8. Сочиняем альтернативную теорию биологии
… В принципе, я категорически против танцев. Но если она хочет, отказывать нельзя…
(Из художественного фильма «Hitch», 2005)
В принципе, я категорически против выдумывания каких бы то ни было сценариев о том, как разворачивались биологические события даже 20.000 лет назад, не говоря уже о миллионах и миллиардах лет. Потому что с очень большой вероятностью это окажутся чистые спекуляции, не соответствующие тому, что там было на самом деле. Но если дарвинист просит выдумать какую-нибудь теорию – отказывать нельзя.
Многие верующие дарвинисты почему-то убеждены, что нельзя ограничиваться одной лишь критикой теории, даже если эта теория явно ошибочна. Нужно обязательно предложить альтернативу. И если альтернатива не предложена, то лучше уж ошибочная теория, чем никакая. Что и говорить, это весьма удивительная позиция. На мой взгляд, всё обстоит с точностью до наоборот. То есть, уж лучше никакой теории, чем ошибочная.
Допустим, лично я не знаю, каким образом установить на Земле всеобщий мир и справедливость. А вот кто-то утверждает, что мира и справедливости можно достичь, используя идеи социального дарвинизма. Или даже расизма, который тоже легко выводится из дарвиновских идей, примененных в отношении человечества. Действительно, если человечество эволюционировало из обезьян, в ходе длительного эволюционного процесса, то получается, что одни расы или народы неизбежно продвинулись в этом отношении дальше, чем другие (хотя бы в какой-то мере). В таком случае, наверное, мы должны сами помочь эволюции, дав возможность жить и оставлять детей только самым эволюционно продвинутым расам, народам или кастам? И когда именно такой порядок будет установлен, тогда и наступит всеобщая гармония и полная эволюционная справедливость.
И вот, допустим, я твердо уверен, что этот «кто-то» абсолютно не прав. То есть, я считаю, что идеи социал-дарвинизма и расизма несут страшные угрозы именно миру и справедливости. Но сам я не знаю, как же всё-таки можно достичь всеобщего мира и справедливости (каким-нибудь другим способом). Разве отсюда следует, что пока я этого не узнаю, лучше будет придерживаться идей социал-дарвинизма или расизма?
Понятно, что идея дарвиновской эволюции – не столь откровенно чудовищна, как, например, идеи расизма. Всё-таки здесь речь идет, во-первых, о далеком прошлом, которое уже не изменить, как бы ошибочно мы его себе не представляли. А во-вторых, речь, как правило, идёт о прошлом других биологических таксонов, каких-нибудь червяков или крокодилов, но не человека. Лишь иногда верующие дарвинисты шокируют нас рассуждениями о том, как «проституция сделала из обезьяны человека» (см. выше), или какими-нибудь другими подобными гадостями, высосанными из пальца. Тем не менее, дарвиновская теория всё-таки не такая безвредная, какой хочет казаться. Трудно оценить, сколько юношей и девушек, прочитав в школьном учебнике про «главный принцип живой природы» – выживание сильнейшего – перенесли этот принцип еще и на взаимоотношения между людьми, и начали его последовательно применять, постепенно к нему привыкая.
Итак, всё-таки, наверное, лучше не иметь вообще никакой теории, чем иметь теорию ошибочную. Но убедить в этом многих верующих дарвинистов крайне сложно. И причины этого достаточно понятны. Дарвинисты искренне убеждены, что придумать какую-нибудь альтернативу дарвиновской эволюции – практически невозможно. Поэтому они думают, что если поставить такую задачу креационисту, тот сразу же впадёт в ступор, и таким образом, проиграет дискуссию. А дарвинист сможет потом глубокомысленно порассуждать о том, что «креационисты ничего не делают для науки, а только паразитируют на её достижениях, критикуя слабые места научных теорий».
Чтобы не доставлять дарвинисту такого удовольствия – отказывать ему нельзя. Если дарвинист хочет, чтобы ему предоставили альтернативную научную теорию, то нужно предоставить ему альтернативную научную теорию.
Поэтому я сейчас набросаю примерную схему такой (альтернативной) теории.
Понятно (еще раз), что это будут, в основном, спекуляции, и это весьма удручает. Нас будет утешать лишь то, что спекуляций здесь будет всё же меньше, чем в вольных рассуждениях дарвинистов на тему «кто выжил, тот и есть самый приспособленный». Или про то, как «одни биологические таксоны эволюционировали, потому что имели такую возможность, а другие биологические таксоны не эволюционировали, потому что не имели такой возможности»…
И уж конечно, в предложенной нами теории не будет хотя бы откровенных глупостей. В отличие, например, от абсолютно диких (с химической точки зрения) сказках про абиогенез.[63]
Как известно, имеющиеся на сегодняшний день факты лучше всего вписываются в рамки так называемой «концепции непрерывного творения». Понятно, что это тоже не самая лучшая концепция, поскольку всё-таки не все известные факты с ней совместимы. Тем не менее, это лучшее, что сегодня можно предложить в этой области. Любым другим биологическим теориям противоречит гораздо большее число фактов.
Итак, в рамках «концепции непрерывного творения», можно предположить, что какая-то разумная сила в ходе истории Земли периодически создавала всё новые таксоны живых существ, с характерными для этих таксонов признаками (и соответственно, генофондами).
Мы не знаем, каким конкретно образом это происходило. Можно предположить, что это происходило либо в результате создания с нуля («вброса» новых биологических таксонов), либо путем генетической модификации, то есть, создания новых биологических таксонов из уже имевшихся (старых) биологических таксонов.
Такое предположение немного напоминает идею Михаила Шермана об «универсальном геноме» (Sherman, 2007). Только у Шермана происходит одноразовый гипотетический вброс в биосферу «универсального генома всех многоклеточных животных» (в кембрийском периоде), а мы можем предположить, что такие вбросы происходили в истории Земли неоднократно: 1) время от времени, 2) периодически, 3) или даже сравнительно постоянно. И ранги создающихся новых таксонов могли варьировать от сравнительно высоких (например, типы, классы, отряды) до самых низких (семейства, роды или даже крупные виды).
Повторю еще раз пару абзацев, которые я уже писал выше по этому поводу:
…Эта (некая) сила могла взять, и смести все «фигуры» с шахматной доски (каким-нибудь глобальным катаклизмом, приводящим к массовому вымиранию). И начать новую «партию», создавая новые биологические таксоны целыми «пачками». Именно такой взгляд на эволюцию озвучивался в знаменитой теории катастроф, выдвинутой Жоржем Кювье и развитой его последователями. Именно так эта теория объясняла массовые вымирания целых (ныне ископаемых) фаун и флор и последующее становление новых.
Однако глобальные катастрофы в рамках концепции непрерывного творения совсем не обязательны. Создающая сила вполне могла заниматься и «штучной работой». То есть, периодически то один новый таксон создаст, то другой. Причем или мгновенно создаст (допустим, из какого-нибудь предыдущего таксона), или может долго «вытачивать» характерные признаки этого таксона. А палеонтологи потом обнаруживают и прослеживают в ископаемых останках соответствующие «эволюционные тенденции» и «явно направленную изменчивость» (см. выше). И ломают себе голову, что же эти (столь упорные) «эволюционные тенденции» могли означать?
Здесь хороша аналогия с тем, как трудится увлеченный садовник в своем саду. Он то копается там целыми днями напролет, что-то создавая и сильно переделывая. А то просто сидит и ничего не делает – просто любуется результатом. Или же периодически подправляет какие-то мелочи. Или, возможно, уделяет внимание какой-то отдельной посадке и т. п.
Вот такую модель развития живой природы и можно назвать «моделью непрерывного творения».
Продолжаем спекулировать.
Можно предположить, что первые представители «вброшенных» таксонов имели богатые, многовариантные генофонды, которые далее уже самостоятельно дробились. То есть, исходный многовариантный генофонд нового таксона сравнительно быстро разбивался на всё более «узкие» геномы, при этом порождая видовое разнообразие нового таксона. Отметим, что в палеонтологической летописи такие вещи потом будут выглядеть как быстрый «эволюционный расцвет таксона».
Впрочем, во многих случаях правильней будет предположить не самостоятельное дробление таксона, а тоже направляемое (контролируемое) какой-то разумной силой. Иначе затруднительно объяснить факты существования (у разных биологических видов) многих признаков, биологическое значение которых незначительно (или вообще непонятно), следовательно, не могло быть отобрано естественным отбором, но, тем не менее, это признаки «проработаны» настолько совершенно, что роль прямого разумного дизайна здесь достаточно очевидна. В первую очередь, это касается «эстетических» признаков большого круга живых существ.
Например, выше я уже упоминал птиц-шалашников, в том числе, разнообразие их брачных беседок (от вида к виду). Я предположил, что это разнообразие могло возникнуть вследствие самостоятельного (автоматического) «дрейфа» вокруг некоего (исходно запрограммированного в генофонде) паттерна «строительства брачных беседок». Но ничего не мешает предположить, что даже такого автоматического дрейфа в данном случае тоже не было. А было именно «поштучное» создание биологического разнообразия этого таксона. За счет направленных генетических модификаций исходного паттерна получилось наблюдаемое разнообразие уже внутри рамок этого таксона.
Под давлением ряда фактов, в рамках концепции непрерывного творения (биологических таксонов) вполне можно предположить не только запрограммированные «эволюционные расцветы таксонов», но и их закономерные угасания. Например, по следующему гипотетическому сценарию.
Постепенно, дробление нового таксона заходило всё дальше, порождая всё более мелкие «виды». Фаза «эволюционного расцвета» таксона сменялась фазой его эволюционного угасания. Возможные биологически целесообразные варианты геномов и морфотипов – исчерпывались, но геномы продолжали дробиться на все более мелкие группы, под действием чисто случайных (энтропийных) сил. Например, и так уже ставшие «мелкими» биологические виды, в результате случайных хромосомных перестроек могли дробиться еще более мелко, порождая «виды-двойники» или даже отдельные изолированные популяции, генетически несовместимые друг с другом. К такому же «измельчанию» могли приводить и другие виды изоляции популяций. Понятно, что у «мелкого вида» шансы вымереть (от любых причин) гораздо выше, чем у крупного.
Можно предположить, что генофонды биологических видов стареющего таксона постепенно обеднялись и деградировали еще и за счет дрейфа генов. А также из-за прямой деградации отдельных генов, вследствие постепенного накопления в этих генах слабо-вредных мутаций, которые очищающий отбор не замечает.[64]
Понятно, что такие мелкие, да еще и деградировавшие, «осколки» прежде полноценных и «генетически здоровых» таксонов имели гораздо более высокие риски вымирания, чем первоначальные таксоны. Вот они и вымирали.
Получается, что каждый новый созданный таксон имеет определенное «время жизни». А так же собственный «онтогенез» – рождение (создание), расцвет, старение (дробление и деградация) и вымирание.
В связи с этим можно даже предположить, что периодический «вброс» в биосферу новых, свежих таксонов происходил не произвольно (по желанию), а это было вообще необходимо, чтобы жизнь на Земле не угасала вследствие постепенного вырождения.
Как уже говорилось, этот «вброс» мог осуществляться тремя возможными способами:
1) Созданием новых таксонов «с нуля».
2) Созданием новых таксонов с использованием некоторых предыдущих «наработок».
И наконец:
3) Путем прямой генетической модификации предыдущих (стареющих) таксонов – их исправлением, «ремонтом» и соответствующими изменениями, необходимыми для нового «стиля» таксона (то есть, нового таксона, с новым стилем дизайна, на основе старого таксона).
Итак, вполне можно представить себе развитие жизни на Земле, как смену создаваемых биологических таксонов, с разными стилями дизайна, последовательно населявших нашу планету. Причем эта смена могла производиться как «точечно» (в рамках отдельных таксонов низкого ранга), так и глобально, сразу целыми биомами.[65] Действительно, вряд ли кто-то будет спорить, что дизайн каких-нибудь «динозавровых ландшафтов» резко отличается от дизайна «птице-млекопитающих ландшафтов» именно по своему стилю (в том числе, эстетическому).
Можно предположить, что такая (периодическая) смена стилей дизайна живых существ происходила без какой-либо особой необходимости. Просто вследствие свободного волеизъявления некоей создающей силы. Например, в целях разнообразия. Или можно предположить, что это делалось вследствие необходимости – замены стареющих биологических таксонов новыми таксонами, молодыми и «полными жизни». Или же справедливо и первое и второе – то есть, таксоны заменялись по необходимости, но стиль большинства заменяемых таксонов не повторялся уже в целях разнообразия.
Хотя в рамках концепции непрерывного творения, в периодических повторениях того или иного стиля дизайна тоже нет ничего удивительного. Современная теория эволюции такие вещи называет конвергенцией, и нередко такая конвергенция плохо объяснима с позиций чисто биологической целесообразности. Например, не очень понятно, зачем так нужна была именно «саблезубость»: 1) хищным зверозубым терапсидам; 2) некоторым травоядным динозаврам; 3) травоядным ископаемым млекопитающим диноцератам (при одновременной редукции верхних резцов); 4) некоторым линиям хищных ископаемых млекопитающих креодонтов (Machaeroides); 5) хищным сумчатым саблезубым тиграм (Thylacosmilus); 6) сразу нескольким линиям хищных плацентарных саблезубых кошек; 7) современным травоядным млекопитающим – мускусному оленю кабарге и водяному оленю (Рис. 28):

Рисунок 28. Саблезубые животные. Вверху слева – череп ископаемого сумчатого саблезубого тигра. Вверху по центру – череп ископаемого плацентарного саблезубого тигра. Внизу слева – череп современного травоядного мускусного оленя кабарги (Moschus moschiferus). Справа – фотография живого самца кабарги.[66]
Особенно если учитывать тот факт, что все современные наземные хищные млекопитающие, представители семейств: псовых, кошачьих, куньих, медвежьих, гиеновых, виверровых, енотовых и др. (включая и сумчатых хищников) – все эти животные сегодня имеют клыки гораздо меньших размеров, чем озвученные выше саблезубые животные.[67] Даже зубы разнообразных хищных рептилий и подавляющего числа хищных рыб – тоже не имеют такой длины, какую демонстрируют нам саблезубые животные. Все эти многочисленные примеры прозрачно намекают, что если бы саблезубые животные не выделывались, а имели зубы, аналогичные по размеру зубам всех существующих сегодня хищных животных,[68] то такие зубы, скорее всего, являлись бы более эффективным средством для нанесения травм добыче (или друг другу).[69]
В общем (еще раз) с позиций чисто биологической целесообразности, «саблезубые» клыки не очень понятны.
А вот в рамках представлений о стилях дизайна нет ничего удивительного в том, что отдельные (популярные?) стили дизайна могут повторяться в разных биологических таксонах (в том числе, и в самых неожиданных, например, у травоядных оленей). Несмотря на сравнительно меньшую эффективность подобных зубов в сравнении с «классическими».
Таким образом, в рамках концепции разумного дизайна, конвергенцию форм можно объяснять просто повторением определенного стиля дизайна. В то время как в рамках дарвинизма эту же конвергенцию приходится объяснять результатом эволюции в сходных условиях. Однако в некоторых наиболее тяжелых случаях (удивительно точной конвергенции) заранее ясно, что такого объяснения совершенно не достаточно.
Действительно, трудно представить, чтобы совершенно случайная эволюция, слепо «бродя» («методом случайного тыка») по практически безграничному пространству эволюционных возможностей… вдруг (чисто случайно) облекала два разных существа в практически одинаковую форму.
Самым известным примером такой потрясающей «конвергенции форм» является, наверное, знаменитый сумчатый волк в сравнении с волком обыкновенным. Можно сравнить между собой их черепа (Рис. 29):
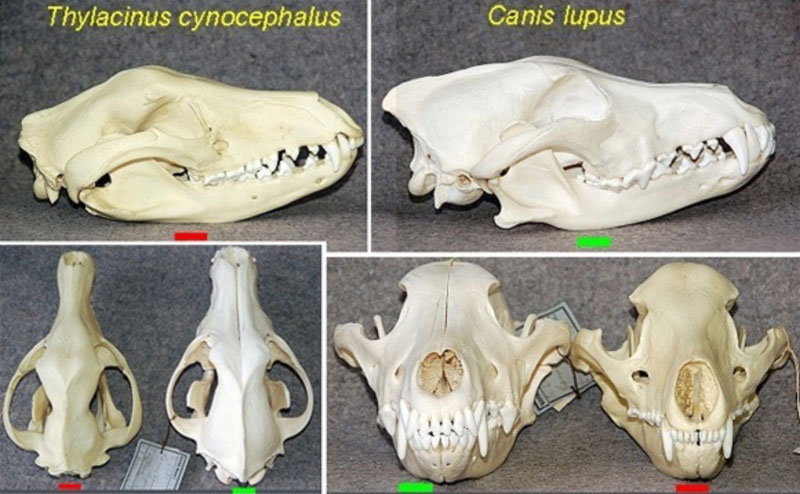
Рисунок 29. Сравнение черепа обыкновенного волка Cani's lupus (зеленые прямоугольники) с черепом сумчатого волка Thylacinus cynocephalus (красные прямоугольники).[70]
Очевидно, что сходство весьма впечатляет. Особенно бросается в глаза общее морфологическое сходство зубного ряда этих двух совершенно разных зверей. А ведь согласно современным представлениям, эволюционные линии сумчатых и плацентарных млекопитающих разделились друг от друга, как минимум, 160.000.000 лет назад.
И вот, спустя 160 млн. лет «случайного эволюционного поиска» в безграничном пространстве возможностей, эта «эволюция»… не нашла ничего более оригинального, чем повторить одну и ту же форму почти буквально![71]
Но может быть, «пространство эволюционных возможностей», на самом деле, совсем не безгранично? А наоборот, очень даже ограниченно? Допустим, именно такая форма черепа является самой эффективной для животного, ведущего хищный образ жизни? Вот поэтому естественный отбор и позаботился о том, чтобы череда совершенно случайных мутаций в конечном итоге «выстроилась» в почти одинаковые черепа?
Что же, давайте попробуем оценить, ограничено ли «пространство эволюционных возможностей» (для черепов хищных зверей), или же всё-таки не ограничено?
Для этого достаточно посмотреть на зубы другого (вымершего) сумчатого хищника – сумчатого льва (Рис. 30):
https://qoo.ql/HlXWOe
Рисунок 30. Слева одна из реконструкций головы сумчатого льва (Thylacoleo). Справа вверху его череп в профиль. Справа внизу череп (со скелетом) в анфас.
Итак, мы видим, что зубной ряд сумчатого льва радикальным образом отличается не только от зубов волка, но и вообще от зубов любого другого хищного млекопитающего. В первую очередь бросаются в глаза два единственных резца спереди. Что совершенно не характерно ни для одного современного представителя отряда хищных (зато немного напоминает грызунов). И конечно же, привлекают внимание абсолютно необычные, огромные хищные зубы в верхней и нижней челюсти сумчатого льва, которые наводят на стойкие ассоциации с режущими ножницами.
Сегодня считается, что зубы сумчатого льва были весьма эффективны для убийства жертв (Wroe et al., 2005). Тем не менее, эти зубы чрезвычайно сильно отличаются от зубов всех известных хищников. Этот факт наглядно показывает, что пространство эволюционных возможностей всё-таки может быть практически безграничным (при желании).
Тем не менее, мы имеем то, что имеем – поразительное сходство сумчатого волка с волком обыкновенным (вообще практически не родственников)… при разительном отличии сумчатого волка от сумчатого льва (эволюционных родственников).
Такие факты, безусловно, делают тезис о случайном характере эволюции очень уязвимым. Заставляя задумываться о существовании каких-то других формообразующих сил, не имеющих никакого отношения к естественному отбору более приспособленных.
А теперь давайте проведем общий анализ достоинств и недостатков теории непрерывного творения в сравнении дарвиновской теорией эволюции.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК