Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва)
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей
Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва)
Изучению фагоцитарной активности лейкоцитов при длительном полном алиментарном голодании посвящены единичные работы.
Согласно данным некоторых авторов, фагоцитарная активность при кратковременном голодании (до 36 часов) увеличивалась в 3 раза (3). Фагоцитарная активность нейтрофилов из эксудата брюшины крыс голодавших до периода, сопровождавшегося потерей веса тела на 25—30% К исходному, фактически не изменялась (10).
Нами было проведено исследование фагоцитарной активности лейкоцитов периферической крови у 21 психически больного в процессе лечебного голодания и в период последующего восстановления. Среди обследованных — 19 мужчин и 2 женщины. Возраст колебался от 25 до 40 лет.
По диагнозу больные распределялись следующим образом: шизофрения, простая форма — 4; шизофрения, параноидная форма — 1; ипохондрический синдром — 7; ипохондрическое развитие личности на соматической основе—3, депрессивное состояние — 1; астено-невротический синдром—1; навязчивость—1; диэнцефальный синдром, остаточные явления инфекционного поражения ЦНС — 1; органическое заболевание ЦНС-1.
Сроки лечебного голодания колебались в пределах от 17 до 37 дней. Потеря веса тела не превышала 20% к исходному.
Фагоцитарная активность лейкоцитов определялась по методу Е. А. Кост и М. И. Стенко (6).
Таблица 1
Фагоцитарное число
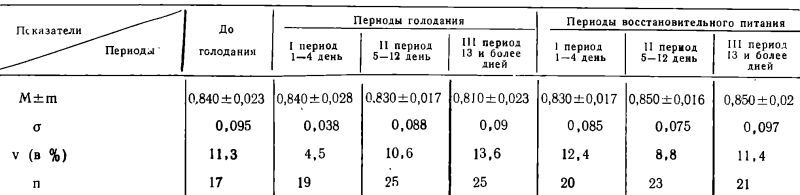
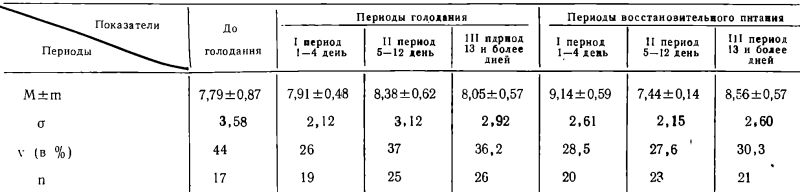
Таблица 2
Фагоцитарный индекс
Методика заключалась в следующем: смесь, состоящая из 1 объема 5% цитрата натрия, 2 объемов крови и I объема однодневной живой культуры золотистого стафилококка в 2 млд разведении помещали на 30 минут в термостат при 37°С. Из смеси приготавливались мазки, которые фиксировались метиловым спиртом. Окраска проводилась по Романовскому — Гимза.
В мазках подсчитывалось число фагоцитировавших нейтрофилов на 100 клеток и количество фагоцитированных микробов в одном нейтрофиле. Количество фагоцитировавших клеток на 100 нейтрофилов обозначалось как «фагоцитарное число» — Ф. Ч. Среднее число фагоцитированных микробов одним нейтрофильным лейкоцитом обозначалось как «фагоцитарный индекс» или «фагоцитарный показатель» — Ф. П.
Наряду с определением фагоцитарного индекса и фагоцитарного. числа изучалась интенсивность перевариваемости микробов. С этой целью последние подразделялись на светлые (подвергшиеся значительному лизису) и темные (лизис был выражен в меньшей степени). Помимо этого учитывалась также степени вариабильности фагоцитоза стафилококков отдельными нейтрофилами (V в %).
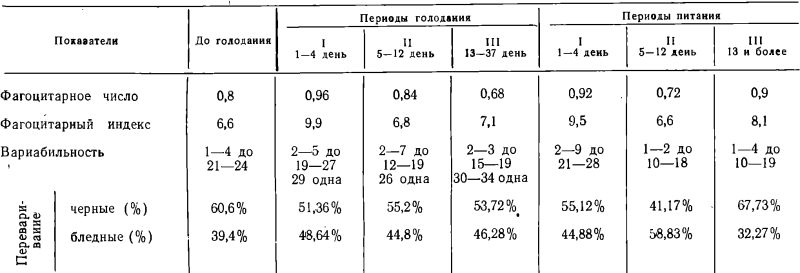
Исследования в большинстве случаев проводились в динамике голодания и последующего питания. Полученные данные группировались но периодам, выделенным при этом состоянии, у людей (Ю. С. Николаев) (2), обрабатывались статистически и были сведены в таблицы 1, 2, 3.
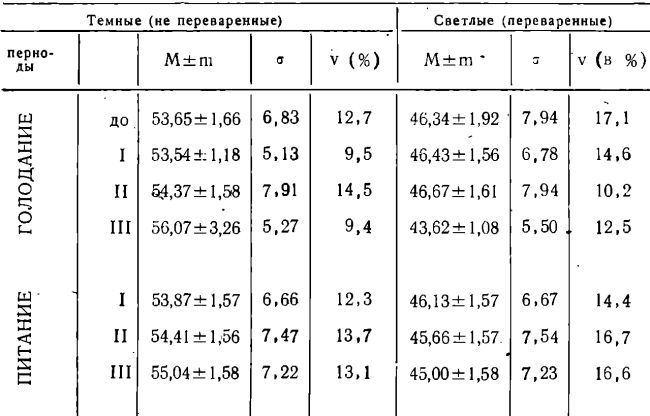
Таблица 3
Внутриклеточное переваривание стафилококков
Как можно видеть из таблицы 1, фагоцитарное число (Ф. Ч.), взятое в среднем во все периоды голодания и последующего питания, практически не отличалось от исходного (до голодания) уровня (Р>0,1).
Фагоцитарный показатель (Ф. П.), отражающий интенсивность фагоцитоза, оказывался столь же стабильным, как и фагоцитарное число, во все сроки голодания и последующего питания (Р>0,05).
Из таблицы 3 можно видеть, что процесс внутриклеточного переваривания стафилококков (в наших исследованиях определяемый по соотношению числа темных-непереваренных к более светлым-лизированным микробам) в течение всех обследованных сроков голодания и питания существенна не изменялся.
Таким образом, средние показатели, отражающие экстенсивность и интенсивность фагоцитарной активности нейтрофилов периферической крови в течение изученных сроков полного алиментарного голодания (с 17 по 37 день) и последующего питания в условиях и в сроки наших наблюдений оставались величинами в достаточной степени стабильными.
Наряду с этим отмечались и разнообразные индивидуальные реакции. Так, в 7 наблюдениях обнаруживалось отчетливое уменьшение фагоцитарного числа и фагоцитарного показателя в первый период голодания (1—4 дни) после чего, как правило, вновь отмечалось их увеличение, и к концу периода голодания (в этих наблюдениях к 17—27 дням) они достигли уровня исходных цифр.
В качестве иллюстрации приводим следующее наблюдение:
Больная А-ва, 1937 года рождения, поступила 27/XI—65 г. диагноз МДП?, депрессивное состояние. Потеря веса к концу голодания 7 кг 500 г (12,8% к исходному) (табл. 4).
В других 6 наблюдениях в первый период голодания обнаруживалось, наоборот, увеличение фагоцитарного числа и фагоцитарного индекса. В дальнейшие сроки голодания эти показатели несколько снижались, но к срокам прекращения голодания (18—22 дни) они вновь достигали исходных цифр.
В качестве иллюстрации приводим следующее наблюдение. Больной А-н, 1928 года рождения, поступил 28/Х—65 года, диагноз — ипохондрическое развитие психопатической личности. Потеря веса к концу голодания 12 кг (16,9% к исходному) (табл. 5).
Интересно отметить, что параллелизм в изменении фагоцитарного числа и фагоцитарного показателя большей частью отмечался лишь в первый период голодания. В отдаленные его сроки можно было видеть, что нередко при снижении фагоцитарного числа фагоцитарный показатель увеличивался, что свидетельствует об интенсификации фагоцитоза. В этих исследованиях обнаруживалась и максимальная вариабильность числа фагоцитируемых микробов (от 2—4 и 23—25 исходных до 2—4 и 32—42 на 7 день голодания).
Таблица 4
Показатели фагоцитарной активности нейтрофилов больной А-ой
Голодала 19 дней
Год рождения— 1937.
Диагноз: МДП? Депрессивное состояние.
Потеря веса — 7 кг 500 г. (12,8% к исходн.)
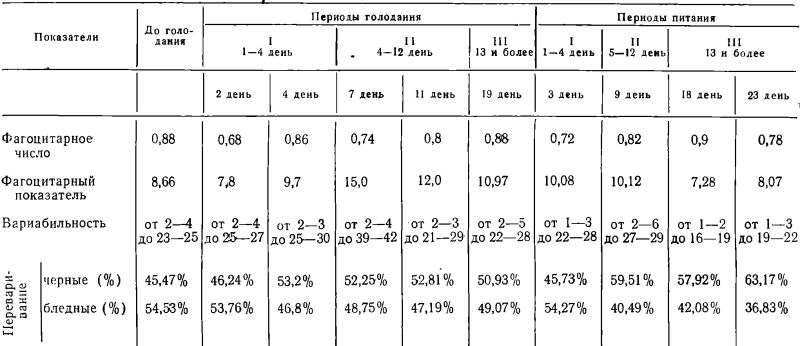
Показатели фагоцитарной активности нейтрофилов больного А-н.
Голодал 22 дня
Год рождения — 1928.
Диагноз: ипохондрическое развитие психопатической личности.
Потеря веса — 12 кг (16,9% к исходн.).
Как уже говорилось выше, фагоцитарная активность лейкоцитов при полном голодании и последующем питании людей практически не изучена. Процесс фагоцитоза, отражающий одно из основных функциональных свойств лейкоцитов, согласно современным представлениям, зависит, по крайней мере, от следующих условий: 1) биологических, физико-химических и прочих свойств фагоцитируемого объекта; 2) воздействий окружающей фагоцит и фагоцитируемый объект среды (белковый, липидный, ионный состав плазмы, содержание опсонинов, концентрация гепарина, стероидные гормоны и прочее); 3) функционального состояния самих фагоцитов.
Со времени работ И. И. Мечникова достаточно твердо установилось мнение, что. особенности среды и фагоцитируемого объекта оказывают лишь стимулирующее или тормозящее влияние на активность фагоцитарного процесса. Основное значение придают функциональной активности самих фагоцитов, в данном случае микрофагов — нейтрофилов (1).
Следует подчеркнуть, что изучению разнообразных характеристик, отражающих функциональные свойства лейкоцитов во время полного голодания и последующего питания посвящены единичные работы. Так, наблюдали (4) уменьшение содержания специфической зернистости в цитоплазме голодавших крыс. Побледнение цитоплазмы (уменьшение в ней базофилии) снижение количества и величины нейтрофильной зернистости отмечал Ю. Л. Шапиро (8), в процессе лечебного голодания психически больных и отдельных «добровольцев-голодалыциков» (сроки голодания от 9 до 40 дней). Эти изменения количественно и качественно увеличивались по мере удлинения сроков голодания. Аналогичные изменения были обнаружены и в костном мозгу в клетках миелоидного ряда. Наиболее отчетливо эти изменения обнаруживались в более зрелых элементах (зрелые миелоциты по Рору, палочкоядерные, сегментоядерные). Одновременно отмечалось и резкое уменьшение митотически делящихся клеток миелоидного ряда. После прекращения голодания (спустя некоторый латентный период) эти изменения претерпевали обратное развитие. Причем, параллельно с усилением базофилии, увеличением числа и укрупнением нейтрофильной зернистости отмечалось нарастание миготической активности клеток миелоидного ряда, а в периферической крови отмечался нейтрофилез с «регенеративным» сдвигом ядра влево.
По некоторым данным при изучении двигательной активности клеток белой крови в течение 28—30 дней лечебного голодания у 7 больных обнаруживалась тенденция к некоторому снижению «скорости» лейкоцитов лишь к концу указанных сроков голодания (с 19,9 µ/мин исходных до 17,7 µ/мин к 28—30 дню голодания) (5). Одновременно было отмечено уменьшение активно двигавшихся нейтрофилов с 51 до 31%, незначительное увеличение числа вялодвижущихся нейтрофилов по первому типу движения (с 32,1% до 44,8%). Интересно отметить, что число неподвижных нейтрофилов, которое и до голода было невелико, в процессе голодания не увеличилось. Было также отмечено нарастание вакуолизации цитоплазмы нейтрофилов, максимально выраженное к 28—30 дню голодания. После прекращения голодания двигательная активность нейтрофилов увеличилась, а количество вакуолей — уменьшилось. Наиболее отчетливо активность нейтрофилов проявлялась с 12 по 16 дни восстановительного периода. Естественно, что указанные данные касались фактически начальных периодов голодания. В терминальные сроки наблюдаются иная картина. Так, согласно некоторым данным, интенсивность амебоидных движений лейкоцитов, полученных от животных, доведенных голоданием до смерти, снижаются быстрее, чем у не голодавших (13). Думм (9), инкубируя суспензию лейкоцитов периферической крови, полученную от здоровых голодавших людей, в плазме той же крови, в которую добавлял глюкозу (2—2,5 части при 37°С), определял убыль глюкозы и прирост молочной кислоты. Согласно полученным им данным, потребление глюкозы лейкоцитами голодавших здоровых людей было несколько ниже, чем у сытых, однако, различие было статистически недостоверно. Образование молочной кислоты лейкоцитами, полученными от голодавших людей, не отличалось от количества, вырабатываемого сытыми. Интересно, что добавление к среде инсулина не всегда повышало потребление глюкозы лейкоцитами голодавших и повышало потребление глюкозы в суспензии лейкоцитов, полученных от сытых людей.
Можно видеть, что данные относительно изучения некоторых показателей, отражающих функциональные свойства лейкоцитов, весьма немногочисленны, и ими трудно объяснить стабильность фагоцитарной активности, отмеченную в наших наблюдениях.
Несомненно, что этот вопрос нуждается в комплексном изучении многих параметров, отражающих как функциональное состояние самих лейкоцитов (содержание в них энергетических веществ, ферментов и т. д.), так и средовых факторов.
В отношении особенностей фагоцитарной активности нейтрофилов, отмеченных в первый период голодания (можно полагать), что они зависят от появления в периферической крови двух различных популяций лейкоцитов. Так, Ю. Л. Шапиро (7) наблюдал в периферической крови при голодании как нейтрофилы «с молодым» рыхлым, двухсегментным ядром, большие по размеру, так и одновременно нейтрофилы, содержащие 4—5 гиперхроматичных сегментов, малых по размеру. Эти данные несколько позже были подтверждены экспериментально. Ряд авторов отмечал, что у голодавших собак с выведенной наружу селезенкой, количество лейкоцитов в последней было на 50% ниже, чем в крови из бедренной артерии (12). При этом ядра гранулоцитов периферической, крови в большинстве клеток содержали 2—3, а в селезенке 4—5 и более сегментов. После раздражения селезенки в периферической крови значительно увеличилось количество многосегментных гранулоцитов. Авторы заключают, что при образовании депо в селезенке в ней избирательно задерживаются более зрелые гранулоциты.
Можно полагать, что в первый период голодания (рассматриваемый как стадия тревоги адаптационного синдрома по Селье), из селезенки эксдепонируется часть многосегментных нейтрофилов. Первая группа (малосегментные нейтрофилы) надо полагать, поступают в периферическую кровь из костного мозга. Можно думать, что от соотношения этих популяций нейтрофилов (отличающихся в «возрастном» и, следовательно, в функциональном отношении) во многом зависят особенности фагоцитарной реакции во время полного голодания, особенно в начальные его сроки.
Открытым остается вопрос о значении миграции во время голодания нейтрофилов вместе с хиломикронами в капиллярную сеть легких, откуда они, как известно, вновь могут возвращаться в периферическую кровь (11).
В целом, как можно было видеть из приведенных данных, фагоцитарная активность нейтрофилов при полном голодании людей в условиях и сроки наших наблюдений остается вполне сохранной. Конкретные механизмы, лежащие в основе стабильности защитных свойств лейкоцитов при голодании, остаются малоизученными, что, естественно, должно явиться стимулом к дальнейшим исследованиям.
ЛИТЕРАТУРА
1. Адо А. А. Патофизиология фагоцитов. М., 1961.
2. Николаев Ю. С. Разгрузочно-диетическая терапия шизофрении и ее физиологическое обоснование. Дисс. докт., М., 1959.
3. Планельес X. Фагоцитоз Б.М.Э., М., 1963, изд. 2, т. 33, с. 428.
4. Рыжих Р. Н. Д.АН СССР, 1952, т. 37, № 6, с. 1051.
5. С р а б и о п о в а В. X., Хотеев а Г. И. Вопр. теоретич, и практич. мед. Ростов-на-Дону, 1965, с. 68.
6. Тодоров И. Клинические лабораторные исследования в педиатрии. София, 1963, 4 изд., 382.
7. Шапиро Ю. Л. Патологическая физиология и экспер. терапия им. В. В. Пашутина, 1963, 1, с. 39.
8. Шапиро Ю. Л. Состояние системы крови при полном длительном алиментарном голодании и последующем питании людей. Дисс. канд., М., 1964.
9. Duram М. Е. Proc. Soc. Exptl. Biol, and Med., 1957, 95, No. 3, p. 571.
10. С u с t a n о L., F e г г i с i i о S. Т. Rend., CI. sci. fis., mat., e natur., 1960 (1961), 29, No. 5, p. 424.
11. Cnderblitzen Th. Schweiz. ned Wochenschz, 1954, 84, No. 40, P- 1150.
12. L u d a n у G., R i g о С., Budavari G., Han To Wu (1964) Med. exptl., 1964, 11, No. 2, 105—109.
13. Nagac T. Nagasaki igakkai Zassci, Nagasaki Med. G., 1958, 33, No. 5, p. 570,
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва)
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва) Лечение ожирения остается одной из наиболее актуальных проблем современной медицины. Число больных с избыточным весом, согласно данным многих
Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва)
Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва) Голодание, как частный раздел науки о питании, за последние годы вновь стало привлекать к себе пристальное внимание в связи со все более широким его
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва)
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва) Как известно, при полном алиментарном голодании в течение довольно длительного времени (по крайней мере до сроков, характеризующихся потерями
Сывороточное и эритроцитарное железо при длительном голодании людей Ю. Л. ШАПИРО, Л. М. ДОНДЫШ, Л. М. ЛЕЙБИН, Э. А. ЛЕЙЗЕРОВИЧ, Л. Ф. ЛЕВИНА (Москва)
Сывороточное и эритроцитарное железо при длительном голодании людей Ю. Л. ШАПИРО, Л. М. ДОНДЫШ, Л. М. ЛЕЙБИН, Э. А. ЛЕЙЗЕРОВИЧ, Л. Ф. ЛЕВИНА (Москва) Данные о содержании железа в жидкой фракции крови при полном алиментарном голодании и последующем питании немногочисленны и
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва)
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва) В литературе имеется значительное число работ, посвященных изучению изменения степени насыщения крови кислородом при разнообразных условиях и
Динамика показателей периферической крови при лечебном голодании у больных гипертонической болезнью и ожирением Г. Н. БЖИШКЯН-БОРОДИНА (Москва)
Динамика показателей периферической крови при лечебном голодании у больных гипертонической болезнью и ожирением Г. Н. БЖИШКЯН-БОРОДИНА (Москва) Литературные данные относительно исследования состава периферической крови при лечебном голодании представляют большую
Сравнительное изучение действия полного длительного голодания И белковой недостаточности на состав периферической крови мышей СС57 Вr И. Л. ПОВЕРИЙ и В. И. ПРИЛЯЦКИЙ (Москва)
Сравнительное изучение действия полного длительного голодания И белковой недостаточности на состав периферической крови мышей СС57Вr И. Л. ПОВЕРИЙ и В. И. ПРИЛЯЦКИЙ (Москва) Полное или частичное голодание является удобной экспериментальной моделью для изучения влияния
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва)
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва) Известно, что так называемые «жесткие константы» сохраняются неизменными в организме В течение длительного срока, несмотря на воздействие каких-либо «отклоняющих» факторов.
О действии полного длительного алиментарного голодания на хромосомный аппарат лимфоцитов периферической крови К. Н. ГРИНБЕРГ, Ю, Л. ШАПИРО, Е. А. КИРИЛОВА, Р. С. КУШНИР (Москва)
О действии полного длительного алиментарного голодания на хромосомный аппарат лимфоцитов периферической крови К. Н. ГРИНБЕРГ, Ю, Л. ШАПИРО, Е. А. КИРИЛОВА, Р. С. КУШНИР (Москва) Полное алиментарное голодание успешно применяется при лечении некоторых психических и
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва)
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва) Половой хроматин — внутриядерное тельце, дающее Фельген-положительную реакцию и
СОСТОЯНИЕ ИММУНОБИОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ ОРГАНИЗМА ЧЕЛОВЕКА ПРИ ПОЛНОМ ДЛИТЕЛЬНОМ ГОЛОДАНИИ Ю. С. НИКОЛАЕВ, В. А. СКОРИК-СКВОРЦОВА (Москва)
СОСТОЯНИЕ ИММУНОБИОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ ОРГАНИЗМА ЧЕЛОВЕКА ПРИ ПОЛНОМ ДЛИТЕЛЬНОМ ГОЛОДАНИИ Ю. С. НИКОЛАЕВ, В. А. СКОРИК-СКВОРЦОВА (Москва) Изучение состояния иммунобиологической реактивности организма в период длительного полного голодания и последующего
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва)
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва) В течение последних лет в нашей стране и за рубежом появилось большое число сторонников применения голодания с лечебной целью при
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва)
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва) Проблема влияния голодания на разные показатели обменных процессов в организме животных и человека продолжает привлекать внимание
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва)
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва) Голодание как метод лечения успешно зарекомендовал себя при некоторых психических и соматических заболеваниях (3, 7, 10—13). Особенно перспективно его применение при обменных, аллергических
Некоторые данные относительно белково-азотистого обмена в процессе лечебного голодания психически больных Л. И. ЛАНДО, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. Я. БАБЕНКОВ (Москва)
Некоторые данные относительно белково-азотистого обмена в процессе лечебного голодания психически больных Л. И. ЛАНДО, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. Я. БАБЕНКОВ (Москва) Белково-азотистый обмен при полном алиментарном голодании как в эксперименте на животных, так и у людей