2.2.3. Мутационная теория де Фриза в аспекте видообразования
Ч. Дарвин разделял положение о том, что природа не делает скачков. Здесь заключался принимаемый дарвинистами постулат о непрерывности и о неограниченности изменчивости. Принималось, что различия между разновидностями и видами обязаны возникновению мелких непрерывных изменений. Хотя Ч. Дарвин знал, что иногда наблюдаются резкие отклонения от нормы, он считал, что эти своеобразные "уродцы" очень редки и в силу слабой жизнеспособности не имеют эволюционной судьбы.
Этот вывод Ч. Дарвина и его последователей был поставлен под сомнение исследованиями Г. де Фриза, который по праву считается основателем мутационной теории, основные положения которой остаются верны и сегодня (Лобашев, 1967, с. 287). К этому можно добавить, что к славе Г. де Фриза как переоткрывателя законов Менделя следует добавить два важных пионерских концептуальных представления: 1) о нестабильном состоянии генов и 2) о существовании критических периодов в проявлении наследственных признаков (Фриз, 1932; Vries, 1912).
Подробно рассматривая динамику идей в области учения о макроэволюции, В. И. Назаров (1991) приходит к выводу, что де Фриз высказал три кардинальных положения, составивших ядро современной теории прерывистого равновесия и находящихся сейчас в фокусе острых дискуссий: идею о неравномерности темпа мутирования и эксплозивности видообразования, представление о двух разных путях видообразования — "групповом" и "филогенетическом" (это близко к современным представлениям о кладистическом и филетическом видообразовании), а также идею независимости акта видообразования от естественного отбора, борьбы за существование.
С точки зрения истории и методологии науки интересно, что нередко исследователь специально отбирает тот материал для исследования, который соответствует его интуитивной гипотезе. Мендель специально выбрал горох как жесткий самоопылитель и специально выбрал для исследования четкие единичные признаки, поведение которых можно было с легкостью изучать в ряду поколений. Он писал, что "выбор растительной группы, которая будет служить опытам подобного рода, должен быть сделан с наивозможной осторожностью, если мы не хотим подвергнуть риску самый успех опыта". Напротив, де Фриз подыскивал для работ по анализу наследственной изменчивости вид растений, который в силу каких-то внутренних причин отличался бы сильной изменчивостью. Только после того, как Г. де Фриз просмотрел 100 видов, он остановился на энотере. И его выбор оказался фантастически удачен для задуманных целей.
На одном заброшенном картофельном поле вблизи деревни Гильверзум в 1886 г. он обратил внимание на популяцию энотеры, растения, ввезенного из Америки и одичавшего в Европе. В следующем 1887 г. он нашел на этом же поле двух мутантов (забегая вперед в терминологии) и заложил многолетний опыт по изучению частоты возникновения мутантов.
Энотера оказалась уникальной в смысле своей генетической конституции, а изучение закономерностей ее наследственной изменчивости и видообразования таит в себе еще множество загадок. Работая с энотерой, Г. де Фриз получил от первых высеянных в 1886 г. девяти растений около 53 тыс. потомков в период 1886–1899 гг. и среди них около 800, или 1,5 % форм, имеющих резкие отклонения от исходного типа. Эти отклонения были названы мутациями. Мутации возникали внезапно, непредсказуемо, в разных направлениях. В ряде случаев, отдельные мутации захватывали сразу множество признаков и полностью изменяли габитус растения, причем эта совокупность признаков передавалась как дискретная единица. Такие резко отличающиеся друг от друга формы, возникающие внезапно, де Фриз назвал "элементарными видами". К их появлению отбор никакого отношения не имеет. Что касается вида в дарвиновском понимании, то он, по мнению де Фриза, представляет собой совокупность "элементарных видов", впоследствии названных линнеоном.
Совершенно сходные идеи в работе "Гетерогенезис и эволюция. К теории происхождения видов" развил необычайно талантливый ученый, директор ботанического сада при Петербургском университете, академик С. И. Коржинский (1861–1900), проживший всего 39 лет. Гетерогенезисом был назван процесс внезапного сальтационного появления форм, отличающихся комплексом новых признаков. "Эти признаки обладают большим постоянством и неизменно передаются по наследству из поколения в поколение. Таким образом, сразу возникает новая раса столь же прочная и постоянная, как и те, которые существуют с незапамятных времен" (цит. по: Филипченко, 1977, с. 162).
Что касается происхождения садовых растений, то С. И. Коржинский категорически утверждал, что во всех известных ему случаях новые разновидности возникли путем внезапных отклонений от исходных форм. В своих лекциях по генетике, опубликованных в Мюнхене в 1911 г. и в русском переводе два года спустя, Рихард Гольдшмидт заключает, что "среди ботаников заслуга накопления наибольшего числа фактических данных, имеющих отношение к внезапному возникновению растительных форм, принадлежит русскому ботанику академику С. И. Коржинскому" (Гольдшмидт, 1913, с. 135).
Инициированные де Фризом исследования по генетике, цитологии и систематике в роде Oenothera, привели к открытию фантастических механизмов в организации генетической системы размножения, способах наследственной изменчивости и видообразования. В сводке по цитогенетике (Суонсон, Мерц, Янг, 1969) ситуация с энотерой образно названа "бунтом против эволюции". Здесь оказалось "можно то, что нельзя", а именно: сочетание таких элементов генетической системы, каждый из которых сам по себе вреден, а в комбинации дает вполне удовлетворительный результат. Среди них: 1) вовлечение группы или целого набора хромосом в реципрокные транслокации, что обычно приводит к резкому падению фертильности вследствие хромосомной нестабильности гамет и зигот; 2) накопление летальных мутаций; 3) резкое ограничение рекомбинаций, когда транслокационный комплекс передается как целое; 4) неспособность целого набора хромосом проходить либо через женский, либо мужской гаметогенез.
И несмотря на это, завезенные два-три века назад в Европу из Северной Америки растения рода Oenothera образовали сложные гибридные формы видового ранга, по терминологии В. Гранта (1984) "гетерогамные микровиды". Они распространились в Европе как сорняки, а вид Ое. lamarckiana, который специально был выбран де Фризом для исследования, не только распространился по всей Европе, а позднее вновь утвердился в Америке!
Распространено мнение, к сожалению кочующее по разным учебникам и сводкам, что де Фриз обнаружил не мутации, а лишь редкие рекомбинации, выщепляющиеся в потомстве транслокационных гибридов. Но это, во-первых, не так. А во-вторых, в случае энотеры трудно отличить мутации и рекомбинации. С современных позиций очевидно, что процессы мутации и рекомбинации на молекулярном уровне переплетены и разграничение это нередко вопрос терминологии (Хесин Р. Б., 1984, с. 279). Г. де Фриз обнаружил три типа мутаций: генные, хромосомные и геномные. Среди типично генных были "brevistytis" с коротким столбиком цветка и nana — карликовые растения (Cleland, 1935,1962; Грант, 1984; Burnham, 1980).
К хромосомным мутантам, обнаруженным Г. де Фризом и его учениками в первые же годы, относятся трисомики (2n = 15 вместо 2n = 14). Сначала де Фриз выделил их как относительно константные формы, отличающиеся множеством признаков, и лишь затем был определен их хромосомный статус как трисомиков.
Он впервые обнаружил формы, которые в терминах классической генетики соответствуют геномным мутациям. А именно, полиплоидную форму gigas, которая возникла на 4-й год его опытов среди около 14000 исследованных растений. И, наконец, в разные годы среди в общей сложности 54 000 исследованных растений де Фриз зафиксировал возникновение так называемых "полумутантов" (частота их появления 0,06 %). Растения этого типа, названные rubrinervis, характеризовались красными жилками на листьях, широкой красной полосой на чашечке и плодах и слабым развитием луба.
Как позднее было показано (Cleland, 1962), кариотип исходного вида Ое. lamarckiana, с которым работал Г. де Фриз, имел 12 пар хромосом, объединенных в транслокационное кольцо, и одну пару свободную. А "полумутант" rubrinervis имел 4 свободных пары и 6 пар, объединенных в кольцо. Для возникновения такой формы в потомстве отдельных растений рекомбинационным путем необходимо, чтобы в отдельных половых клетках одномоментно происходили множественные рекомбинационные обмены, причем в определенных участках! Такое событие можно представить себе только в рамках современной генетики, когда стало известно, что множественные обмены могут происходить в местах локализации мобильных элементов, а также быть следствием так называемых "транспозиционных взрывов" и множественных упорядоченных транспозиций (Engels, 1983; Герасимова, 1990; Гвоздев, Кайданов, 1986, 1990). Можно суверенностью полагать, что подобные события будут описаны и у энотеры, исследование которой сулит множество открытии в области механизмов наследственной изменчивости.
В эволюционной генетике, помимо энотеры, доводы в пользу множественных одномоментных хромосомных перестроек получены при исследовании хромосомного полиморфизма у роющих грызунов — слепушонок группы Ellobius tancrei, относящихся к надвиду Е. talpinus. Здесь были открыты так называемые "робертсоновские веера" с варьированием в числе хромосом от 54 до 32. При этом получены доводы, что "54 и 32-хромосомные кариотипы должны были возникнуть за счет резкой скачкообразной перестройки всего кариотипа в целом, приведшей к одномоментному слиянию 16–20 акроцентрических хромосом и 8–10 метацентриков" (Воронцов, 1988, 1999).
Наконец, с точки зрения путей видообразования де Фриза следует считать первооткрывателем двух новых типов видообразования: 1) хромосомного, в частности полиплоидного и 2) гибридогенного. В случае гибридогенного видообразования один вид содержит два разных хромосомных комплекса, которые передаются потомству как целое и в разных комбинациях скрещивания "порождают" новые виды. Ученик и последователь де Фриза О. Реннер открыл, что геном вида Ое. lamarckiana, с которым работал де Фриз, состоит из двух транслокационных комплексов gaudens и velans. В них все 7 пар хромосом связаны взаимными обменами и передаются совместно как одна гигантская хромосома.
При скрещивании растений вида Ое. lamarckiana (комплексы velans/gaudens) и Ое. strigosa (комплексы deprimens/stringers) получаются все четыре возможные комбинации, имеющие статус вида и распространенные в природных популяциях (Грант В., 1984, с. 368). Другими словами, в каждом скрещивании происходит "порождение видов", которое можно наблюдать воочию!
Открытые у энотеры транслокационные гибридные видовые системы, названные комплексами Реннера, обладают еще двумя поразительными особенностями. Во-первых, летальностью каждого из них в гомозиготе, так что возникает сбалансированная перманентная гетерозиготность, когда выживают только гетерозиготы. Именно этот, открытый в опытах де Фриза феномен, натолкнул Г. Меллера на идею использовать метод сбалансированных леталей для создания генетического метода количественного учета возникновения мутаций у дрозофилы. Во-вторых, один из реннеровских комплексов передается только через яйцеклетку, а другой — только через пыльцевые клетки, т. е. в гаметогенезе у гибридогенного вида происходит элиминация одного из геномов.
Кратко суммируем значение работ де Фриза для генетики видообразования и эволюционной теории. Г. де Фриз установил неравномерность темпа эволюционного процесса, наличие вспышек мутаций, которые происходят в "мутационные периоды". В период этих вспышек возникают скачкообразные мутации, резко меняющие облик растений, вызывающие резкие изменения в системе воспроизведения, в результате чего сальтационно возникают формы видового ранга. Подобные сальтационные изменения могут происходить повторно. В опытах Г. де Фриза и его учеников и последователей на энотере было открыто хромосомное видообразование (полиплоидия) и гибридогенные виды, образованные на основе транслокационных комплексов.
Справедливо считается, что "классическая дарвиновская концепция понимает видообразование как постепенный градуалистический процесс" (Воронцов, 19886). Генетические исследования де Фриза доказали возможность хромосомного сальтационного (полиплоидия) и гибридогенного видообразования. Оно оказалось повсеместно распространенным у растений. У покрытосемянных растений около 52–58 % видов являются полиплоидами, по преимуществу это аллополиплоиды, растения объединяющие геномы разных видов. Причем, с такими сложными переплетениями полового, бесполого, гибридного размножения, с такими поразительными системами естественной хромосомной инженерии, что приходится расширить концепцию вида до разных по емкости видовых единиц (Грант, 1984, Малецкий, 1995).
Ведущее концептуальное открытие в этой области было сделано в 1917 году датским генетиком О. Винге, который предположил, что у межвидовых гибридов в случае удвоения у них числа хромосом, могут восстановиться мейоз и плодовитость. "Настоящее исследование имеет непосредственное отношение к этой идее. Оно может рассматриваться как экспериментальное обоснование теории гибридного происхождения полиплоидных видов", — таково введение к классической работе Г. Д. Карпеченко (1927). Он впервые синтезировал новую неизвестную в природе видовую форму Raphanobrassica, константный полиплоидный межродовой гибрид между редькой и капустой. Совершенно справедливо Н. Н. Воронцов (1999) называет синтез рафанобрассики первым случаем конструирования нового генома, того, что в конце 70-х стало называться генетической инженерией.
Через три года шведский генетик Арне Мюнтцинг впервые осуществил ресинтез дикорастущего в природе аллополиплоидного вида пикульника рода Galeopsis. Природная хромосомная инженерия создает гибридогенные полиплоидные комплексы видов, открытые и изученные американским ботаником Ледьярдом Стеббинсом. В этих комплексах геномы нескольких диплоидных исходных видов могут вступать между собой во всевозможные гибридные аллотетраплоидные комбинации. Объединяться могут сразу несколько геномов, так что предком одного вида может ни один, а несколько видов, как, например, у обычной мягкой пшеницы, у видов хлопчатника (Грант, 1984).
Неожиданно выяснилось, что гибридогенное, сходное в принципе с энотерой, видообразование регулярно встречается в ряде групп позвоночных и беспозвоночных животных. Гибридогенное видообразование описано уже у 50 видов позвоночных (Боркин, Даревский, 1980, 1989; Кирпичников, 1988; Tunner, Heppich-Turmer, 1991; Даревский, 1995). Оно принимает три основные формы: партеногенез (спермин не нужны для развития яйцеклеток видов-гибридов); гиногенез (спермин нужны лишь для активации развития, но развитие происходит на основе женских гамет и наследование матроклинно); и собственно гибридогенез, когда гибридный вид образуется на основе гибридных зигот, но один из родительских геномов селективно элиминируется в клетках полового пути в ходе гаметогенеза и мейоза.
Гибридогенез оказался широко распространен у скальных ящериц рода Lacerta, живущих в горных районов Закавказья. Среди 18 близких видов комплекса L. saxicola 7 имели гибридогенное происхождение. Все они состояли из партеногенетических самок, способных размножаться без самцов. Но они способны также скрещиваться со своими собратьями из двуполых видов и порождать новые гибридогенные виды. В последнем варианте примерно в 10 % случаев возникают триплоиды. Причем у некоторых групп ящериц, вьетнамских гекконов и агам, триплоиды оказались плодовитыми Поскольку гибридогенез здесь является нормой, то (1) одни и те же партеновиды порождаются вновь и вновь в разное время и (2) одни и те же двуполые виды способны вступать в разные гибридогенные связи и порождать таким образом разные виды! (Даревский, Гречко, Куприянова, 1999).
У животных обнаружено также удивительное явление избирательной элиминации одного из геномов получило название "геномное исключение". У гибридного вида хромосомы одного из родительских видов как бы одалживаются для построения соматических тканей. Вот почему было предложено именовать такой способ видообразования как "кредитогенез" (Боркин, Даревский, 1980). Элиминация одного из родительских геномов в клетках полового пути может быть названа также как "геномоцид". Это явление было описано и у растений в случае отдаленных гибридов ячменя. Природный гибридогенез в сочетании с геномоцидом в зародышевом пути был обнаружен у некоторых видов беспозвоночных.
Особенно поразительной оказалась гибридогенная система у палочников рода Bacillus, где сосуществуют пять способов воспроизведения (Giorgi, 1992; Mantovani, Scali, 1992):
1) обычное бисексуальное размножение, 2) гибридогенез, 3) партеногенез, 4) гиногенез и 5) андрогенез — первый случай такого рода, обнаруженный как регулярное событие в природных популяциях (схема на рис. 1).
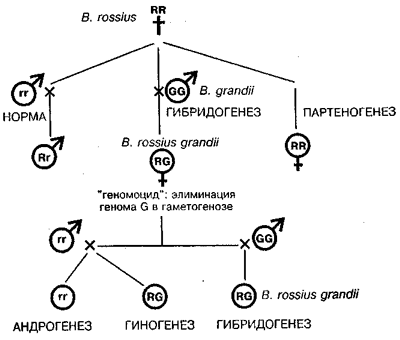
Рис. 1. Гибридогенез у видов палочника (род Bacillus) как пример недарвиновского видообразования, открытого на энотере Г. де Фризом. R и r обозначают геномы вида В. rossius у самок и самцов, G — геном В. grandii (no Mantovani, Scali, 1992; Giorgi, 1992).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК