Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва)
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания
С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва)
Половой хроматин — внутриядерное тельце, дающее Фельген-положительную реакцию и образующееся, согласно современным представлениям, гетеропикнотической Х-хромосомой, в связи с чем оно встречается лишь у индивидуумов с 2 и более Х-хромосомами. Гетеропикнотизация Х-хромосомы приводит к инактивации (по мнению некоторых авторов, частичной) генов (10).
Вопрос об изменчивости полового хроматина привлекает к себе все большее внимание исследователей. В психиатрии, в частности, актуальность этой проблемы обусловлена широким внедрением метода определения «ядерного пола» в повседневную лабораторную практику, что несомненно требует тщательного изучения «нормальных» вариаций полового хроматина.
Как известно, в психиатрии терапевтическая тактика связана, как правило, с рядом мощных стрессорных воздействий, которые, согласно отдельным данным, могут изменять количество и некоторые качественные характеристики полового хроматина (14).
С другой стороны, условия содержания психически больных, отличающиеся в зависимости от характера заболевания и структуры лечебных учреждений, также, по-видимому, могут отразиться на морфологических особенностях и функции клеток, в частности, клеток эпителия ротовой полости, служащего обычно материалом для определения полового хроматина (19).
Теоретическое значение изменчивости полового хроматина связано с проблемой механизма регуляции его количественных и морфотинкториальных параметров, а также с вопросами поддержания генного баланса, «компенсации дозы гена» и т. д. (10).
Несмотря на значительный интерес указанной проблемы, данные относительно изменчивости полового хроматина недостаточно однородны и убедительны, что связано, в частности, с тем, что единичные работы, выполненные в этом направлении, различны по применявшимся методам и используемым моделям.
Показано, что количество полового хроматина может зависеть от метаболизма и т. д. Рядом исследователей показано изменение в частоте хроматни-положительных ядер и в морфологии (размеры, степень окрашиваемости) полового хроматина при лечении антибиотиками (18), преднизолоном (1), кортикостероидамн (8), при применении электросудорожной терапии (13). Лечение адаптивными гормонами приводит у больных женщин, как правило, к снижению количества хроматин-положительных ядер.
Попыток проанализировать количественные и качественные закономерности полового хроматина в динамике стрессорной ситуации, насколько нам известно из литературных данных, не было проведено[15]. Кроме того, в указанных выше работах изменение полового хроматина не сопоставлялось с клинико-физиологическими параметрами стрессорной реакции, а применявшиеся стрессы были разнообразны по темпу нарастания отклоняющего фактора.
Мы попытались использовать для изучения изменения полового хроматина полное алиментарное голодание, применяемое в лечебных целях (4). Выбор модели связан, в частности, с тем, что полное длительное алиментарное голодание рассматривается в качестве пролонгированного («дозированного» стресса), что дает возможность проведения некоторых корреляций (9, 10).
Объект и методика исследования
Половой хроматин изучался в клетках буккального эпителия по общепринятой методике с окраской ацеторсеином (15). Подсчитывался процент интерфазных ядер с половым хроматином и средняя площадь поверхности телец Барра. С помощью рисовального аппарата РА-4 на миллиметровую бумагу наносились контуры 30—50 телец полового хроматина. Рассчитывалась площадь, ограниченная каждым контуром, суммарная и средняя площадь для каждого тельца. Площадь выражалась в условных единицах, что на наш взгляд является достаточным для изучения ее динамики. Исследования проводились при увеличении 90x15.
Были обследованы женщины в возрасте от 25 до 60 лет (шизофрения — 1, фобический синдром — 4, инволюционная депрессия — 7), из них с нормальным весом (от 53 кг до 75 кг) — 10 больных, с ожирением (125—134 кг) — 2 больных. Лечение голоданием проводилось в клинике экспериментальной терапии психозов (зав. — проф. Ю. С. Николаев). Минимальный срок голодания 8 дней, максимальный — 30 дней.
Исследование проводилось до начала голодания (исходный фон), в 1—4 дни голодания (первый период), 5—14 дни (второй период), с 15 дня до конца голодания (третий период) и в первые дни начала питания. Всего было обследовано 12 больных, проведено 90 исследований.
Результаты исследования
В связи с тем, что при предварительном анализе были отмечены различия в динамике количества и площади поверхности полового хроматина у больных с высоким и с обычным исходным весом, анализ данных был нами проведен отдельно по указанным группам.
У больных с нормальным исходным весом (10 человек) в первом периоде голодания отмечается некоторое (статистически вероятное) увеличение процента хроматин положительных ядер. По мере дальнейшего увеличения сроков голодания количество полового хроматина неуклонно снижается, приобретая в III периоде высокую достоверность (Р<0,001) (график 1).
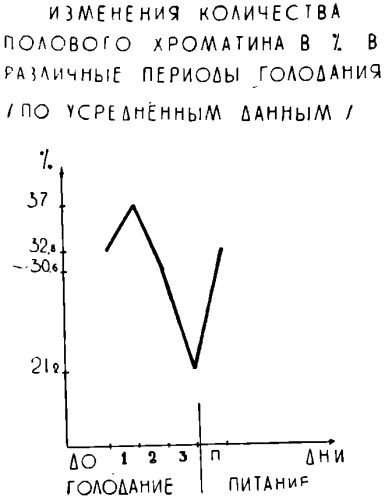
Как видно из этого графика, количество полового хроматина в III периоде голодания составляет в среднем 21,2% сравнению с 32,8% исходного уровня.
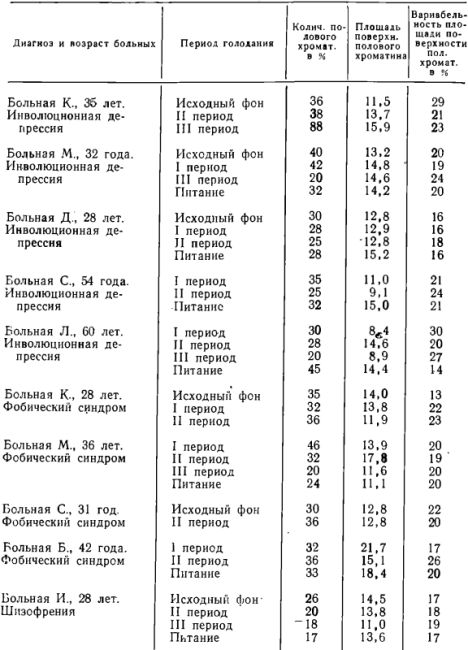
В восстановительном периоде количество полового хроматина вновь увеличивается, приближаясь к исходному уровню (<0,05) (табл. 1 и график 2).
При анализе табл. 1 и граф. 2 видно, что у большинства изученных больных изменение количества полового хроматина проходило довольно однотипно. Исключение составляет больная И-ва (№ 6, граф. 2), у которой в восстановительном периоде количество полового хроматина продолжало уменьшаться, хотя по сравнению с III периодом это уменьшение не было статистически достоверным.
Таблица 1
Количество полового хроматина в % и площадь поверхности полового хроматина у психически больных женщин с нормальным весом в разные периоды полного длительного алиментарного голодания
Анализ данных по средней площади поверхности полового хроматина в различные периоды голодания показывает, что существенных изменений в размерах последней не происходило. Колебания усредненных значений площади поверхности находились в пределах ошибки исследования (фон — 13,1 ±0,4; 1 период — 13,7±1,3; II период — 13,5±0,6; III период — 12,4±1,3).
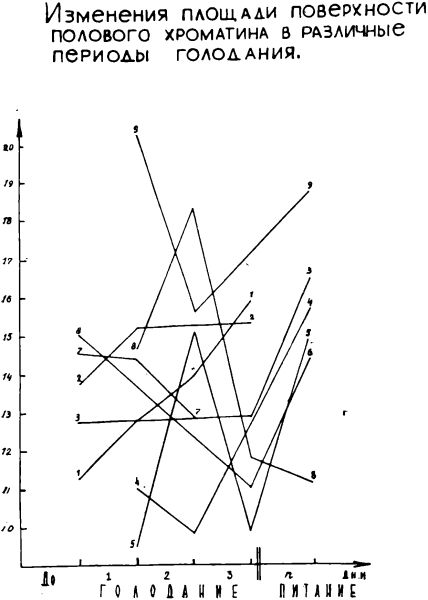
В первые дни питания средняя площадь поверхности полового хроматина значительно возрастала и достоверно (Р<0,01) превышала значение соответствующих цифр исходного периода.
В различные периоды голодания средние значения площади поверхности полового хроматина варьируют у всех больных (граф. 3). В отдельных наблюдениях колебания этого параметра в I, II и III периодах голодания достоверно отличаются от предыдущего.
Следует отметить, что нам. не удалось обнаружить каких-либо корреляций между изменением количества и площади поверхности полового хроматина.
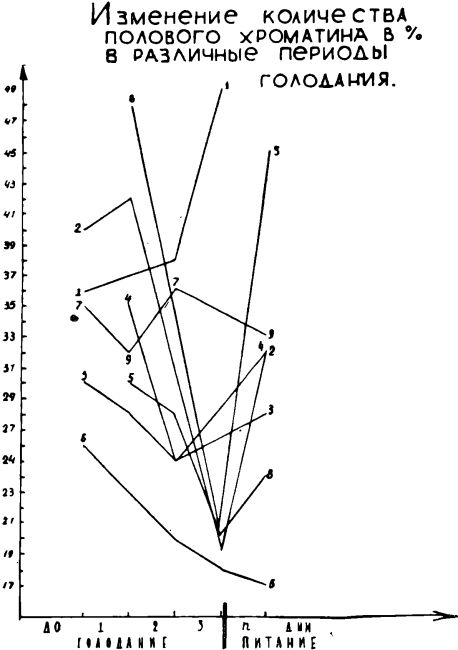
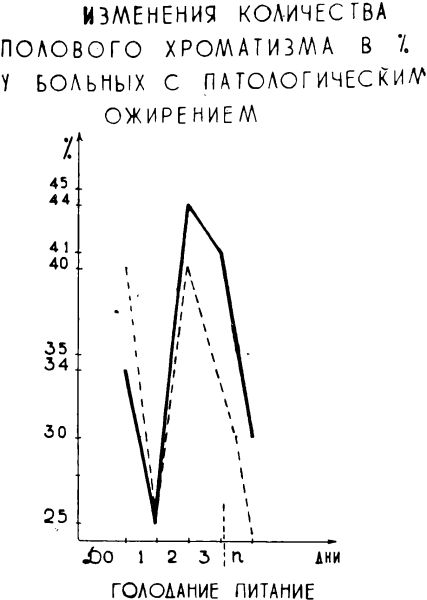
У двух больных с ожирением динамика количества полового хроматина отличалась от средних закономерностей, отмечаемых для больных с нормальным исходным весом (график 4).
Данные по изменению площади поверхности полового хроматина у больных с ожирением представлены в таблице 2.
Таблица 2
Количество полового хроматина в % и площадь поверхности полового хроматина у психически больных женщин с ожирением в разные периоды полного длительного алиментарного голодания

Обсуждение результатов
Анализ полученных данных весьма затруднителен как в связи с относительно небольшим количеством наблюдений, так и с недостаточной разработкой патофизиологических механизмов, лежащих в основе голодания. Исходя из факторов, могущих оказать влияние на количество и морфотинкториальные свойства полового хроматина, в первую очередь возникает мысль о проведении корреляций между вышеуказанными параметрами и некоторыми показателями, отражающими адаптивные перестройки в голодающем организме.
Как уже говорилось, голодание выступает в качестве пролонгированного стресса и сопровождается, в частности, изменением функции коры надпочечников (10). Так, согласно данным отдельных авторов, активность коры надпочечников существенно возрастает в I и особенно в III периодах полного длительного алиментарного голодания, нормализуясь во II периоде (7 и др.).
Как видно из приведенных нами данных, наиболее выраженное количество падения полового хроматина приходится именно на III период голодания, т. е. совпадает во времени с максимальной активизацией коры надпочечников.
Естественно, указанный параллелизм не является достаточным доказательством существования связи между отмеченными сдвигами. Тем не менее, он не противоречит вышеизложенным данным о снижении количества полового хроматина при лечении кортизоном и АКТГ (8).
Обращают на себя внимание существенные различия в изменении полового хроматина в I периоде голодания у больных с нормальным и избыточным весом, что, по-видимому, стоит в связи с клинико-физиологическими особенностями протекания голодания у указанных групп. Можно думать, что особенности поведения полового хроматина в I периоде голодания у больных с ожирением связаны с большей биологической значимостью лишения пищи по сравнению с лицами с нормальным весом[16].
Вопрос о механизмах влияния гормонов на количество и площадь поверхности полового хроматина не представляется сколько-нибудь ясным. По мнению некоторых исследователей, гормоны непосредственно влияют на степень спирализации Х-хромосомы, активизируя отдельные локусы или хромосомные сегменты при ряде ситуаций, требующих повышенной выработки отдельных ферментов (2).
По мнению других (3, 20), уменьшение процента хроматин-положительных ядер при повышенной активности стероидных гормонов связано с резким смещением в соотношении длительности отдельных фаз клеточного цикла. Как показано (11, 12, 16), частота клеток с половым хроматином различна в популяциях клеток, отличающихся по митотическому индексу. Согласно отдельным данным (9) в клетках с укороченным циклом половой хроматин может не выявляться.
Анализ материала по изменению площади поверхности полового хроматина в различные периоды голодания представляется затрудненным в связи с некоторой неоднородностью сдвигов у разных больных.
Таким образом, полученные нами данные относительно динамики изменения количества полового хроматина укладываются в представлении о стрессорной природе сдвигов, наблюдаемых в полностью голодающем организме, хотя последние требуют дальнейшего изучения (17 и др.).
ЛИТЕРАТУРА
1. Беленький Г. Б., Егоркина Д. А., Кострова А. А., Кряжева С. С. Клинич. медицина, 1967, 4, 32.
2. Дэвидсон Э. «Гормоны и гены». В кн.: «Молекулы и клетки». М., 1967.
3. Епифанова О. И. «Гормоны и клеточное размножение». М., 1965.
4. Николаев Ю. С. «Лечение нервно-психических заболеваний дозированным голоданием» (Инструктивно-методическое письмо). М., 1965.
5. Селье Г. «Очерки об адаптационном синдроме». М.( 1960.
6. Шапиро Ю. Л. «Состояние системы крови при полном длительном алиментарном голодании и последующем питании людей». Дисс. канд., М., 1964.
7. Шапиро Ю. Л., Минскер Э. И., Орловская Д. Д. Патологическая физиология и экспериментальная терапия, 1963, 3, 70.
8. Fraccaro М. et al. Lancet, 1962, 1, 1379.
9. J a m е s. Z. Zellforsch., 1964, No. 2, 178.
10. Layon M. F. Lancet, 1963, 2, 1120.
11. Mars De. Science, 1964, 146, 424.
12. Miles. Acta cytol., 1962, 6, 84.
13. P i г о z у n s k i T.,S с г i p с а г u G. H. et al. Ann. Med. psy?hol., 1965, 123 (1), 4, 609.
14. Piatt L. I., К ail in E. W. J. Amer. Med. Ass., 1964, 187, 3 182
15. Sanderson A R. Lancet, 1960, 7136, 1252.
16. Schnedl W. Acta anat., 1964, 57, 52.
17. Schultz A. L., Keklow A., Ulstson R. A. J. Endo-ciinul. and metabol., 1964, 24, No. 12, 1253.
18. Sohval А. В., Casselman W. G. B. Lancet, 1961, 2, 7217:1386.
19. S q u i г s e s B. Brit. J. Nutr., 1963, 17, 303.
20. Therkelsen A. a. Lamm L. Exptl. Cell Ress, 44, 363.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Клинико-электроэнцефалографичские корреляции у больных шизофренией с вялым течением в процессе лечебного голодания и последующего питания Б. В. КРАЙЦЕРОВ (Москва)
Клинико-электроэнцефалографичские корреляции у больных шизофренией с вялым течением в процессе лечебного голодания и последующего питания Б. В. КРАЙЦЕРОВ (Москва) Резистентность некоторых групп больных шизофренией с вялым течением ко многим терапевтическим
О динамике основного обмена жак отражении общей реактивности в процессе лечебного голодания психически больных В. Б. ГУРВИЧ, Г. И. БАБЕНКОВ (Москва)
О динамике основного обмена жак отражении общей реактивности в процессе лечебного голодания психически больных В. Б. ГУРВИЧ, Г. И. БАБЕНКОВ (Москва) В настоящее время в литературе встречаются указания «а расстройства обмена веществ при различных психических заболеваниях
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва)
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва) Лечение ожирения остается одной из наиболее актуальных проблем современной медицины. Число больных с избыточным весом, согласно данным многих
Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва)
Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва) Голодание, как частный раздел науки о питании, за последние годы вновь стало привлекать к себе пристальное внимание в связи со все более широким его
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва)
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва) Как известно, при полном алиментарном голодании в течение довольно длительного времени (по крайней мере до сроков, характеризующихся потерями
Сывороточное и эритроцитарное железо при длительном голодании людей Ю. Л. ШАПИРО, Л. М. ДОНДЫШ, Л. М. ЛЕЙБИН, Э. А. ЛЕЙЗЕРОВИЧ, Л. Ф. ЛЕВИНА (Москва)
Сывороточное и эритроцитарное железо при длительном голодании людей Ю. Л. ШАПИРО, Л. М. ДОНДЫШ, Л. М. ЛЕЙБИН, Э. А. ЛЕЙЗЕРОВИЧ, Л. Ф. ЛЕВИНА (Москва) Данные о содержании железа в жидкой фракции крови при полном алиментарном голодании и последующем питании немногочисленны и
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва)
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва) В литературе имеется значительное число работ, посвященных изучению изменения степени насыщения крови кислородом при разнообразных условиях и
Сравнительное изучение действия полного длительного голодания И белковой недостаточности на состав периферической крови мышей СС57 Вr И. Л. ПОВЕРИЙ и В. И. ПРИЛЯЦКИЙ (Москва)
Сравнительное изучение действия полного длительного голодания И белковой недостаточности на состав периферической крови мышей СС57Вr И. Л. ПОВЕРИЙ и В. И. ПРИЛЯЦКИЙ (Москва) Полное или частичное голодание является удобной экспериментальной моделью для изучения влияния
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва)
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва) Изучению фагоцитарной активности лейкоцитов при длительном полном алиментарном голодании посвящены
К вопросу о реакции различных форм ожирения на голодание Д. Д. ФЕДОТОВ, Ю. Л. ШАПИРО, Ф. А. ВАЙНДРУХ (Москва)
К вопросу о реакции различных форм ожирения на голодание Д. Д. ФЕДОТОВ, Ю. Л. ШАПИРО, Ф. А. ВАЙНДРУХ (Москва) Ожирение, как и вообще проблема неправильного питания, становится в последнее время все более актуальной.В то время, как большая часть человечества (по данным ВОЗа —
О действии полного длительного алиментарного голодания на хромосомный аппарат лимфоцитов периферической крови К. Н. ГРИНБЕРГ, Ю, Л. ШАПИРО, Е. А. КИРИЛОВА, Р. С. КУШНИР (Москва)
О действии полного длительного алиментарного голодания на хромосомный аппарат лимфоцитов периферической крови К. Н. ГРИНБЕРГ, Ю, Л. ШАПИРО, Е. А. КИРИЛОВА, Р. С. КУШНИР (Москва) Полное алиментарное голодание успешно применяется при лечении некоторых психических и
Условнорефлекторные лейкоцитарные реакции в динамике лечебного голодания при некоторых нервно-психических заболеваниях С. М. ЛУРЬЕ (Москва)
Условнорефлекторные лейкоцитарные реакции в динамике лечебного голодания при некоторых нервно-психических заболеваниях С. М. ЛУРЬЕ (Москва) Учение И. П. Павлова «Об организме, как о целостной системе, как о функциональном единстве всех органов и тканей, психического и
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва)
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва) Проблема влияния голодания на разные показатели обменных процессов в организме животных и человека продолжает привлекать внимание
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва)
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва) В связи с тем вниманием, которое в настоящее время уделяется роли лизосом в развитии повреждения клеток тканей различных
Некоторые данные относительно белково-азотистого обмена в процессе лечебного голодания психически больных Л. И. ЛАНДО, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. Я. БАБЕНКОВ (Москва)
Некоторые данные относительно белково-азотистого обмена в процессе лечебного голодания психически больных Л. И. ЛАНДО, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. Я. БАБЕНКОВ (Москва) Белково-азотистый обмен при полном алиментарном голодании как в эксперименте на животных, так и у людей