8.5. Проблема красоты
Еще одна группа фактов, которая не укладывается в современный дарвинизм, зато идеально укладывается в теорию непрерывного творения (и даже предсказывается этой теорией) – это так называемая проблема красоты.
В чем заключается эта проблема, наверное, ясно уже из названия. Все мы знаем, что очень многие живые существа на нашей планете имеют такие признаки, которые трудно объяснить острой необходимостью успешного выживания. Зато эти признаки нередко откровенно эстетичны. То есть, всё выглядит так, как будто целью существования таких признаков является не успешное выживание организма, а чисто эстетические мотивы.
Однако этого не может быть в рамках дарвинизма.
С позиций теории Дарвина, любой признак организма является продуктом естественного отбора (более приспособленных) в условиях борьбы за существование в ходе длительной эволюции жизни на Земле. Точнее, допускается возможность, что признак может быть и нейтральным, следовательно, может возникнуть под действием чисто случайных эволюционных факторов. Но только при условии, что этот признак не отнимает у организма слишком много ресурсов на свое создание и поддержание. Если же признак ярко выражен, то из самой логики теории следует, что этот признак должен иметь какое-то полезное, приспособительное значение. Иначе он был бы давно отсеян естественным отбором (хотя бы в целях экономии ресурсов).
Понятно, что если биологический признак ярко выражен, например, признак является громоздким, или очень сложным, то его появление у живых существ в результате чистой случайности – практически исключено. Над появлением этого признака должна была поработать главная творческая сила дарвиновской теории эволюции – естественный отбор более приспособленных.
Но естественный отбор может работать над созданием только таких признаков, которые полезны для выживания и размножения организмов. А над созданием признаков, бесполезных для выживания (нейтральных), и уж тем более, вредных для выживания – естественный отбор не может трудиться просто по определению (естественный отбор – это автоматический процесс преимущественного выживания более приспособленных).
Таким образом, в рамках теории естественного отбора, ярко выраженных признаков, не полезных для выживания или размножения – у организмов просто не может быть. Максимум, что следует ожидать – это наличие каких-нибудь рудиментов (ранее полезных признаков, а сейчас бесполезных), находящихся в стадии деградации.
Однако, несмотря на столь строгий запрет дарвиновской теории, очень многие живые существа имеют ярко выраженные признаки, которые, вроде бы, совершенно бесполезны для их прямого выживания (или даже вредны). То есть, многие живые существа (причем из самых разных биологических таксонов) имеют разнообразные «архитектурные излишества», которые нередко выглядят еще и эстетичными (хотя это и не обязательно).
Одним из самых одиозных примеров в этом отношении является, конечно, знаменитый хвост павлина, в котором роскошная расцветка сочетается со столь буйной гипертрофией самого «хвоста» (на самом деле, надхвостья), что это просто не может не доставлять павлину неудобств при выживании в дикой природе. И павлин поражает нас не только своим хвостом. Внешность павлина содержит целый ряд признаков, имеющих откровенно эстетическое значение.
Начиная от знаменитых переливающихся глазков на хвосте, и кончая красивым венцом и «маской» на голове (Рис. 47):

Рисунок 47. Самец обыкновенного павлина (Pavo cristatus).[117]
Создаётся ощущение, что весь облик павлина преследует цель максимизации именно эстетической составляющей – многие признаки согласованно вносят свой вклад в усиление общего эстетического эффекта. То есть, в случае самца павлина мы наблюдаем явную эстетическую целесообразность.
Эстетическую целесообразность можно рассматривать, как аналогию биологической целесообразности.
О явной биологической целесообразности живых существ я уже говорил выше. Биологическая целесообразность – это когда мы видим, что определенные признаки живого существа явно предназначены для какой-то определенной биологической цели. Например, никто не будет спорить, что цель наличия глаз (у того же павлина) – это обеспечение способности видеть. Крылья – предназначены для полета. Ноги – для передвижения по земле. И так далее. Биологическая целесообразность живых существ настолько очевидна, что не оспаривается ни кем.
Эта биологическая целесообразность сегодня может быть объяснена двумя принципиально разными способами:
1. Это продукт разумного дизайна.
2. Это результат длительной естественной эволюции в ходе естественного отбора более приспособленных.
Второй вариант объяснения в отношении глаз, крыльев и ног – теоретически вполне возможен. Так как глаза, крылья и ноги, действительно, полезны живому организму, повышая его приспособленность.
Далее по аналогии – мы ясно видим, что целью специфической внешности павлина является максимизация эстетической составляющей. То есть, мы явно наблюдаем эстетическую целесообразность.
Но в отличие от предыдущей (биологической) целесообразности, эстетическая целесообразность может быть объяснена только одним способом:
1. Это продукт эстетического дизайна.
Вторым способом (эволюцией более приспособленных) эта целесообразность объяснена быть не может, потому что чистая эстетика ничем не полезна её обладателю (а в случае павлина – даже вредна).
Причем мы все знаем, что откровенно эстетичная внешность павлина – далеко не единственный такой факт в живой природе, а скорее, лишь крошечная часть общего, огромного массива таких же фактов. Сюда в первую очередь относится яркая, красивая окраска многих живых существ (птиц, рыб, рептилий, насекомых, ракообразных, моллюсков и даже червей). А так же (помимо окраски) другие «архитектурные излишества», которые затрудняют жизнь своим обладателям.
Приходится признать, что проблема красоты родилась вместе с самой дарвиновской теорией. Ведь как только появилась идея, что живые организмы приобретают свои признаки благодаря естественному отбору более приспособленных, так и возникла необходимость объяснять, зачем же множеству живых существ такие признаки, которые неполезны для их выживания. Можно сказать, что дарвинизм родился на свет сразу с этой «родовой травмой» (проблемой красоты). И эта «родовая травма», похоже, неизлечима. Во всяком случае, сегодня факты складываются именно так.
На мой взгляд, эта группа фактов имеет очень большое значение для теоретической биологии. Потому что такие факты не только опровергают дарвиновские представления о развитии живой природы, но еще и (одновременно) свидетельствуют в пользу существования интеллектуальной силы, которая была «повинна» в создании разнообразия живых существ. Действительно, нам (людям) чисто эстетические мотивы (при создании тех или иных вещей) настолько знакомы и понятны, что обнаружение таких же фактов в живой природе (эстетических мотивов) почти единодушно воспринимается нами, как свидетельство в пользу разумного дизайна. Более того, позволяет судить о тех целях, которые преследовала разумная сила при создании живых существ. Вне всякого сомнения, чисто эстетические мотивы при создании разных групп живых существ прослеживаются достаточно часто. А в определенных биологических таксонах – очень часто.
В связи с чрезвычайной важностью (на мой взгляд) этой группы фактов для понимания реальных принципов, по которым создавалась живая природа, я сейчас не буду останавливаться на проблеме красоты подробно. Я посвящу этому вопросу свою следующую книгу. Ведь (еще раз) факты этой категории не только опровергают дарвинизм, но одновременно свидетельствуют в пользу разумного дизайна. А мы сейчас, в основном, занимаемся тем, что критически разбираем «доказательства эволюции», которые насобирали в свою копилку дарвинисты.
Поэтому здесь мы ограничимся лишь тем обстоятельством, которое уже отметили выше – многочисленные факты наблюдаемых «архитектурных излишеств» в живой природе – превосходно укладываются в рамки концепции разумного дизайна (в любой из его вариантов). И одновременно со страшным «скрипом» сопротивляются дарвиновской концепции. Верующие дарвинисты упорно пытаются всё-таки впихнуть эти факты (явной эстетики в живой природе) в свой несчастный «чемодан» дарвинизма, который просто не в состоянии их принять, а факты всеми силами упираются, возмущаясь подобным насилием.
Если коротко, то дела на этом «фронте» сегодня обстоят так:
1. Как правило, верующие дарвинисты пытаются запихнуть все те случаи, когда живые организмы (явно) эстетичны, в свой пресловутый «половой отбор». То есть, предлагается считать, что это просто самцы так перед самками стараются (отбираются), поскольку самки предпочитают именно эстетичных самцов.
А вот почему самки предпочитают именно самых эстетичных самцов (а не, допустим, самых энергичных) – над этим дарвинисты обещают подумать потом («требуются будущие исследования»).
То, что дарвинисты стараются избежать ответа на этот вопрос, вовсе не удивляет. Потому что здесь имеется логическое противоречие. В рамках самого дарвинизма, те самки, которым нравятся биологически бесполезные (и уж тем более, вредные) черты самцов – должны проигрывать (в размножении собственных генов) тем самкам, которые предпочитают самцов с биологически полезными признаками. По сути, самка, предпочитающая самца с вредными признаками – является просто носителем вредной мутации, и должна быть уничтожена естественным отбором. Это логическое противоречие дарвинисты пытаются преодолеть с помощью специальных (чисто теоретических) моделей полового отбора: модель «убегания» Фишера, и модель гандикапа.[118] Однако эти модели, в свою очередь, не могут ответить на первоначальный вопрос – почему же конечный результат этого (воображаемого) отбора самцов под давлением (воображаемых) предпочтений самок – столь часто получается именно красивым. А не, допустим, безобразным. Или просто эстетически нейтральным.
Пожалуй, приведу характерную цитату из соответствующего обзора, посвященного «успехам» теории полового отбора (Jones & Ratterman, 2009):
«Почему украшения, используемые птицами и другими животными, как правило, кажутся столь поразительно красивыми людям – это уже другой вопрос, но это тайна, которую не обязательно решать для понимания полового отбора».
То есть, как я и говорю – что такое половой отбор – понять можем. А вот решить, собственно, проблему красоты – не можем. Это пока «тайна», видите ли. Но если это «тайна», тогда зачем нам вообще теоретические модели полового отбора, если они как раз и создавались (изначально) для того, чтобы объяснить именно «тайну» наличия красоты в живой природе.
В общем, дарвиновские объяснения проблемы красоты получаются, во-первых, слишком громоздкими. Где одна теоретическая модель полового отбора взгромождается на другую модель полового отбора, при этом погоняя третьей моделью полового отбора.[119]
А во-вторых, в конечном итоге, все эти теоретические нагромождения (бесценных дарвиновских идей) всё равно не дают ответа на исходный вопрос – почему обсуждаемые признаки живых существ так часто кажутся эстетически привлекательными? Тем не менее, дарвинисты продолжают упорно цепляться за гипотезу полового отбора. Потому что цепляться больше не за что.
2. Для подтверждения своей гипотезы про «половой отбор», верующие дарвинисты проводят полевые исследования, где пытаются «поймать» соответствующие (эстетические) предпочтения самок. И естественно, много исследований (особенно выполненных ближе к концу 20 века) действительно находили то, что хотелось найти.
3. Гораздо хуже, когда такие (положительные) результаты, полученные в более ранних исследованиях, потом не воспроизводятся в более поздних независимых исследованиях. Что наводит на грустные мысли о том, что «ученые тоже люди» и что подгонку результатов (под нужную теорию) пока еще никто не отменял.
Дело дошло до того, что несмотря на огромное число проведенных исследований, сегодня всё еще нельзя назвать ни одного биологического вида, в отношении которого можно было бы уверенно сказать (на основании нескольких независимых исследований с согласующимися результатами), что данный вид подтверждает теорию полового отбора (Roughgarden & Akcay, 2010).
4. Еще хуже дело становится тогда, когда самец какого-либо биологического вида имеет явно вредный признак, но тем не менее, какую-либо роль этого признака в пресловутом половом отборе так и не удаётся показать. Не удаётся показать сразу в нескольких независимых исследованиях. В случае некоторых животных, даже множество проведенных исследований всё равно оказались не в состоянии найти ожидаемую связь между эстетичными признаками самцов и их успехом в размножении.
Именно такая скандальная ситуация сложилась сейчас, например, с птицами отряда куриных. В том числе, и со знаменитыми павлинами. Это, конечно, вообще анекдот – когда «икона полового отбора» оказалась не в состоянии подтвердить теорию полового отбора (сразу в трех последних независимых исследованиях). Но как я уже сказал выше, подробный анализ этих исследований – тема для следующей книги.
5. Еще хуже дела становятся, когда «архитектурные излишества» демонстрирует не только самец, но и самка тоже. То есть, сразу оба пола. Такие вещи теория полового отбора объяснить вообще не может. Конечно, выдвигаются определенные предположения, но настолько слабые, что их даже лень критически разбирать.[120]
6. И наконец, совсем плохо становятся дела, когда живое существо демонстрирует явно «радикальную» внешность, совершенно не полезную для выживания… но при этом имеет такие органы чувств, что просто не в состоянии оценить радикальные особенности собственной внешности.
Может быть, Вы помните, как мы разбирали выше «проблему белых бабочек». Для тех, кто уже не помнит, вкратце повторю, о чем речь. На рисунке 48 слева показана ночная бабочка березовая пяденица, которая имеет прекрасную маскирующую окраску, помогающую ей избегать внимания дневных насекомоядных птиц.
А справа мы видим тоже ночную бабочку, и тоже пяденицу. Но эта бабочка хорошо заметна на том фоне, где она «спряталась». Одновременно её внешность очень удачно совпадает с нашими эстетическими предпочтениями. Неслучайно название этой бабочки – пяденица великолепная (Iotaphora admirabilis).
Причем мы видим в её внешности много очень тонких деталей (обратите внимание на «проработку» цветных линий на крыльях), которые сама эта бабочка не в состоянии оценить чисто физически (Рис. 48):
https://goo.gl/f1KgiC
Рисунок 48. Слева березовая пяденица, справа пяденица великолепная. Пояснения в тексте.
Если мы предполагаем, что окраску березовой пяденицы под фон дерева так хорошо подобрал естественный отбор (под давлением хищных птиц), тогда какая сила подбирала наряд пяденицы великолепной?
В следующей книге мы обязательно вернемся к этой теме еще раз. И познакомимся еще с парочкой «открытий чудных» в этой области. Здесь же я приведу только три конкретных примера, которые одновременно соответствуют озвученным выше пунктам 5 и 6 (Рис. 49):

Рисунок 49. Eudaemonia troglophylla. Бабочка из трибы Urotini, подсемейство Saturniinae.[121]
Обладательницей этих поразительно длинных «хвостов» на крыльях является бабочка Eudaemonia troglophylla (подсемейство Saturniinae). Одновременно, сотни видов бабочек из этого же подсемейства вообще не имеют ничего похожего на такие «хвосты». В качестве всего пары наглядных иллюстраций – например, бабочки Antherina suraka и Maltagorea fusicolor, относящиеся к этой же трибе бабочек, не имеют даже намека на какие-либо «хвостики» (Рис. 50):

Рисунок 50: Antherina suraka (самец) и Maltagorea fusicolor. Бабочки из трибы Urotini, подсемейство Saturniinae.[122]
Тем не менее, эти бабочки, вроде бы, не собираются вымирать из-за полного отсутствия каких-либо хвостов на крыльях (как и еще сотни видов бабочек из этого же подсемейства). Следовательно, в данном случае мы вряд ли можем сказать, что гигантские «хвосты» на крыльях бабочки Eudaemonia troglophylla – имеют страшную биологическую важность (для этого вида бабочки). Скорее уж, наоборот, здесь напрашивается вывод о несомненной биологической помехе. Это ведь надо, чтобы так угораздило – сначала отращивать, а затем еще и всё время таскать с собой такой вот дополнительный вес (и объем).
Похоже, это как раз тот случай, как нам надо «доставать из широких штанин» (С) именно версию полового отбора. Иначе объяснить столь радикальные особенности дизайна этого вида бабочки – вряд ли удастся. Но с позиций теории полового отбора, для того чтобы отобрать «хвосты» такой длины (и потом стабильно поддерживать этот признак из поколения в поколение), самки этого вида бабочек должны упрямо предпочитать самцов с более длинными «хвостами», жестоко отказывая самцам с более короткими хвостами. Но как же самки сделают это, если для этого нужно видеть разницу между «хвостами» разных самцов? Если самки не будут видеть тонких различий между длиной «хвостов» у разных самцов, тогда ничего не мешает этим «хвостам» постепенно уменьшиться (до биологически оптимальных размеров) незаметно для самок.
Я уже писал выше, что зрение бабочек не настолько хорошее, чтобы визуально оценивать подробные детали своего возможного полового партнера. Например, длину его «хвостов». Уже на расстоянии в два десятка сантиметров они этого сделать не смогут. А на очень близкой дистанции (в несколько сантиметров) такое «чудо природы», наверное, невозможно окинуть оценивающим взором уже чисто «технически».
Так какая же сила отбирала столь длинные «хвосты» бабочке Eudaemonia troglophylla, при этом не отобрав совершенно никаких хвостов бабочке Maltagorea fusicolor, хотя эти виды бабочек принадлежат к одной и той же трибе?
Причем у многих «длиннохвостых» бабочек хвосты имеют и самец, и самка. То есть, сразу оба пола. Поэтому о половом отборе здесь речи, похоже, вообще нет. А речь здесь идет (давайте честно признаемся) об определенном стиле дизайна, в соответствии с которым был создан данный вид бабочек. И этот стиль дизайна выдерживается и в отношении самца, и в отношении самки. Правда, в отношении самки этот стиль дизайна всё-таки выглядит более умеренным – как правило, у самок подобных бабочек «хвосты» заметно короче, чем у самцов. Оно и понятно – на самку ложится более серьезная нагрузка по жизни – ей надо создать много питательных яиц. Поэтому слишком увлекаться «архитектурными излишествами» самкам нельзя.
И это правило широко распространено в природе. Там, где мы видим у животных «архитектурные излишества», самки носят такие излишества в более умеренном варианте (если вообще носят). Помните, выше мы разбирали жирафов? Там самки тоже имели более «здравые» размеры тела, чем самцы. И соответственно, выживали, действительно, лучше (см. выше).
То есть, здесь всё продумано – стили дизайна разных живых существ могут быть поразительными и даже радикальными, но в таком случае самки демонстрируют эти стили дизайна в «щадящем режиме». А у многих видов «архитектурные излишества» демонстрируют вообще только самцы. В рамках концепции разумного дизайна, здесь действительно всё разумно. А именно, и одна из основных целей дизайна достигается – создание максимального разнообразия форм жизни на Земле. И в то же время, конкретные биологические виды не поставлены на грань вымирания из-за слишком накладных «архитектурных излишеств». Наиболее накладные «архитектурные излишества» вообще несут только самцы. Самки же полностью высвобождаются для успешного продолжения рода. Например, у тех же индийских павлинов самки в период высиживания яиц и выращивания птенцов гибнут от хищников гораздо чаще, чем самцы (Takahashi et al., 2008), несмотря на то, что вообще не имеют никаких «украшений». А имеют скромную окраску, выполняющую классические функции маскировки. Понятно, что тут уже не до «архитектурных излишеств».
Но наивные дарвинисты, не понимая истинных причин красивой окраски самца павлина, объявляют индийских павлинов… классическим примером полового отбора. Они заявляют, что павлин такой красивый потому, что это его так отобрала павлиниха (так эстетично). Здесь перепутано вообще всё, что только можно, и поставлено с ног на голову. Более того, в качестве гениев эстетического дизайна выступают… курицы.[123]
К счастью, последние полевые исследования успешно провалили теорию полового отбора именно в отношении павлинов. И теперь верующим дарвинистам стало совсем плохо. Теперь им приходится рассуждать о воображаемых прошлых половых предпочтениях самок павлинов, которые уже успели измениться (Takahashi et al., 2008). То есть, соответствующие предпочтения самок «уже улетели, но обещали вернуться» (С). Как в том мультфильме про неуловимого Карлсона. И поэтому никаких связей между размером «хвоста» павлина и успехом его размножения никак не удаётся обнаружить (Loyau et al., 2005; Takahashi et al., 2008; Dakin et Montgomerie, 2011). Эту поучительную историю мы тоже подробно разберем в следующей книге. А пока второй пример «архитектурных излишеств» (Рис. 51):

Рисунок 51. Жук арлекин (Acrocinus longimanus). Вверху самка, внизу самец.[124]
Это знаменитый жук-арлекин (Acrocinus longimanus). Как видим, здесь всё повторяется снова. Опять «архитектурные излишества» – непропорционально длинные передние лапы, которыми этот жук, собственно, и выделяется среди всех своих коллег – сотен тысяч (!) других видов жуков. И опять видим, что и самка (вверху) «выполнена» в соответствующем стиле дизайна, схожим с дизайном самца. Но одновременно хорошо видно, что дизайн самки гораздо более скромный (более благоразумный), чем «буйно-фантастический» дизайн самца. И снова не очень понятно, для чего же это «архитектурное излишество» вообще нужно.
Была идея, что такие длинные лапы нужны самцу этого вида для того, чтобы бороться за самку (и чтобы охранять те места, где самка откладывает яйца). Поскольку самцы этих жуков борются между собой, в том числе, и с помощью передних лап. Кроме того, самцы еще и кусают друг друга, и бодаются головами (Zeh et al., 1992). Однако проведенные исследования показали, что успех в этой борьбе способствует тем самцам, которые просто имеют большие размеры тела (Zeh et al., 1992). Какую-то особую роль именно длинных передних лап в поединках самцов выявить не удалось. В связи с этим, авторы исследования написали, что «фактор длины ног» маскируется корреляцией с общим размером тела (Zeh et al., 1992). Действительно, если большие размеры тела благоприятствуют победе, а длина передних ног, в свою очередь, коррелирует с размером тела, то длина передних ног тоже будет коррелировать с успехом. Вот только здесь получается такая «связь», которая аналогична «успеху боксеров в красных трусах» – если красные трусы надеть на боксеров весом 100 кг, а синие трусы – на боксеров весом 50 кг, то в поединках между этими боксерами чаще станут побеждать боксеры в красных трусах. Следовательно, именно красные трусы помогают побеждать в боксерских матчах?
Ну а если серьезно, то понятно, что бороться с другими самцами, имея такие «ходули» – вряд ли более удобно, чем имея «среднестатистические» ноги, как у сотен тысяч других видов жуков (в том числе, и у тех, кто любит бороться между собой). Здесь преимущество даёт общий размер тела, а не длина передних ног, тем более, такая «запредельная». В конце концов, если уж передние лапы так важны этим жукам для борьбы друг с другом, то в ходе эволюции следовало бы ожидать появления чего-то похожего на передние лапы богомола. Вот передние лапы богомола – это, действительно, крайне эффективное оружие. Но никак не те «ходули», которые имеются у жуков-арлекинов. Похоже, столь удлиненные передние лапы жукам-арлекинам вообще не нужны (ни для чего биологически полезного). Иначе из почти 400 тысяч (!) видов жуков мы непременно нашли бы еще тысяч десять аналогичных примеров. Вместо этого имеется лишь несколько видов жуков с длиннющими передними лапами на всё огромное «жучиное царство».[125]
В общем, самая правдоподобная и простая версия в данном случае напрашивается сама собой – у жуков-арлекинов просто такой уникальный стиль дизайна.
И уникальных стилей дизайна в природе великое множество. Одни только «стебельчатоглазые» мухи чего стоят (Рис 52):
https://goo.gl/xgl5zn
Рисунок 52. Разные представители «стебельчатоглазых» мух. Да-да, не удивляйтесь, глаза всех этих мух находятся именно на концах длинных стебельков. Хотя в случае верхней мухи слева (Plagiocephalus latifrons) это уже больше похоже на огромные рога.
Остаётся только удивляться, как эти мухи всё еще умудряются летать.
Причем этот уникальный стиль дизайна характерен для многих видов мух сразу из нескольких семейств. А вот степень выраженности этого стиля различается даже среди близких биологических видов (Рис. 53):
https://goo.gl/Tc37ww
Рисунок 53. 1,2,3 – самцы разных видов рода Plagiocephalus: 1 – P. latifrons; 2 – P. lobularis; 3 – P. intermedius. 4,5 – головы самок: 4 – P. latifrons; 5 – P. intermedius. Изображения взяты из работы (Kameneva, 2004).
То есть, мы можем наблюдать в природе такие радикальные стили дизайна, в которых «выполнены» целые таксономические группы живых существ (вплоть до уровня семейств). Несмотря на отсутствие биологической необходимости в соблюдении подобного стиля. А внутри этого общего стиля дизайна, в свою очередь, наблюдается разнообразие его вариантов.
Кстати, в приведенном случае самки вообще не отягощены соответствующими «архитектурными излишествами» (см. рис. 53, изображения 4,5).
В тех же случаях, когда самки разделяют сходный стиль дизайна вместе со своими самцами, это обычно опять происходит в более «скромном» (биологически более благоразумном) варианте, чем у самцов (Рис. 54):
https://goo.gl/PkN2OR
Рисунок 54. Спаривающиеся стебельчатоглазые мухи. Хорошо видно, что у данного биологического вида стебельчатоглазых мух: 1) самка имеет стиль дизайна, сходный со стилем дизайна самца, 2) но в более «скромном» (биологически более благоразумном) варианте.
Впрочем, среди стебельчатоглазых мух есть и такие виды, у которых и самцы и самки имеют «архитектурные излишества», развитые в совершенно одинаковой степени! Однако у таких видов, в целом, гипертрофия обычно выражена скромнее, чем у тех видов, где самцы по этому признаку отличаются от самок (Рис. 55):
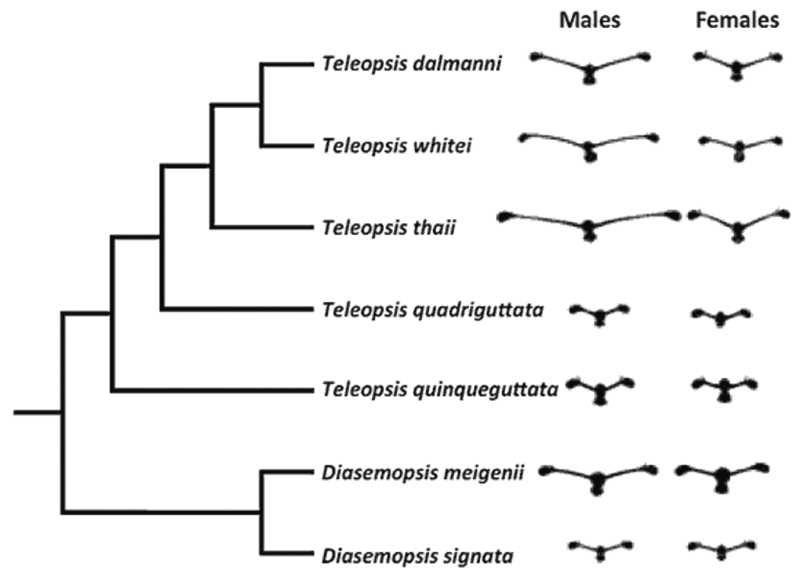
Рисунок 55. Разные виды и роды стебельчатоглазых мух (с предполагаемыми филогенетическими связями). Слева силуэт головы самцов разных видов, справа – самок. Рисунок взят из работы (Husak et al., 2011).
Из рисунка хорошо видно, что стебельчатоглазые мухи представлены как мономорфными видами, то есть такими, где морфология самца и самки вообще не различается. Так и диморфными видами, у которых имеется половой диморфизм по обсуждаемому признаку. Заметно, что в случае мономорфных видов, гипертрофия «стебельков», в целом, выражена слабее, чем в случае диморфных видов. В свою очередь, у самок диморфных видов гипертрофия признака всегда выражена скромнее, чем у самца.
И вот нам предлагается поверить, что этот радикальный стиль дизайна, в котором «выполнены» целые биологические роды в разных семействах мух (данный стиль дизайна повторяется еще и в разных семействах)… так получилось исключительно потому, что самки мух упорно предпочитали самцов с самыми широко расставленными глазами. И в результате этих, поразительно устойчивых половых предпочтений (не изменявшихся на протяжении долгих миллионов лет!) образовались целые роды мух с видами, поголовно обладающими именно данным стилем внешности (в той или иной степени). Получается, что в рамках сразу нескольких биологических родов, имела место упорная «эволюционная движуха» в строго определенном направлении, да еще и вопреки давлению обычного естественного отбора. Ведь никто, надеюсь, не будет спорить, что мухам со «среднестатистической» головой выживать всё-таки легче, чем с такой головой, которую имеют стебельчатоглазые мухи.
Интересно, что имеется ряд публикаций, в которых версия полового отбора у стебельчато-глазых мух… получила подтверждение. Будто бы самки стебельчатоглазых мух, действительно, предпочитают самцов с самыми широко расставленными глазами. И самцы с такими глазами, будто бы, тоже имеют преимущество в поединках с другими самцами.[126]
Такое явление я называю «телекинезом в биологии». Когда очень хочется, чтобы результаты твоих собственных исследований сошлись с прогнозами господствующей теории, то (о чудо!) результаты действительно могут сойтись с тем, с чем тебе очень хотелось, несмотря на реальное положение дел в данной области. Видимо, сила мысли некоторых исследователей является настолько мощной, что сама реальность начинает искажаться, подстраиваясь под теоретические ожидания этих ученых. Такое явление я и называю «телекинезом в биологии». Особенно легко «телекинез в биологии» может происходить там, где исследования связаны с выявлением каких-нибудь статистических закономерностей, то есть там, где несколько лишних точек на графике способны изменить результат работы с формулировки «ожидаемой связи не выявлено» до формулировки «ожидаемая связь установлена».
К сожалению, у научных результатов, добытых таким «телекинетическим способом», обычно начинаются проблемы с воспроизводимостью, как только другие исследователи начинают проводить независимые исследования. Видимо, другие исследователи уже не обладают достаточной силой мысли, чтобы влиять на реальность правильным образом. Поэтому они и не могут воспроизвести результаты, добытые с помощью «телекинеза в биологии». Таким образом, «телекинетические» результаты обычно живут в науке только до того момента, пока не будут проведены другие (независимые) исследования в этой же области.
К сожалению, независимых специалистов часто не хватает для того, чтобы проводить повторные исследования одних и тех же биологических объектов. Поэтому иногда «телекинетические» результаты в биологии могут спокойно «жить» годами. И даже попадать в соответствующие учебники в качестве «доказанных фактов». В итоге складывается ситуация, когда одни «телекинетические» результаты опровергаются независимыми исследованиями. А другие всё еще «живут», дожидаясь своего опровержения. Например, «обнаруженные» в ранних исследованиях половые предпочтения пав (самок павлинов) к самым большим хвостам своих кавалеров – уже опровергнуты в более поздних работах. А вот «половые предпочтения стебельчатоглазых мух», вроде бы, пока еще только дожидаются своего «разрушителя мифов».
Впрочем, некоторые авторы, стараясь подтвердить половой отбор слишком рьяно, иногда неправильно понимают «линию партии»… и в результате получают такие результаты, которые, наоборот, опровергают версию про половой отбор. Например, в работе (Wilkinson & Reillo, 1994) авторы установили, что если начать искусственно отбирать линию мух с коротким расстоянием между глазами, то уже через 13 поколений такой селекции, самки этой линии начинают предпочитать самцов с коротким расстоянием между глазами! То есть получается, что самки всего за 13 поколений самостоятельно изменили свои исходные половые предпочтения не просто на другие, а вообще на противоположные.
По сути, получилось, что половые предпочтения самок стебельчатоглазых мух просто пассивно следуют за тем направлением отбора, которое имеется в данный момент, не демонстрируя никакой устойчивости. Понятно, что такой результат сразу же обесценивает версию о половом отборе, который якобы упорно происходил в строго определенном направлении в рамках целого биологического семейства, на протяжении долгих миллионов лет. Ведь если половые предпочтения самок настолько неустойчивы, что могут изменяться (вообще на противоположные) за 13 поколений, то ни о каком формировании (под действием столь изменчивых половых предпочтений) единого морфологического облика сразу у множества биологических видов (относящихся к целому семейству мух) – не может быть и речи.
В заключение давайте еще раз подчеркнем общую иронию ситуации в случае со стебельчатоглазыми мухами:
1. Стебельчатоглазые мухи имеют явно радикальную внешность.
2. В рамках дарвинизма, эта внешность объясняется тем, что, во-первых, самки этих мух предпочитают самцов с самыми широко расставленными глазами. А во-вторых, самцы с широко расставленными глазами имеют преимущество в поединках с другими самцами. Таким образом, обсуждаемый признак связан исключительно с отбором самцов.
3. Тем не менее, самки стебельчатоглазых мух этого рода – тоже стебельчатоглазы.
4. Более того, чуть ли не у половины представленных видов, самки «стебельчатоглазы» совершенно в той же степени, что и самцы (см. рис. 55).
5. Причем этот поразительный стиль дизайна наблюдается практически поголовно в рамках целого биологического семейства (и не только).
Так какое же объяснение причин подобной внешности (у целых биологических семейств, у самцов и самок (!) тех видов, которые принадлежат к этой группе) звучит более правдоподобно? Что это результат полового отбора самцов? Или, что это такой уникальный стиль дизайна, характерный для соответствующих биологических таксонов?
Итак (заканчиваем), теория разумного дизайна объясняет факты существования подобных «архитектурных излишеств» в живой природе, в целом, более правдоподобно и намного проще, чем дарвиновская теория. А именно, разнообразием стилей дизайна живых существ. Где создаваемое разнообразие, скорее всего, не преследовало никаких других целей, кроме, собственно, самого биологического разнообразия.
Ну а подробно на эту тему мы еще поговорим позже, в следующей книге.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК