Некоторые данные относительно белково-азотистого обмена в процессе лечебного голодания психически больных Л. И. ЛАНДО, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. Я. БАБЕНКОВ (Москва)
Некоторые данные относительно белково-азотистого обмена в процессе лечебного голодания психически больных
Л. И. ЛАНДО, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. Я. БАБЕНКОВ (Москва)
Белково-азотистый обмен при полном алиментарном голодании как в эксперименте на животных, так и у людей исследовался неоднократно (5, 9, 14, 16-18, 21, 29, 31, 34, 35, 39, 41-43, 47, 48, и др.)
Интерес к этой проблеме, несомненно, связан, прежде всего, с той особой ролью в жизнедеятельности организма, которую играет белок как источник специфического пластического материала и энергии, а также в силу его транспортных и других свойств.
В задачу наших исследований входило динамическое изучение ряда компонентов белково-азотистого обмена в крови и в моче при лечении психически больных методом полного дозированного голодания (7).
У 29 больных были проведены исследования в сыворотке крови: общего белка (рефрактометрически), белковых фракций (электрофорезом на бумаге), у 19 из них — транаминазы (по Умбрайту в модификации Т. С. Пасхиной); в моче определялись: общий азот (сжиганием и отгонкой в чашках Конвея), мочевина (уреазным методом), аминоазот (по Поп-Стивенсону), аммиак (в чашках Конвея).
Забор материала производился: троекратно до начала голодания (исходный фон), во время голодания и питания также 3 раза в сроки, соответствующие периодам наиболее выраженных клинических изменений в состоянии больных (7).
Полученные результаты обработаны методом вариационной статистики.
По клинической характеристике больные распределялись следующим образом: по нозологическому диагнозу—шизофрения—18 чел. (простая форма — 9, ипохондрическая — б, параноидная — 3), затянувшееся реактивное состояние — 3 чел., патологическое развитие — 8 человек. Всего было 25 мужчин и 4 женщины. В период обследования 3 больных были в возрасте от до 20 лет, 16 — от 21 года до 30 лет, 6 — от 31 года до 40 лет, 4 — старше 40 лет. Давность заболевания у 7 больных была до 1 года, у 18 — от 1 года до 5 лет, у 4 — свыше 5 лет. Из 29 обследованных больных 9 чел. госпитализировались впервые и ранее не лечились, другие 20 — в прошлом получали курсы инсулиношоковой терапии, а также лечились психотропными средствами, не давшими заметного лечебного эффекта. К моменту настоящего обследования больные имели перерыв в лечении от 5 до 10 месяцев.
Контрольная группа состояла из 35 психически здоровых лиц, преимущественно мужчин в возрасте 25—35 лет.
У всех испытуемых и здоровых лиц и больных до начала лечения пищевой рацион существенно не различался.
Первоначальные исследования газообмена (1, 8, 12, 45) позволили в самых общих чертах составить представление о белковом метаболизме при полном алиментарном голодании.
Было, в частности, показано, что удельный вес белка, как источника энергии, при полном лишении пищи невелик и энергия, приходящаяся на долю белка, не превышает, примерно 1/7 части всей суммарно определяемой энергии. Судя по динамике дыхательного коэффициента, максимальное окисление белка наблюдается в первые дни голодания, непосредственно вслед за быстронаступающей утилизацией углеводных запасов. По мере удлинения сроков голодания (исключая его терминальные периоды) участие белка в энергетическом балансе голодающего организма все более снижается.
Эти данные были вполне подтверждены биохимическими исследованиями диссимиляторных процессов белкового обмена, основанных на изучении общего азота мочи, азота мочевины, аммиака и т. д. (5, 9, 16, 18, 21, 31, 35, 42—44, 47).
Так, было показано, что общий азот, элюминируемый с мочой и образующийся, как известно, на 90% за счет распада белка, обнаруживает заметное увеличение в сроки, примерно, с 3 по 5—7 дни полного голодания крупных животных и человека. После этих сроков устанавливается обычно более ограниченное выделение азота, которое с некоторыми колебаниями в течение довольно длительного голодания продолжает держаться на относительно невысоком уровне.
Результаты наших исследований экскреции азотистых соединений с мочой в процессе лечебного голодания (средние значения) представлены в таблице № 1.
Анализируя полученные нами данные о количестве суточной мочи (табл. № 1), можно видеть, что среднее суточное ее количество оставалось фактически стабильным в течение двух первых периодов голодания. Лишь во второй половине третьего периода голодания (20—25 день) среднесуточное количество выделяемой мочи статистически достоверно увеличивалось по сравнению с исходным фоном на 59,43% (0,01<Р<0,02).
Таблица 1
Динамика азотистых соединении мочи
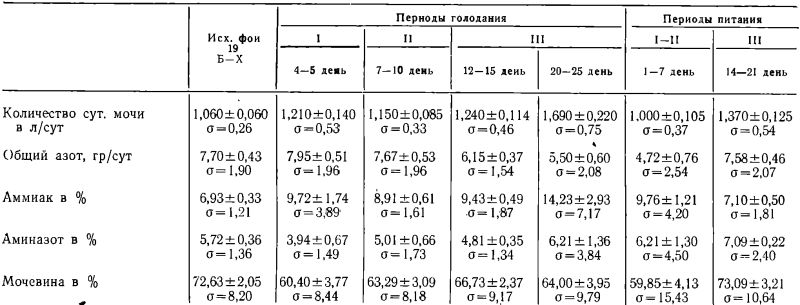
Из той же табл. № 1 можно видеть также, что среднее количество общего азота в суточной моче не обнаруживало существенных колебаний в первые два периода голодания по сравнению с исходным фоном (Р>0,05) и лишь в третьем периоде голодания отмечалась достоверное снижение общего азота по сравнению с его исходным уровнем на 20,10% (0,01 <Р<0,01) — в начале третьего периода (12—15 день голодания), а на 25—90% (0,001<Р<0,001) — в конце третьего периода (20—25 день голодания).
Поле прекращения голодания (в первые два периода питания) продолжалось дальнейшее уменьшение количества общего азота в моче. Так, по сравнению с исходным фоном азот мочи в первые два периода понизился ниже своего исходного уровня до голодания на 38,70% (Р<0,001). Однако, в третьем периоде питания количество выделяемого с мочой азота возросло, по сравнению с первыми двумя периодами питания, на 60,85% (0,001<Р<0,01) и фактически достигло исходного уровня.
Процентное содержание азота аммиака во время голодания неуклонно возрастало: во втором периоде голодания оно достигло (0,01<Р<0,02), а в третьем периоде голодания к 12—15 суткам — 9,43% (Р<0,001) и к 20—25 суткам — 14,23% (0,02<Р<0,05). Во время питания процентное содержание азота аммиака снижалась и в третьем периоде питания (14— 21 сут.) составило 7,10%, т. е. достигло уровня исходного фона.
Процент аминного азота во всех трех периодах голодания и в первых двух периодах питания статистически достоверно не менялся. Лишь в третьем периоде питания (14—21 день) процентное содержание аминного азота возросло до 7,09% (0,001<Р<0,02).
Процентное содержание азота мочевины уже в первом периоде голодания снизилось с 72,63% до 60,40% (0,01<Р<0,02) и оставалось примерно на этом уровне вплоть до третьего периода питания, где к 14—21 дням вновь возросло до 73,09% (0,02<Р<0,05).
Полученные нами данные оказываются во многом совпадающими с данными литературы (21, 39), где в конечные сроки голодания (исключая его терминальные стадии) отмечалось уменьшение выделения азота с мочой, а в начальные сроки голодания, в некоторых случаях, незначительное увеличение экскреции азота с мочой.
Эти данные в свое время трактовались (29,34), с точки зрения гипотезы Е. Фонта (48), полагавшего, как известно, что на энергетические нужды организма в первую очередь расходуется так называемый «лабильный» или циркулирующий белок, в то время как потребление «организованного клеточного белка» предотвращается более интенсивным расходованием вначале углеводных, а затем и жировых запасов.
Следует отметить, что эта точка зрения уже давно была подвергнута критике (10) и не соответствует современным представлениям, признающим универсальные генетико-молекулярные механизмы белкового анаболизма и не проводящей различия между «лабильным» и «организованным» белком. При сдвигах в белковом балансе речь идет, скорее всего об изменении интенсивности анаболических и катаболических процессов в различных органах и тканях, что, в частности, отражается и на «белковой картине крови».
В связи с этим определенный интерес могут представить данные относительно содержания белка и его отдельных фракций в сыворотке крови при полном голодании и последующем питании.
Необходимо отметить, что литературные данные, посвященные этому вопросу, относительно невелики и результаты их во многом неоднотипны.
Так, согласно исследованиям Кордиера Д. с соавт. (31) содержание белка в крови морского угря в течение 15 дней полного голодания не изменялось.
24-часовое голодание (41) не изменило общей картины в распределении компонентов белка в сыворотке крови крыс. Бенедетти А (20) не нашел изменения общего содержания белка в сыворотке крови у 10 тучных больных, голодавших с лечебной целью в течение 5—9 суток. Не изменялось содержание белка в сыворотке крови больных с ожирением в процессе их лечебного периодического голодания (голодание по 10 дней, 3 раза с незначительными перерывами) (44).
В то же время, по данным К. М. Хритиннной (17), исследовавшей 12 полностью голодавших кроликов (голодание свыше 20 дней, без ограничения воды) в большинстве случаев общее содержание белка в сыворотке снижалось в период максимальной потере веса (после 20 дня голодания). В отдельных случаях ее наблюдении общий белок к начале голодания повышался, и лишь после 10—15 дня снижался до нормального уровня.
По Кордиеру Д. (31) при 15-дневном голодании рыб весом 50—100 г общее содержание белка снизилось с 1,67 г% до 0,6 г%.
Увеличение белка в сыворотке крови во время добровольного 33-дневного голодания людей отмечал Шенк Е. Г. и Майер X. Е. (46).
По данным Н. Ф. Солодюк (14) при полном голодании 8 собак содержание общего белка крови увеличилось в период до 5 дня голодания и затем несколько снизилось к 12 его дню.
А. Рапопорт, Г. С. Фром, X. Хасден (42) в первую неделю голодания нашли повышение сывороточного белка в крови у 3 из 4 обследованных женщин, страдавших ожирением и голодавших с лечебной целью 2—4 недели. Во вторую неделю голодания содержание белка несколько снижалось.
Столь же неопределенные данные были получены и при исследовании белковых фракций. Так, согласно Е. Г. Шенку и X. Е. Майеру (46) в начале голодания происходит увеличение и альбуминов, и глобулинов, причем альбумины увеличиваются относительно больше, что приводит к увеличению коэффициента А/Г. Лишь к 30 дню голодания количество глобулинов начало относительно возрастать, что и привело к снижению коэффициента А/Г до первоначального значения.
А. Киммо (30) при абсолютном голодании крыс не отметил изменений А/Г коэффициента. При голодании без ограничения воды, по данным этого автора, концентрация глобулинов падала на 10%.
Кретчмер (32) при 72-часовом голодании мышей обнаружил статистически достоверное уменьшение ?1 и ?-глобулиновых фракций. Фракция у1-глобулинов, статистически достоверно увеличивалась.
Увеличение альбуминовых фракций в случае 5-дневного голодания собак отметила Н. Ф. Солодюк (14).
Е. Генри и Р. Бел (27) при голодании крыс-самцов отметили понижение содержание альбуминов и глобулиновых фракций ?1, ?2 и ?. Концентрация у-глобулинов, напротив, возрастала. Коэффициент А/Г—снижался.
А. Бенедетти (20) при 5—9-дневном голодании 10 тучных больных лишь в отдельных случаях обнаруживал незначительное снижение концентрации альбуминов при увеличении содержания а2-глобулинов.
По данным К. М. Хритининой (17) содержание отдельных фракций белка при голодании изменялось неравномерно. Так, содержание альбуминов уменьшилось абсолютно и относительно, в то время как относительное содержание у-глобулинов возрастало.
А. Рапопорт с соавт. (42) отметили, что у 2 из 4 обследованных ими тучных женщин в первую неделю лечебного голодания повышалось содержание альбуминов, которое затем несколько снижалось при продолжении голодания. У 2 других голодавших альбумины оставались без изменений. Содержание глобулинов у всех 4 больных в процессе 2—З-иедельного голодания оставалось на уровне, близком к исходному.
Ещё менее разработанным оказывается вопрос относительно содержания белка и его фракций в сыворотке крови и последующие периоды питания.
Е. Г. Шенк и X. Е. Майер (46) обнаружили после голодания падение количества как альбуминов, так и глобулинов, причем альбумины снижались относительно менее выраженно, чем, глобулины, что и привело к увеличению А/Г коэффициента. Интересно, что после приема белковосодержащей пищи уровень альбумина в сыворотке крови резко упал (более чем на 50%), а количество глобулинов столь отчетливо увеличилось (А/Г коэффициент снизился). Соотношение альбумины—глобулины нормализовалось лишь к 10 дню восстановительного периода.
По данным Н. Ф. Солодюк (14), уменьшение к концу голодания собак содержания белка в сыворотке крови продолжалось и в первые дни питания оставалось на этом же низком уровне и снизилось еще более отчетливо в срок с 7 по 14 дни питания. Восстановление белкового состава оказалось растянутым у различных испытуемых групп в сроки с 18 по 36 и с 28 по 86 дни восстановительного периода. Вновь отмечаемое в период питания увеличение содержания белка (15) происходит за счет альбуминовой фракции.
Из приведенного краткого обзора можно видеть, что вопрос относительно изменения содержания как общего белка в сыворотке крови, так и его фракций при полном голодании (и тем более при последующем питании) разработан крайне недостаточно. Можно думать, что на неоднозначность приводимых в литературе данных влияло множество факторов. Из них в первую очередь хотелось бы обратить внимание на различия, зависящие, возможно, от того, что наблюдения, приведенные на людях, крайне немногочисленны, в то время как более широкие экспериментальные исследования ставились на животных, различающихся в видовом отношении.
Так, например, Е. Генри и Р. Бел (27) подчеркивали, что белковые фракции у крыс «... более чувствительны к голоданию, чем у собак или человека».
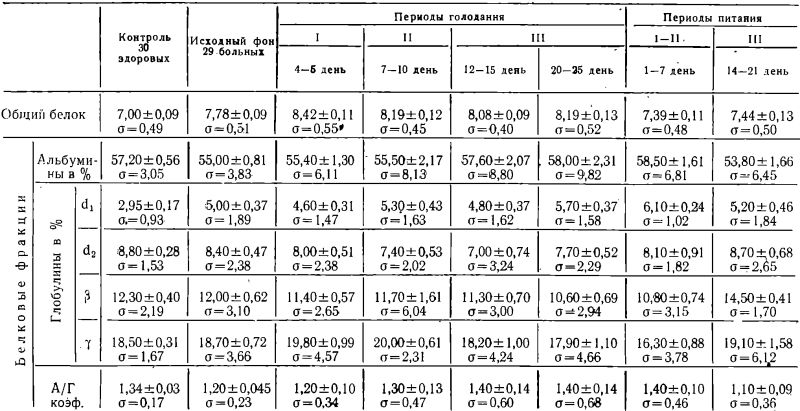
Полученные нами данные содержания общего белка и белковых фракций в сыворотке крови в процессе лечебного голодания после соответствующей математической обработки сведены в таблице № 2.
Как можно видеть из таблицы 2, среднее содержание общего белка в сыворотке крови у больных до голодания превышает уровень белка у здоровых лиц, исследованных в качестве контроля, на 11,14% (Р<0,001).
В первом периоде голодания отмечалось статистически достоверное увеличение белка в сыворотке крови у больных, по сравнению с их исходным фоном на 8,22% (0,001<Р<0,01).
На протяжении всех дальнейших периодов голодания среднее содержание общего белка не уменьшалось ниже данных, полученных для первого периода, оставаясь в то же время несколько выше исходных цифр (соответственно на 5,27%, 0,001 <Р<0,01 — второй период на 3,85%, 0,01<Р<0,02 — 12—15 дни третьего периода и на 6,27%, 0,001<Р<0,01 — 20—25 дни третьего периода).
Следует отметить, что содержание общего белка в начальные сроки третьего периода голодания (12—15 дни) снизилось достоверно по сравнению с уровнем первого периода на 4,03% (0,01 <Р<0,02).
После прекращения голодания содержание сывороточного белка статистически достоверно уменьшилось как по сравнению со средними цифрами всего третьего периода голодания (соответственно на: 9,10% Р<0,001 — в первом—втором восстановительных периодах и на 8,48%, Р<0,001—в третьем периоде питания), так и по отношению к исходному уровню до голодания (соответственно на: 5,01%, 0,02<Р<0,05 — в первом, втором периодах и на 4,37%, 0,02<Р<0,05 — в третьем периоде восстановления).
Из той же таблицы № 2 следует, что в процессе всего курса лечебного голодания в белковых фракциях сыворотки (по средним данным) существенных изменений-не прослеживается, за исключением превышения процента глобулиновой фракции ?1 — у больных до голодания по сравнению со здоровыми лицами на 2,05% (Р<0,001), а также увеличения процента ?-глобулиновой фракций у больных в третьем периоде питания на 2,50% (Р<0,001) по сравнению с исходным фоном.
Таблица 2
Динамика общего белка и белковых фракций (средн. данные)
Немногочисленные работы, посвященные изучению содержания свободных аминокислот в крови при голодании, не представляются однотипными.
Так, С. Мак Дональд (36) не отметил изменений в содержании свободных аминокислот в плазме крови при голодании крыс. Снижение уровня-аминокислот наступало под влиянием введения голодающим крысам адреналина и глюкэгона.
В то же время Л. В. Чорки, А. К. Кого, Д. Ф. Хугам (26) у 6 людей, после их 48-часового голодания отметили увеличение содержания в крови лейцина и валина. Одновременно снижалось содержание в крови треонина, лизина, триптофана, метионина и аргинина. Выведение последних аминокислот с мочой было менее интенсивным, чем до голодания. Выведение с мочой лейцина и валина практически не изменялось.
Аналогичные данные были отмечены Э. Я. Скуинь (13), наблюдавшей, увеличение концентрации в крови гистидина, триптофана, цистина при лечебном голодании больных шизофренией.
Увеличение в плазме крови голодавших кроликов лейцина и валина обнаружили также Е. Марти и П. Рейно (38). Концентрация в крови аланина, глутаминовой и аспарагиновой кислот у этих же кроликов оставалось, без изменения. По мнению авторов, отмеченные изменения состава аминокислот в крови не были следствием биохимических сдвигов в самой крови, поскольку при отключении перфузионной системы (с помощью которой ставили этот эксперимент) от слепой кишки эти сдвиги не отмечались. Поскольку хроматографически было установлено, что в гомогенатах тканей слепой кишки у голодающих кроликов уменьшается содержание свободных аминокислот, авторы заключают, что в обнаруженном ими эффекте известную роль может играть всасывание продуктов автолиза содержимого слепой кишки.
То, что аминокислотное равновесие крови при голодании, скорее всего, зависит от уравновешивания деградирующих аминокислот за счет аминокислот, освобождающихся из тканевых белков, было показано Ф. Кричен и А. Сеффати (24), изучавших длительно голодавших карпов.
Интересный эффект перераспределения аминокислот при голодании описал Н. Н. Демин (2). Согласно его данным, в первые периоды голодания концентрация свободных аминокислот в плазме крови остается почти неизменной, в то время как в эритроцитах концентрация аминокислот возрастает.
С. Ц. Мадден и Ю. X. Уипл (37) обсуждали, в свое время, возможность перехода введенных извне белков в белки тканей голодающего организма- без предварительного глубокого расщепления до аминокислот. Однако эти предположения представляются маловероятными. По крайней мере, согласно данным И. А. Поберий (11), применявшей изотопные и авторадиографические исследования, белки плазмы полностью голодающих крыс не используются для синтеза протоплазмы эритробластов без предварительного расщепления до аминокислот.
Таким образом, можно видеть; что исследования аминокислотного состава при голодании представлены отдельными, довольно разрозненными, работами. Единственный вывод, который можно с большей или меньшей уверенностью сделать в настоящее время на основании вышеприведенных работ, это то, что для голодающего организма характерен феномен перераспределения аминокислотного состава (между различными тканями и кровью, между плазмой и эритроцитами), что следует, на наш взгляд, рассматривать, как одно из проявлений адаптационной, саморегуляторной реакции организма на острое лишение пищи.
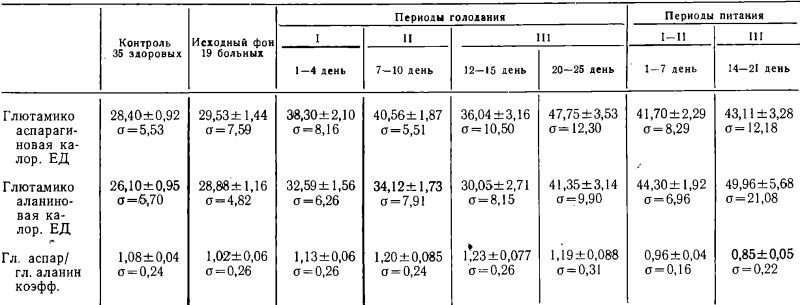
Полученные нами средние данные динамического изучения активности трансаминаз отражены в таблице № 3.
Из таблицы № 3 следует, что активность глютамикоаспарагиновой трансаминазы во время голодания, постепенно увеличиваясь, на 29,72% (Р<0,001)—в первом периоде, на 39,06% (Р<0,001)—во втором, достигла превышения исходного своего значения к конечным срокам третьего периода (20—25 день голодания) на 61,76% (Р<0,001).
После прекращения голодания активность глютамикоаспарагиновой трансаминазы значительно превышала свое исходное значение: на 37,86% (Р<0,001) — в первых двух периодах питания и на 45,01 % (Р<0,001)—в третьем периоде восстановления.
Активность глютамикоаланиновой трансаминазы в сыворотке крови, начиная со второго периода голодания, также увеличивалась по сравнению с исходным уровнем на 18,13% (0,01<Р<0,02) и достигал а увеличения на 43,84 % (Р<0,001) — к конечным срокам третьего периода (20—25 день голодания).
Во время питания активность глютамикоаланиновой трансаминазы превышала исходную на 53,35% (Р<0,001)—в первых двух восстановительных периодах и на 72,94% (Р<0,001)—в третьем периоде питания.
Однако, анализ отношения активности глютамикоаспарагиновой трансаминазы к активности глютамикоаланиновой — гл. асп./гл. алан, коэфф. показывает, что к начальным срокам третьего периода голодания (12—15 день) этот коэффициент увеличивался на 0,21, т. е. на 21,00% (0,02<Р<0,05).
На питании, ко времени первых двух периодов этот коэффициент трансаминаз понижался, по сравнению с его значением с 12—15 сутками третьего периода голодания на 22,50% (0,001<Р<0,01) и, примерно, достигал вновь своего исходного значения. В третьем периоде питания уменьшение гл. асп./ гл. алан, коэффициента трансаминаз продолжалось и оказывалось ниже своего исходного значения на 17,00% (0,02<Р<0,05).
Подобная динамика гл. асп./гл. алан, коэффициента трансаминаз, возможно, говорит от относительном перераспределении в процессе лечебного голодания активности указанных трансаминаз с превалированием активности глютамикоаспарагиновой трансаминазы во время полного голодания и превалирования активности глютамикоаланиновой трансаминазы во время питания.
Таблица 3
Динамика трансаминаз (средние данные)
Следует отметить, что подобную динамику активности трансаминаз в свое время наблюдала Л. И. Ландо (6) у психически больных в процессе их активной терапии.
Подобным же образом активность трансаминаз возрастает и -при влиянии многих неспецифических стрессорных факторов: шум, вибрация, нагревание, введение адреналина (23, 28, 40).
* * *
Все вышеизложенное можно кратко резюмировать следующим образом: полное дозированнное лечебное голодание в условиях и в сроки наших наблюдений (20—25 суток голодания с последующим восстановлением), видимо, не приносит патологии в процесс белковообразовательной функции печени.
Сдвиги в изучавшихся нами компонентах белково-азотистого обмена в процессе лечебного голодания носят неспецифический, стрессорный характер и могут рассматриваться как частный пример адаптационных, саморегуляторных изменений, лежащих в основе поддержания гомеостаза при полном голодании.
ЛИТЕРАТУРА
1. Авроров П. П. Обмен веществ и развитие энергии в организме при полном голодании. Дисс. докт., М., 1900.
2. Демин Н. Н. Журн. Вопр. питания, 1941, 10, 3—4, 16.
3. Дюбуа Е. Обмен веществ, И. Л., М., 1927.
4. Ежова Е. Н. и др., в кн.: Труды 3 Всесоюзн. съезда физиологов, Л., 1928, т..№ 1, 227.
5. Ежова Е. Н., Молчанова О. П. и др., в кн.: Сб работ по физиологии I Моск. мед. ин-та, М.^1939, 85.
6. Ландо Л. И. в кн.: Труды 4 Всесоюзн. съезда невропат, и психиатров, т. 3, в. 1, М., 1965, 486.
7. Николаев Ю. С. Разгрузочно-диетическая терапия шизофрении и ее физиологическое обоснование. Дисс. докт., М., 1959.
8. Охотин И. А. Патологоанатомические изменения и газовый обмен у голодающих кроликов. Дисс., Спб, 1885.
9. Пашутин В. Курс общей и экспериментальной патологии, т. 2, ч. 1, Спб., 1902.
10. Перельман Л. Р. Руководство по патологической физиологии. Под редакцией А. А. Богомольца, Киев, 1947, т. 2, гл. 12, $9.
11. Поберий И. А. Цитология, 1964, т. 6, № 3, 330.
12. Садовень А. А. в кн.: Труды русского общества охраны народного здоровья, 1888, в. 12, 88.
13. Скуинь Э. Я. в кн.: Вопросы психиатр., Авторефераты научных работ ин-та психиатр. МЗ РСФСР (1945—1953), М., 1956, 274.
14. Солодюк Н. Ф. Физиол. журнал, 1958, 4, 450.
15. Солодюк Н. Ф. Восстановление белкового ~и морфологического состава крови после кровопотери и голодания (экспериментальное исследование). Дисс. докт., Киев, 1958.
15а. Солодюк Н. Ф. Дисс. докт. мед. наук, Киев, 1958.
16. Фром гольд Е. Е., Михайлов И. Ф. Терапевт, архов, 1925, т. 3, в. 5—6, 109.
17. Хритинина К. М. в сб.: Некоторые вопросы физиологии и биофизики. Воронежский ин-т, 1964, 245.
16. Шатерников М. Н. Русская клиника, 1929, т. 12, № 68, 860.
19. Albschul е W. D., Altschule L. Н., Tillotson К. J-J. clin. Endocrinol., 1949, g. 548.
20. Benedetti A., Arch studio fisiopatol. e. clin. rikambio, 1959, 23, 1, 24.
21. Benedict F. A study of prolonged fasting, 1915.
22. Brodie В. В., Shore P. A., Ann. N. J. Acad. Sci., 1957, v. 66, p. 631.
23. Cope F., Pol is B. Javiat. Med., 1959, v. 30, p. 90.
24. Сreасh V., S e г f a t у A. Compt. rend. Soc. beol., 1964, 158, 5, 1152.
25. Charkey L. W., Kano Adeline K., Hangman Dunne F.f Nutrition, 1955, 55, 3, 469.
26. Heath R. G., Leach В. E. Arch. Neurol. Psychiat., 1956, 76, 444.
27. Henrу E., Вell, Rоbert T.t N i s h i h а г a H i s a k o. Arc. Soc., Expte Biol..and Med., 1959, 100, 4, 853.
28. Highman В., M a 1 i n g H., Thompson E. Am. J. Physiol., 1959, v. 196, p. 436.
29. К a p 1 а л S. A., Shimizu C. G. N. Amer. J. Physiol., 19G2r 202, 4, 695.
30. К i ni m о A h o, As to la Erva. Ann. med. exptl. et biol. fen-rial., 1956, 34, 4, 341.
31. К о г d i e г D., Barnoud R., Brandon A. M. Compt, rent. Soc. biol., 1957, 1958, 151, 11, 1912.
32. Kretzschmar, Z. ges. innere Med., 1957, 12, 10, 472.
33. L a b о г i t H., С о i г a u 1 t R., В г о u s s о 11 e В., et al.r Ann. med.-psychol., 1958, v. 116, 2, p. 60.
34. Leuman, Mueller, Munk, Senatar, Zuntz, Virch. Aich. f. pathol. Anat. und Physiol., 1893, 131, Suppl. I, Heft I.
35. L u с i a n i L. Der Hunger, 1890.
36. Mac Donald Sara J., De Boer Katharine O. Proc, Soc. Expte. Biol. and Med., 1965, 119, 4, 1221.
37. Madden S. С., Whipple J. H. Physiol. Rev., 1940, 20, 1, 194.
38. Marty J., Raunaud P., Arch Sci. physiol., 1965, 19, 3, 213.
39. M о г g u 1 i s S., Fasting and undernutrition, N. J., 1923.
40. Nosek J., S eve la M. Csl. Gastroen, Vyz., 1959, t. 13, 559.
41. Peterson Ruth D., Beatti Clarissa n, Amer. J.^ Physiol., 1958, 193, h 79.
42. R a p p о p о г t A., From G. Z., H u s d a n H. Metabolism, 1965, 14, 1, 47.
43. R e i s s Eric, Metabolism, 1959, 8, 2, 151.
44. Riet H. G., Van Schwarz F.f Kinderer V. J. Metabolism, 1964, 13, 4, 291.
45. R u b n e r M., Die Gesetze des Energieverbrauchs bie Ersnahrung, Leipzig—Wien, 1902.
46. S с h e n с k E. G. und Meyer H. E. Das Fasten, Stuttgart— Leipzig, 1938.
47. Swendseid M. E., Mulcare D. В., Drenick E. J. J. Amer. Diet. Assoc., 1965, 46, 4, 276.
48. Voit E„ Zeitschr. f. Biologie, 1866, 2, 3, 309.
49. Wooley D. W., Shaw E. N. Ann. N. J Acad. Sci., 1957, v. 66, p. 649.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Развитие идей лечебного голодания Ю. С. НИКОЛАЕВ (Москва)
Развитие идей лечебного голодания Ю. С. НИКОЛАЕВ (Москва) Можно считать, что люди еще с доисторических времен применяли голодание с лечебной целью; это подтверждается наблюдениями за животными, которые при заболевании отказываются от приема пищи, этому же инстинктивно
Результаты разгрузочно-диетической терапии психически больных с синдромом дисморфофобии Ю. С. НИКОЛАЕВ, Г. И. БАБЕНКОВ (Москва)
Результаты разгрузочно-диетической терапии психически больных с синдромом дисморфофобии Ю. С. НИКОЛАЕВ, Г. И. БАБЕНКОВ (Москва) Синдром дисморфофобии с давних пор привлекает внимание психиатров, как отечественных (3—15), так и зарубежных: (1,2, 16—18).Мнение о прогностической
Клинико-электроэнцефалографичские корреляции у больных шизофренией с вялым течением в процессе лечебного голодания и последующего питания Б. В. КРАЙЦЕРОВ (Москва)
Клинико-электроэнцефалографичские корреляции у больных шизофренией с вялым течением в процессе лечебного голодания и последующего питания Б. В. КРАЙЦЕРОВ (Москва) Резистентность некоторых групп больных шизофренией с вялым течением ко многим терапевтическим
О динамике основного обмена жак отражении общей реактивности в процессе лечебного голодания психически больных В. Б. ГУРВИЧ, Г. И. БАБЕНКОВ (Москва)
О динамике основного обмена жак отражении общей реактивности в процессе лечебного голодания психически больных В. Б. ГУРВИЧ, Г. И. БАБЕНКОВ (Москва) В настоящее время в литературе встречаются указания «а расстройства обмена веществ при различных психических заболеваниях
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва)
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва) Лечение ожирения остается одной из наиболее актуальных проблем современной медицины. Число больных с избыточным весом, согласно данным многих
Случай применения лечебного голодания у тучного больного, перенесшего инфаркт миокарда М.И. МИНЯЙЛЕНКО (Москва)
Случай применения лечебного голодания у тучного больного, перенесшего инфаркт миокарда М.И. МИНЯЙЛЕНКО (Москва) В литературе нам встретились ряд работ, указывающих на эффективность кратковременного воздержания от приема пищи при целом ряде соматических заболеваний.Так
Разгрузочно-диетическая терапия больных гипертонией Я. Я. РУДАКОВ, М. И. МИНЯЙЛЕНКО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва)
Разгрузочно-диетическая терапия больных гипертонией Я. Я. РУДАКОВ, М. И. МИНЯЙЛЕНКО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва) При проведении разгрузочно-диетической терапии больным с психическими расстройствами сосудистого генеза нами наблюдалось достаточное количество
Лечение дозированным голоданием больных инфекционным неспецифическим полиартритом с психическими нарушениями Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ, В. А. БРЮЗГИН, Я. Я. РУДАКОВ (Москва)
Лечение дозированным голоданием больных инфекционным неспецифическим полиартритом с психическими нарушениями Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ, В. А. БРЮЗГИН, Я. Я. РУДАКОВ (Москва) В числе многих больных с так (Называемыми пограничными заболеваниями в нашу клинику поступали
Особенности подвижности основных нервных процессов у больных с различными психическими заболеваниями в процессе лечения их дозированным голоданием Ю. С. НИКОЛАЕВ, В. А. БРЮЗГИН, В. Б, ГУРВИЧ (Москва)
Особенности подвижности основных нервных процессов у больных с различными психическими заболеваниями в процессе лечения их дозированным голоданием Ю. С. НИКОЛАЕВ, В. А. БРЮЗГИН, В. Б, ГУРВИЧ (Москва) В ряде предыдущих сообщений было высказано мнение, что при лечении
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва)
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва) Изучению фагоцитарной активности лейкоцитов при длительном полном алиментарном голодании посвящены
Динамика экскреции адреналина, норадреналина и их предшественников с мочой у психически больных в процессе разгрузочно-диетической терапии Л. Я. ЛАНДО, Г. И. БАБЕНКОВ (Москва)
Динамика экскреции адреналина, норадреналина и их предшественников с мочой у психически больных в процессе разгрузочно-диетической терапии Л. Я. ЛАНДО, Г. И. БАБЕНКОВ (Москва) Симпато-адреналовой системе и ее гормонам и медиаторам — катехоламинам отводится значительная
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва)
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва) Половой хроматин — внутриядерное тельце, дающее Фельген-положительную реакцию и
О динамике основного обмена при лечебном голодании больных с ипохондрическим синдромом В. Б. ГУРВИЧ (Москва)
О динамике основного обмена при лечебном голодании больных с ипохондрическим синдромом В. Б. ГУРВИЧ (Москва) Вопрос об обмене веществ, в частности, основном обмене, является одним из ведущих при объяснении патофизиологических механизмов голодания (1, 5, 9, 11, 12, 14).В. В.
Условнорефлекторные лейкоцитарные реакции в динамике лечебного голодания при некоторых нервно-психических заболеваниях С. М. ЛУРЬЕ (Москва)
Условнорефлекторные лейкоцитарные реакции в динамике лечебного голодания при некоторых нервно-психических заболеваниях С. М. ЛУРЬЕ (Москва) Учение И. П. Павлова «Об организме, как о целостной системе, как о функциональном единстве всех органов и тканей, психического и
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва)
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва) В течение последних лет в нашей стране и за рубежом появилось большое число сторонников применения голодания с лечебной целью при