Гормоны коры надпочечников
Гормоны коры надпочечников
В экспериментах на животных не было обнаружено отчетливого поведенческого эффекта кортикостероидов (т. е. глюкокортикоидов и минералокортикоидов), в противоположность другим гормонам гипофиз-адреналовой системы. В частности, ни скорость обучения, ни память не изменяются после удаления коры надпочечников или введения кортикостероидов так сильно, как это происходит после манипуляций с уровнем других гормонов гипофиз-адреналовой системы (см. ниже). По всей вероятности, это связано с тем, что глюкокортикоиды тормозят по механизму обратной связи секрецию таких мощных психотропов, как кортиколиберин и АКТГ, а также влияют на секрецию многих других гормонов. Поскольку у человека и животных разных психологических типов активность отдельных звеньев гипофиз-адреналовой системы различна (см. главу 6), эффект введения глюкокортикоидов может быть противоположным, и при усреднении результата по группе испытуемых суммарный эффект оказывается нулевым. Например, на рис. 4.22 показано противоположное влияние кортизола на поведение крыс с различным генотипом.
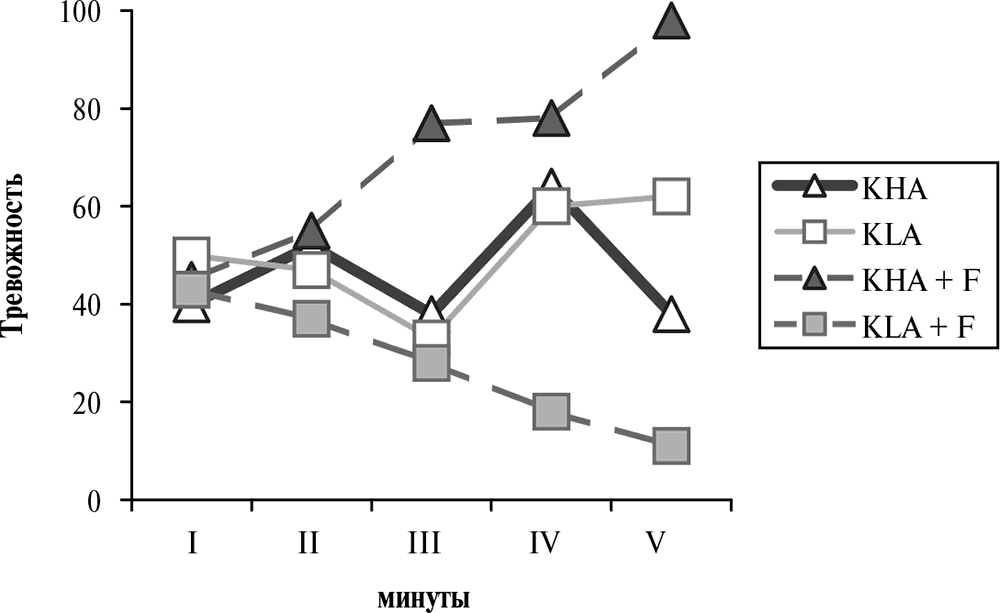
Рис. 4.22. Тревожность у крыс двух генетических линий после введения кортизола[121]. По оси абсцисс – минуты (общее время теста – 5 минут). По оси ординат – время, проведенное животными в открытых освещенных пространствах экспериментальной камеры, для каждой минуты теста. KHA и KLA – названия линий крыс. Пустые значки – группы животных, без введения гормона. Черные значки – группы животных, которым за три дня до теста вводили кортизол (F) в дозе 50 мг/кг. На момент тестирования уровень кортикостерона у этих животных был почти нулевым вследствие торможения секреции глюкокортикоидов по механизму отрицательной обратной связи
Разнонаправленное изменение тревожности у крыс двух групп вызвано, вероятно, торможением различных звеньев гипофиз-адреналовой системы, например кортиколиберина и АКТГ – гормонов, которые оказывают разное влияние на поведение. Поскольку у крыс одной линии тревожность повышалась, а у крыс другой – понижалась, очевидно, что в группе, составленной из представителей обеих линий, эффект кортизола отсутствовал бы.
Только два психотропных эффекта из многих описанных в литературе являются первичными для глюкокортикоидов, а не связаны с их влиянием на другие железы. При лечении большими дозами глюкокортикоидов у большинства больных отмечается эйфория (беспричинно повышенное настроение), доходящая порой до степени психоза. Это так называемые кортизоловые психозы.
Основная психотропная функция глюкокортикоидов – обеспечение реакции затаивания
Вторая первичная психотропная функция глюкокортикоидов – обеспечение реакции затаивания. При удалении коры надпочечников животные постоянно находятся в движении, исчезает реакция затаивания (одна из двух основных форм стрессорной реакции), а при введении глюкокортикоидов в компенсирующей дозе она восстанавливается. Поскольку увеличение дозы вводимого гормона не ведет к увеличению времени, которое животное проводит в неподвижности, глюкокортикоиды не модулируют (не стимулируют), а обеспечивают реакцию затаивания (см. главу 1).
В последние годы интенсивно изучается эффект влияния глюкокортикоидов на гибель нейронов в некоторых структурах головного мозга, в особенности гиппокампа. В малых концентрациях глюкокортикоиды предотвращают гибель нейронов, а в больших, возникающих при длительном стрессе, вызывают клеточную смерть. Поскольку гиппокамп – структура, принимающая участие в организации процессов памяти, аффективного состояния, мотивации, адаптации к изменениям во внешней среде и многих других, значение этого эффекта глюкокортикоидов весьма велико.
Рассмотрим один пример сложной интерпретации результатов психоэндокринного эксперимента[122] (рис. 4.23).
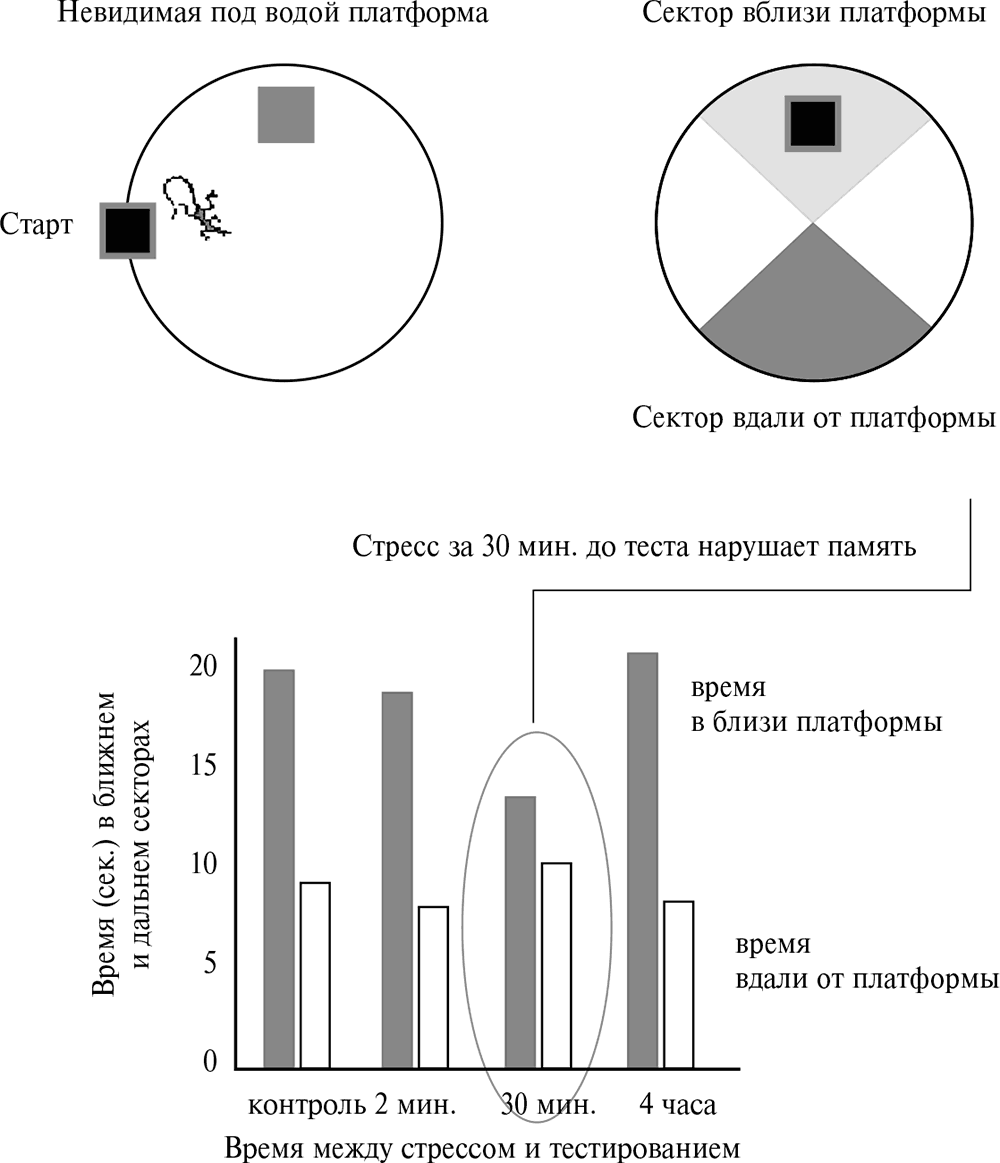
Рис. 4.23. Влияние стресса на память. На верхней панели слева схема лабиринта Морриса – бассейн, куда помещают крысу, которая должна найти платформу, расположенную ниже уровня воды и поэтому невидимую для нее. По тому, как изменяется время, затраченное на поиски при последовательном тестировании, судят о функциях памяти. Количественно такая функция отражается в соотношении времени, проводимом животным в секторе, прилегающем к платформе, и секторе, противоположном ей (верхняя правая панель). Внизу даны результаты эксперимента. Если животное подвергали стрессу именно за 30 минут до теста, то снижается время, проведенное ею вблизи платформы. Авторы работы трактуют это изменение в поведении как ухудшение памяти. Однако возможно, что возрастает комфортное поведение из-за уменьшения тревоги вследствие увеличения в крови концентрации дезоксикортикостерона, который может легко образовываться из кортикостерона – основного стрессорного гормона у крыс
Один из тестов на память крыс – решение лабиринта Морриса. Он представляет собой бассейн, в котором ниже уровня воды находятся платформы, невидимые для плавающей крысы. Случайно наткнувшись на одну из них, животное должно запомнить ее положение. Если в последующей серии тестов крыса плавает вблизи платформы, значит, она помнит ее положение. Если время, проводимое ею в разных секторах бассейна, примерно одинаково, значит, ее память ослаблена. В указанном эксперименте память ухудшалась, если за 30 минут до тестирования крысу подвергали слабому стрессу или вводили ей кортикостерон (основной глюкокортикоид у мышевидных грызунов). Причем эффект отмечался, если животное подвергали стрессу только за 30 минут до тестирования. Если воздействие производили за две минуты или за четыре часа до теста, то эффект отсутствовал – крыса б?льшую часть времени проводила в секторе около платформы, пытаясь ее нащупать. Авторы исследования делают вывод об ухудшении памяти у животных, у которых за 30 минут до теста повышали содержание экзогенного или эндогенного кортикостерона.
Утверждение об изменении именно памяти спорно. Лабораторная крыса происходит от пасюка, серой крысы Rattus norvegicus, который экологически является околоводным видом. Дикие крысы предпочитают селиться вблизи воды, а их лабораторные потомки часто любят купаться. Поэтому у крысы, помещенной в бассейн, конкурируют две мотивации: избавления от опасности и комфортное поведение. Последнее реализуется при низком уровне тревоги. Сам кортикостерон не обладает противотревожным действием, но только одна химическая реакция отделяет его от дезоксикортикостерона (см. рис. 2.4 и 2.5), противотревожное действие которого хорошо известно. Следовательно, поведенческий эффект и стресса, и введения кортикостерона может иметь механизм, ключевым звеном которого является увеличение концентрации дезоксикортикостерона. Возможно, что именно этот гормон, снижая тревогу, увеличивает возможность реализации комфортного поведения, что проявляется в том, что животные не торопятся выбираться из воды (рис. 4.24).
Интерпретация результатов экспериментов сложна, поскольку разные особи (и люди, и животные) в одинаковой ситуации имеют разные мотивации
Возможность такого объяснения подтверждается тем, что у крыс, которым вживляют медленно растворяющуюся таблетку дезоксикортикостерона, уменьшается количество попыток выбраться из лабиринта Морриса. Таким образом, интерпретация данных экспериментов с введением гормонов всегда затруднительна из-за взаимных влияний эндокринных систем и взаимных превращений стероидных гормонов.
Дополнительную трудность представляет неопределенность мотивации объекта исследования. В рассмотренном примере у животного могут существовать обе мотивации – избавления от опасности и гедонистическая. Определить доминирующую довольно сложно.
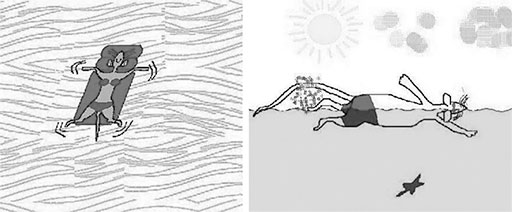
Рис. 4.24. Различные мотивации крыс в бассейне. Они относятся к околоводному виду животных, поэтому многие из лабораторных крыс любят плавать. При этом они могут демонстрировать либо пассивное плавание (панель слева), либо активное (панель справа), что соответствует двум различным поведенческим типам (см. главу 6). Таким образом, погружение в воду крысы могут воспринимать и как стрессорную ситуацию, и как комфортную обстановку. Существование двух стилей поведения – избегания и затаивания – еще больше затрудняет интерпретацию поведения крыс в эксперименте
В исследованиях на человеке, проводимых с помощью опросов и тестирований, опасность ошибки связана с утомлением респондента и потерей интереса к предложенной задаче. Если не учитывать доминирующие мотивации в соответствующих социальных группах, можно прийти к неправильным выводам. Например, простейший тест на способность к экстраполяции: испытуемому предлагают найти предмет, скрытый под одним из бумажных конусов, расположенных в ряд. В каждом из последовательных испытаний предмет перемещается по определенному закону, например на один шаг вправо. Испытуемый должен уловить эту закономерность. Скорость решения растет у детей от 3 до 15 лет, т. е. способность к экстраполяции с возрастом увеличивается. Взрослые же демонстрируют нулевой результат – они опрокидывают все конусы подряд. Эти результаты не означают, что экстраполяционные способности человека достигают пика в 15-летнем возрасте, после чего сходят на нет. Дело в том, что взрослые значительно слабее мотивированы. Они воспринимают подобное занятие как пустую забаву, не заинтересованы заслужить похвалу экспериментатора, их самооценка не увеличится от успешного решения подобной задачи, к тому же конфета как чаще всего используемый «предмет» не интересует взрослого человека. Если же повысить его мотивацию, объявив, например, что результаты тестирования будут влиять на карьерный рост, то результаты взрослых значительно превышают результаты 15-летних подростков.
Еще одним примером неопределенной мотивации испытуемого служит ухудшение результатов при неоднократном предъявлении одной и той же задачи. Если с самого начала человек или животное быстро с ней справляется, то при последующих предъявлениях такого же теста процент успешных решений значительно снижается. Если же задача сложна для испытуемого, то процент правильных решений постепенно, но монотонно нарастает. Л. В. Крушинский предположил, что снижение процента правильных решений связано с неврозоподобным состоянием, развивающимся у тех, кто успешно решает задачу, вследствие сильного напряжения нервно-психических функций. Но, возможно, что у испытуемого меняется мотивация. Решение конкретной задачи не представляет для него интереса, и он начинает экспериментировать со средой.
Для выявления мотивации объекта, поведение которого изучается, очень полезным бывает определение гормональных показателей. Рассказ о роли гормонов как биологических маркеров психических состояний продолжим в следующих главах.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
III. ОБРАЗОВАНИЕ ЗЕМНОЙ КОРЫ
III. ОБРАЗОВАНИЕ ЗЕМНОЙ КОРЫ Огненно-жидкое состояние земли закончилось, как мы видели, образованием сплавленной коры, которую всю покрыла вода. Как же образовались настоящие, поднимающиеся над водою в виде островов, материки и те разнообразные массы камней и слои земель,
6.4. Аналитико-синтетическая деятельность коры больших полушарий
6.4. Аналитико-синтетическая деятельность коры больших полушарий Множество раздражителей внешнего мира и внутренней среды организма воспринимаются рецепторами и становятся источниками импульсов, которые поступают в кору больших полушарий. Здесь они анализируются,
ГЛАВА 3 Эволюция земной коры. Дрейф континентов и спрединг океанического дна. Мантийная конвекция
ГЛАВА 3 Эволюция земной коры. Дрейф континентов и спрединг океанического дна. Мантийная конвекция Горные породы, формирующие кору Земли, как мы помним, бывают изверженные — первичные, образовавшиеся при охлаждении и затвердевании магмы, и осадочные — вторичные,
Как научились читать историю земной коры
Как научились читать историю земной коры В предыдущей главе мы пытались окинуть взглядом великую книгу природы. Но и это самое общее представление могло сложиться только в результате упорного труда выдающихся умов как основателей геологической науки, так и оставшихся
УЧАСТИЕ КОРЫ В ОРГАНИЗАЦИИ НАГЛЯДНЫХ ПРОСТРАНСТВЕННЫХ СИНТЕЗОВ
УЧАСТИЕ КОРЫ В ОРГАНИЗАЦИИ НАГЛЯДНЫХ ПРОСТРАНСТВЕННЫХ СИНТЕЗОВ Зоны, участвующие в организации наглядных пространственных синтезов, находятся на стыке затылочных, височных и постцентральных областей коры больших полушарий (нижнетеменная область – поля Бродмана 21, 37,
Гормоны надпочечников
Гормоны надпочечников Гормоны мозгового вещества надпочечников В мозговом веществе надпочечников в хромаффинных клетках синтезируются катехоламины – дофамин, адреналин и норадреналин. Непосредственным предшественником катехоламинов является тирозин. Норадреналин
Гормоны мозгового вещества надпочечников
Гормоны мозгового вещества надпочечников В мозговом веществе надпочечников в хромаффинных клетках синтезируются катехоламины – дофамин, адреналин и норадреналин. Непосредственным предшественником катехоламинов является тирозин. Норадреналин образуется также в
Гормоны коры надпочечников (кортикостероиды)
Гормоны коры надпочечников (кортикостероиды) В коре надпочечников синтезируются более 40 различных стероидов, различающихся по структуре и биологической активности. Биологически активные кортикостероиды объединяются в 3 основные класса:1. глюкокортикоиды, оказывающие
Периферические гормоны
Периферические гормоны В этом разделе будут рассмотрены локализация, функции, регуляция синтеза и секреции гормонов периферических желез. Расположение основных желез в теле человека показано на рис. 2.3. Рис. 2.3. Схема расположения основных желез в теле человека Масса
Стероидные гормоны
Стероидные гормоны Все вышеперечисленные гормоны относятся к пептидам. Периферические гормоны, продуцируемые корой надпочечников и половыми железами, относятся к химическому классу стероидов.Стероиды отличаются от пептидов не только химически, но и физиологически.
Гормоны при стрессе
Гормоны при стрессе Любое воздействие на организм приводит прежде всего к активации симпатической нервной системы, в результате чего усиливается выделение адреналина из мозгового слоя надпочечников и норадреналина из нервных окончаний в стенках сосудов. Следует
Гормоны и депрессия
Гормоны и депрессия Если гуморальные факторы, модулирующие тревожные состояния, достаточно хорошо известны (табл. 5.1), то «гормон депрессии» остается загадкой. Не вызывает сомнения, что гормоны участвуют в патогенезе депрессивных состояний.Таблица 5.1. Гормоны при
Гормоны и мозг
Гормоны и мозг В определенном смысле, причина межполовых различий — не в том, что у женщин и мужчин гены поведения сами по себе разные. Допустим, у плейстоценового мужчины возникает ген, улучшающий чувство направления, но при этом ухудшающий социальную интуицию. Ему он
5. Анализаторная и синтетическая деятельность коры головного мозга
5. Анализаторная и синтетическая деятельность коры головного мозга Высшая нервная деятельность животного дает возможность выделять ему из сложной массы всевозможных раздражений, идущих из внешнего мира, те, которые имеют для животного то или иное значение. Собака при
Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва)
Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва) Успехи, современной нейрофизиологии, связанные с использованием новых тонких методов электрофизиологического исследования мозга, позволили подойти к изучению центральных механизмов
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва)
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва) Изучению реакции организма на длительное голодание посвящено много работ. Большинство из них касается обменных процессов, биохимии синтеза и распада
Жуков. Дмитрий Анатольевич
Просмотр ограничен
Смотрите доступные для ознакомления главы 👉