Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва)
Особенности возбуждения коры головного мозга у животных при длительном голодании
Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва)
Изучению реакции организма на длительное голодание посвящено много работ. Большинство из них касается обменных процессов, биохимии синтеза и распада пластических структур живой клетки, влиянию голода на систему кроветворения и т. д. (5, 7, 11).
Рядом исследователей, изучавших поведенческие реакции у длительно голодавших животных, было отмечено нарастание агрессивности пищедобывательных реакций, свидетельствующие о вовлечении в процесс симпатоадреналовой системы (7).
Вместе с тем в последние годы появился ряд работ, рассматривающих проблему голода с нейрофизиологической точки зрения.
В лаборатории П. К. Анохина (2) работами К. В. Судакова (8, 9) было показано, что состояние физиологического голода у животных определяется избирательной активацией передних отделов коры мозга, которая носит строго специфический характер и обусловливается восходящими активирующими влияниями гипоталалических отделов пищевого центра. Эта избирательная активация коры головного мозга у голодных животных как выражение определенного доминантного состояния, отчетливо регистрируется даже у животных, находящихся под уретановым наркозом. Таким образом оказалось, что уретан, как наркотическое вещество с избирательным действием на механизмы корково-подкорковых взаимоотношений, не блокирует систему пищевого возбуждения у голодных животных.
В электроэнцефалограмме животных, находящихся под уретановым наркозом, состояние физиологического голода выражается в форме низкоамплитудной высокочастотной электрической активности в передних отделах коры, тогда как в теменно-затылочных отделах наблюдается высокоамплитудная медленная электрическая активность. В работах ряда исследователей (3, 6) также показано, что электрическая, активность коры мозга и ретикулярной формации различная в состоянии голода и насыщения.
Однако специальных работ, посвященных изучению электрической активности коры мозга при длительном голодании животных, нам обнаружить не удалось.
В связи с этим мы поставили перед собой задачу исследовать, как окажется в ЭЭГ животного состояния длительного голодания? Имеются ли при этом какие-либо отличия от ЭЭГ животных при кратковременном голодании?
МЕТОДИКА
Эксперименты были проведены на 20 кошках, которые в течение определенного времени (от 2 до 6 суток) полностью лишались пищи. Вода предоставлялась животным в неограниченном количестве.
В 15 опытах использовались животные, голодавшие более 4-х суток, в 5 опытах от 1 до 3 суток. Опыты проводились под уретановым наркозом. Уретан вводился внутрибрюшинно в 7,5% растворе из расчета 1,5 г на 1 кг веса животного.
Под наркозом у животных обнажалась поверхность черепа, в кость вкалывались стальные игольчатые электроды. Отведение биопотенциалов от лобных, сензомоторных, теменных и затылочных областей коры мозга осуществлялось униполярным методом. Индиферентный электрод располагался по средней линии, над лобной пазухой. ЭЭГ регистрировалась на 10-канальном электроэнцефалографе фирмы «Альвар-Электроник».
РЕЗУЛЬТАТЫ ОПЫТОВ
В первой серии экспериментов изучались изменения ЭЭГ при длительном лишении животных пищи (свыше 3 суток). ЭЭГ этих животных в ряде опытов сравнивалась с ЭЭГ животных после 1—2-суточного голодания. Как уже указывалось выше, у животных после 1—2-суточного голодания наблюдается избирательная активация ЭЭГ передних отделов коры мозга, тогда как в теменно-затылочных отведениях регистрируется высокоамллитудная медленная активность (8). Этот факт был подтвержден вашими исследованиями.
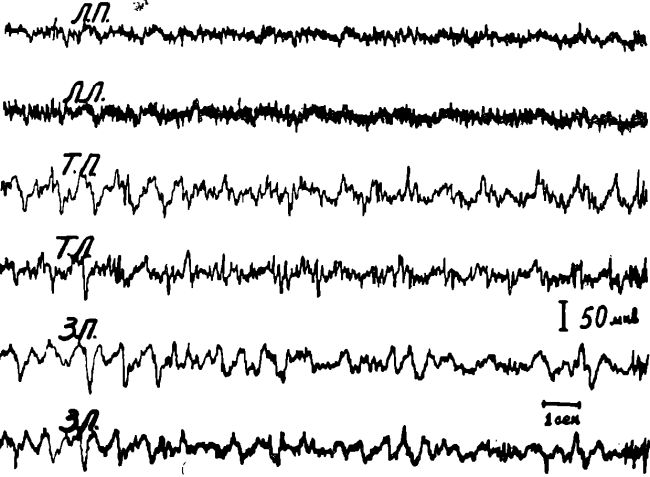
Рис. 1. Электроэнцефалограмма у животного (кошки) в состоянии физиологического голода (1—2 суток).
Отведение от областей (здесь и на последующих рисунках):
ЛП и ЛЛ — лобная правая и левая; СМП и СМЛ — сензомоторная правая и левая; ТП и ТЛ — теменная правая и левая; ЗП и ЗЛ — затылочная правая и левая.
Однако наши опыты показали, что по мере удлинения сроков голодания, начиная с 3-го дня, высокочастотная низкоамплитудная активность (реакция десинхронизации) начинает регистрироваться не только в передних отделах коры, но и все в большей степени в теменно-затылочных отделах.
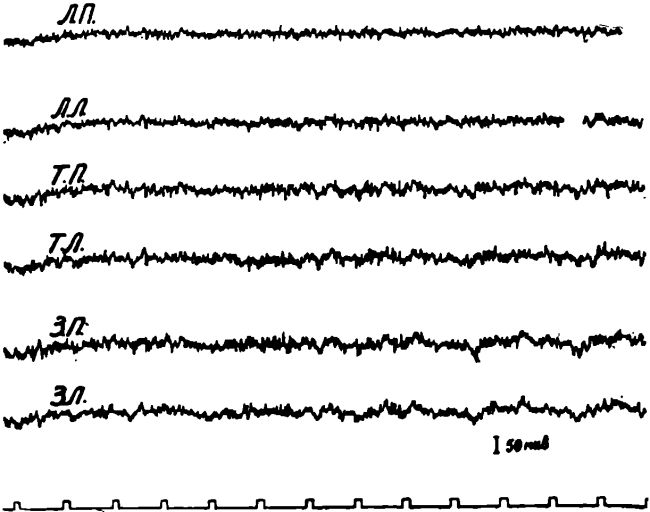
Рис. 2. ЭЭГ у животного в состоянии длительного голода (свыше 4 суток).
а — исходная ЭЭГ животного после 4-суточного голодания; б — после введения аминазина 5 мг/кг.
В ряде опытов у длительно голодавших животных, находящихся под уретановым наркозом, мы наблюдали периодические вспышки высокоамплитудного упорядоченного ритма (8—10 к/сек), временами группирующегося в веретена. Эта активность несколько напоминает тот высокоамплитудный упорядоченный ритм, который ряд авторов рассматривает, как «ритм напряжения» (4, 12, 13). Все эти опыты указывали на то, что при длительном голодании животных наблюдается резкое повышение восходящих активирующих влияний со стороны подкорковых образований на кору мозга.
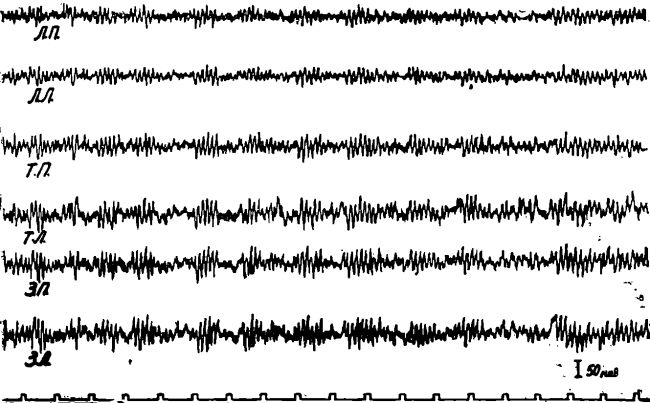
Рис. 3. Высокоамплитудный упорядоченный ритм в электроэнцефалограмме кошки при длительном голодании.
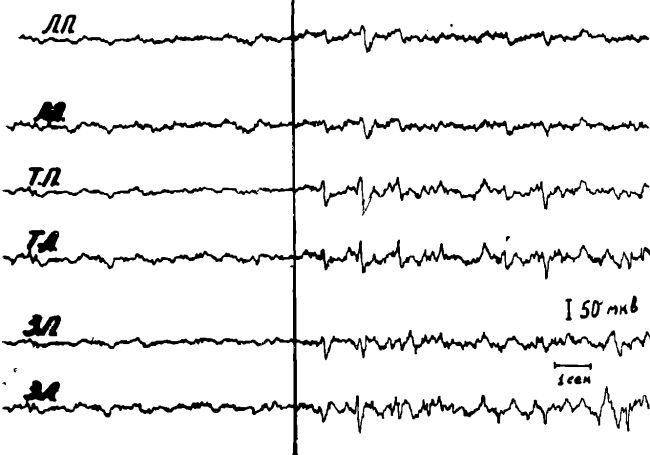
Рис. 4. Изменение ЭЭГ у длительно голодавших кошек после введения в сонную артерию 5% глюкозы.
а — исходная ЭЭГ животного после 4-суточного голодания; б — через 2 мин. после введения 10,0 мл 5% глюкозы.
Известно, что состояние голода первично складывается в гипоталамических отделах пищевого центра под влиянием раздражения «голодной» кровью, и только после этого пищевое возбуждение избирательно вовлекает в свою деятельность синаптические и клеточные образования коры мозга (14, 15).
Поэтому для того, чтобы установить в какой степени генерализованная активация коры мозга у животных при длительном голодании связана с деятельностью именно «пищевого центра», мы решили исследовать, как скажется на ЭЭГ этих животных введение раствора глюкозы, как фактора насыщения. Для этой цели в кровяное русло животных вводился 5% раствор глюкозы. Опыты показали, что введение глюкозы животным после длительного голодания приводило только к кратковременному 2—3 мин. устранению ЭЭГ активации преимущественно в теменно-затылочных отделах коры, в то время как в передних ее отделах сохранялась стойкая реакция десинхрнизации.
Таким образом, введение глюкозы животным после длительного голодания не могло заблокировать у них восходящие активирующие влияния гипоталамических отделов пищевого центра на передние отделы коры мозга. Все это указывало на то, что функциональная активность этих центров гипоталамуса у животных при длительном голодании резка возрастала и малые дозы глюкозы уже не были способны ее подавить. Вместе с тем введение глюкозы приводило к устранению генерализованной активности коры у этих животных и превращало ее в локальную ЭЭГ активацию, подобную той, какая наблюдалась у животных после кратковременного голодания. Все это позволяло думать, что генерализованная активация, наблюдаемая у животных после длительного голодания, определялась распространением влияний сильна возбужденных гипоталалических отделов пищевого центра-на другие активирующие аппараты подкорковой области.
Рядом авторов было установлено, что генерализованная активация коры мозга может быть обусловлена адренэргическим субстратом ретикулярной формации ствола мозга (18).
В связи с этим для анализа участия активирующих восходящих аппаратов ретикулярной формации в формировании генерализованной реакции десинхронизации ЭЭГ у животных при длительном голодании в наших опытах был применен аминазин.
Все это позволяло думать, если генерализованная активация ЭЭГ, которая наблюдалась у животных после длительного голодания, обусловливается восходящими активирующими влияниями андренэргичеаких элементов ретикулярной формации, то введение аминазина должно избирательно устранить эту реакцию, не повлияв заметным образом на регионарную активацию ЭЭГ в передних отделах коры мозга. Наши опыты подтвердили такое предположение.
Известно, что аминазин как адренолитическое средство, избирательно блокирует механизмы восходящих активирующих влияний ретикулярной формации ствола мозга, которые осуществляются андренэргическим субстратом (1, 2, 16).
Экспериментальные факты (10) показали, что внутримышечное введение аминазина в дозе, не превышающей 5 мг на 1 кг веса животного, как правило, не устраняет избирательной активации передних отделов коры у животных после 1—2-суточного голодания, оказывая в то же время блокирующее действие на механизмы генерализованной по коре болевой активации.
В подавляющем большинстве наших опытов, введение животным после длительного голодания аминазина в указанной выше дозе устраняло активацию коры только в теменно-затылочных областях. При этом в передних ее отделах по-прежнему сохранялась реакция десинхронизации. Последняя устранялась только после введения больших доз аминазина (свыше 5 мг на 1 кг веса животного).
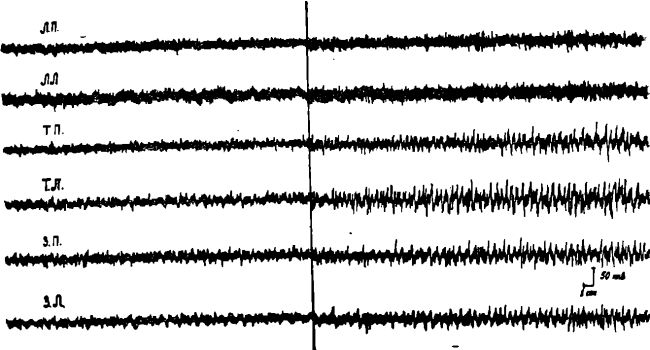
Рис. 5. Изменение ЭЭГ у длительно голодающих кошек после внутримышечного введения аминазина.
Таким образом, проведенные нами эксперименты дают основание считать, что по мере увеличения сроков голодания, в механизм восходящей активации коры мозга, кроме специфических гипоталамических структур пищевого центра, все в большей степени вовлекаются активирующие аппараты ретикулярной формации среднего мозга. Это отличает состояние длительного голода от физиологического, при котором активация коры головного мозга осуществляется преимущественно за счет восходящих активирующих влияний пищевых центров гипоталамуса.
На основании вышеизложенного, механизмы генерализованной восходящей активации коры мозга при длительном голодании можно представить следующим образом.
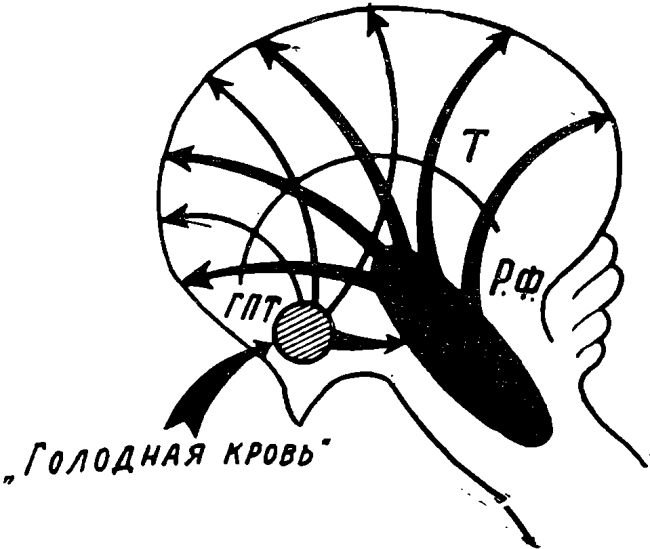
Рис. 6. Схематическое изображение восходящих активирующих влиянии гипоталамуса на кору головного мозга при длительном голодании.
Обозначения: T — таламус, РФ — ретикулярная формация, ГПТ — гипоталамус.
Схема показывает, что при длительном голодании генерализованная активация коры головного мозга осуществляется за счет влияния со стороны гипоталамуса и адренэргических элементов ретикулярной формации.
По мере увеличения сроков голодания под влиянием действия «голодной» крови и нервной сигнализации от пищеварительных органов происходит значительное повышение активности пищевых центров гипоталамуса. Это, в свою очередь, ведет к иррадиации возбуждения в сторону андренэргического субстрата ретикулярной формации ствола мозга, возбуждение которой приводит к расширению потока восходящих активирующих влияний и генерализованной активации всей торы мозга. Такая реакция, по нашему мнению, в отличие от чисто пищевого возбуждения носит уже характер реакции общего напряжения организма (Г. Селье) (19). Она развивается в связи с угрозой для его жизни.
Именно на основе таких генерализованных влияний на все элементы коры мозга организм приобретает способность направить все свои усилия на поиск и потребление пищи. Это влияние, формируясь на основе адренергических субстратов ретикулярной формации ствола мозга, обладает высокоэнергетической силой воздействия на кору мозга, подчиняя всю ее деятельность единой доминирующей функции — восполнению питательных веществ в организме.
ЛИТЕРАТУРА
1. Агафонов В. Н. Журн. невропатол. и психиат., 1964, 56 (2), 94.
2. Анохин П. К. Физиол. журнал СССР, 1957, 43, № 11, 1072.
3. Бакурадзе А. Н. Матер. I конф. по ретикулярной формации ствола мозга. 1961.
4. Банцекина М. М. Канд. дисс., М., 1963.
5. Ильин-Куку ев Б. И. Сборник работ по физиологии, I МОЛМИ, М., 1939, стр. 114.
6. Мариц А. М. Физиол. журнал СССР, 1962, т. 48, № -8, с. 889.
7. Молчанова О. П., Ежова Е. Н. Сборник работ по физиологии, I МОЛМИ, М„ 1939, стр. 85.
8. Судаков К- В. Физиол. журн. СССР, 1962, 48, № 2, 165.
9. Судаков К. В. Физиол. журн. СССР, 1963, 49, № 9, 901.
10. Фадеев Ю. А., Судаков К- В. Физиол. журн. СССР, 1963, 49, № 11, 1310.
11. Шапиро Ю. Л. Канд. дисс. Москва, 1965.
12. Шумилина А. И. >Kvp<h. невропатол. психиатр., 1956, 56, № 2.
13. Шумилина А. И. Физиол. журн. СССР, 1956, 45, № 10, 1176.
14. An and В. К-, Brobeck J. R. Vale journ. Biol. Med., 1951, 24 123.
15. Brobeck J. R. Vale journ. Biol. a. Med., 1957, 29, 565.
16. Dell P., Bonvallet M., H u g i 1 i n A., EEG a. clin. Neu-rophyciol., 1954, 6, 599.
17. Кен нон В. Физиология эмоций. Л., 1927.
18. М о г u z z i G., Magoun Н. W. EEG clin. Neurophysiol., 1949, 3, 455.
19. Селье Г. «Очерки об адаптационном синдроме». И. Л., М., 1960.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Травмы головного мозга
Травмы головного мозга Встречаются относительно редко.¦ ЭТИОЛОГИЯ И ПАТОГЕНЕЗТравмы головного мозга регистрируются после ударов или падения с высоты. Сопровождаются сотрясением мозга и кровоизлияниями различной степени.¦ СИМПТОМЫПосле удара или падения собака
5. Анализаторная и синтетическая деятельность коры головного мозга
5. Анализаторная и синтетическая деятельность коры головного мозга Высшая нервная деятельность животного дает возможность выделять ему из сложной массы всевозможных раздражений, идущих из внешнего мира, те, которые имеют для животного то или иное значение. Собака при
Восходящие активирующие влияния на кору головного мозга у голодных животных К. В. СУДАКОВ (Москва)
Восходящие активирующие влияния на кору головного мозга у голодных животных К. В. СУДАКОВ (Москва) В последние годы благодаря применению новых методов физиологического эксперимента (электронные усилители, стереотаксическая техника) проблема голода, пищевого поведения
К характеристике изменения ЭЭГ у больных, находящихся на длительном лечебном голодании В. В. АРШАВСКИЙ, Б. В. КРАЙЦЕРОВ (Москва)
К характеристике изменения ЭЭГ у больных, находящихся на длительном лечебном голодании В. В. АРШАВСКИЙ, Б. В. КРАЙЦЕРОВ (Москва) Из многочисленных данных литературы известно, что центральный отдел висцерального анализатора как у животных, так и у человека проецируется в
Анализ физиологических механизмов восходящих активирующих влияний на кору мозга у голодных животных методом аппликации различных фармакологических веществ А. И. ТУРЕНКО (Москва)
Анализ физиологических механизмов восходящих активирующих влияний на кору мозга у голодных животных методом аппликации различных фармакологических веществ А. И. ТУРЕНКО (Москва) Многочисленные исследования показали, что основным регулятором потребления пищи является
Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва)
Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва) Успехи, современной нейрофизиологии, связанные с использованием новых тонких методов электрофизиологического исследования мозга, позволили подойти к изучению центральных механизмов
Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва)
Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва) Голодание, как частный раздел науки о питании, за последние годы вновь стало привлекать к себе пристальное внимание в связи со все более широким его
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва)
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва) Как известно, при полном алиментарном голодании в течение довольно длительного времени (по крайней мере до сроков, характеризующихся потерями
Сывороточное и эритроцитарное железо при длительном голодании людей Ю. Л. ШАПИРО, Л. М. ДОНДЫШ, Л. М. ЛЕЙБИН, Э. А. ЛЕЙЗЕРОВИЧ, Л. Ф. ЛЕВИНА (Москва)
Сывороточное и эритроцитарное железо при длительном голодании людей Ю. Л. ШАПИРО, Л. М. ДОНДЫШ, Л. М. ЛЕЙБИН, Э. А. ЛЕЙЗЕРОВИЧ, Л. Ф. ЛЕВИНА (Москва) Данные о содержании железа в жидкой фракции крови при полном алиментарном голодании и последующем питании немногочисленны и
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва)
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва) В литературе имеется значительное число работ, посвященных изучению изменения степени насыщения крови кислородом при разнообразных условиях и
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва)
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва) Известно, что так называемые «жесткие константы» сохраняются неизменными в организме В течение длительного срока, несмотря на воздействие каких-либо «отклоняющих» факторов.
О динамике основного обмена при лечебном голодании больных с ипохондрическим синдромом В. Б. ГУРВИЧ (Москва)
О динамике основного обмена при лечебном голодании больных с ипохондрическим синдромом В. Б. ГУРВИЧ (Москва) Вопрос об обмене веществ, в частности, основном обмене, является одним из ведущих при объяснении патофизиологических механизмов голодания (1, 5, 9, 11, 12, 14).В. В.
СОСТОЯНИЕ ИММУНОБИОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ ОРГАНИЗМА ЧЕЛОВЕКА ПРИ ПОЛНОМ ДЛИТЕЛЬНОМ ГОЛОДАНИИ Ю. С. НИКОЛАЕВ, В. А. СКОРИК-СКВОРЦОВА (Москва)
СОСТОЯНИЕ ИММУНОБИОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ ОРГАНИЗМА ЧЕЛОВЕКА ПРИ ПОЛНОМ ДЛИТЕЛЬНОМ ГОЛОДАНИИ Ю. С. НИКОЛАЕВ, В. А. СКОРИК-СКВОРЦОВА (Москва) Изучение состояния иммунобиологической реактивности организма в период длительного полного голодания и последующего
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва)
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва) В течение последних лет в нашей стране и за рубежом появилось большое число сторонников применения голодания с лечебной целью при
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва)
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва) Проблема влияния голодания на разные показатели обменных процессов в организме животных и человека продолжает привлекать внимание
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва)
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва) Голодание как метод лечения успешно зарекомендовал себя при некоторых психических и соматических заболеваниях (3, 7, 10—13). Особенно перспективно его применение при обменных, аллергических