между крупными и гигантскими нейронами. Характерный признак — быстрая аккомодация к деполяризующему току, благодаря чему у этих клеток нельзя получить длительно поддерживаемой активности. Ацетилхолин их деполяризует. Возможно, клетки с такими же свойствами лежат в примыкающей области висцерального ганглия.
Группа F. Крупные пигментированные нейроны, лежащие в заднелатеральной части висцерального ганглия (где имеются и клетки с другими свойствами). Для нейронов этой группы характерна довольно регулярная активность, которая определяется богатым притоком ВПСП, создающим постоянный фон деполяризации. ТПСП нет. Потенциалы действия двухкомпонентные, не очень большие (до 50-70 мВ), с небольшим овершутом. Ацетилхолин деполяризует, учащая разряд. Как генерация потенциалов действия, так и эффект ацетилхолина снимаются в течение нескольких минут в протоке безнатриевого Трис-рингера. Серотонин деполяризует.
Группа G. Крупных или средних размеров клетки, часто с многокомпонентной спайковой активностью. Создаётся впечатление, что генерация потенциалов действия имеет место в нескольких аксонных ветвях, тогда как сома неактивна. Характерный признак — тормозное действие серотонина (у зимних улиток). Клетки этой группы на дорзальной поверхности висцерального ганглия не образуют компактной группы и смешаны с другими нейронами, в частности, с относящимися к группе F. Напротив, на вентральной поверхности ганглия в той его дольке, которая находится между корешком анального нерва и левым плевральным ганглием, почти все поверхностные нейроны относятся к этой группе. Расположенные здесь клетки довольно крупные, а одна из них, лежащая у латерального края доли, — гигантская, её легко визуально идентифицировать, и можно зарезервировать за этой клеткой индекс В7. На этой гигантской клетке мы с Г. Н. Коробцовым неоднократно отмечали двойной эффект ацетилхолина — тормозной гилерполяризующий при аппликации на сому и деполяризующий, возбуждающий при аппликации на отросток на расстоянии от сомы. Пока неясно, все ли клетки группы обладают двумя механизмами холинореактивности, как эта гигантская клетка, но по крайней мере некоторые неидентифицируемые нейроны этой области, вероятно, разделяют это свойство.
Гигантские метацеребральные нейроны ПЦ1 и ЛЦ1. На вентральной поверхности каждого из церебральных ганглиев имеется по гигантскому нейрону, расположенному возле корешка внутреннего губного нерва. Это пигментированные клетки размером около 140 мк, легко различимые среди окружающих нейронов меньшего размера. Будучи единственными исследованными идентифицированными нейронами церебральных ганглиев, они могут быть обозначены как клетки ПЦ1 и ЛЦ1. Гистохимическое исследование обнаружило в этих клетках серотонин (см. раздел 4.3.); благодаря этому свойству они представляют для нас специальный интерес, о чём речь пойдет в главе 5.
Кандель и Тауц [204] нашли, что потенциал покоя равен примерно 50 мВ; потенциалы действия, имеющие аксонный и соматический компоненты, достигают, по нашим данным, 60- 70 мВ. Клетка молчит или проявляет довольно продолжительные периоды активности. При раздражении нервов наблюдаются крупные ВПСП, а в спонтанном притоке более мелкие ВПСП от двух интернейронов [204]. Клетки получают также тормозной приток от тентакулярных нервов [304]. Ацетилхолин деполяризует, при этом клетки проявляют хорошо выраженную десенситизацию к ацетилхолину. Действие ацетилхолина блокируется кураре. Глутамат имитирует синаптическое торможение [304]. Клеточная мембрана обладает аномальными выпрямляющими свойствами [205].
Следует упомянуть замечательную серию исследований Коттрелла и соавторов [см. 131, 266а], которые нашли, что ПЦ1 и ЛЦ1 являются интернейронами, дающими возбуждающие синаптические окончания на некоторых идентифицированных клетках буккальных ганглиев. Особенно интересно, что аксонные ветви каждой из двух клеток приходят в каждый буккальный ганглий с двух сторон — через правый и левый церебробуккальный коннективы. Всего, как это нашли ещё Кандель и Тауц, каждый из двух нейронов даёт начало трём аксонным ветвям: одна следует в ипсилатеральный наружный губной нерв, другая — в ипсилатеральный церебробуккальный коннектив и третья — в церебральную комиссуру; попав в симметричный ганглий, эта ветвь вступает в контралатеральный церебробуккальный коннектив.
Хотя клетки содержат серотонин и выделяют его в своих аксонных окончаниях [см. 131], сами они, по нашим наблюдениям, чувствительны к этому медиатору, который их возбуждает. Деполяризующее действие серотонина быстро исчезает в безнатриевой среде, как и способность генерировать потенциалы действия. Этот эффект обратим.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК